 2004, Vol. 40
2004, Vol. 40文章信息
- 张国盛, 刘海东, 李玉灵, 王林和, 吉川贤.
- Zhang Guosheng, Liu Haidong, Li Yuling, Wang Linhe, Ken Yoshikawa.
- 切断匍匐茎对臭柏叶绿素荧光特性的影响
- Effects of Severing Stolon on Chlorophyll Fluorescence Characteristics of Sabina vulgaris
- 林业科学, 2004, 40(3): 60-65.
- Scientia Silvae Sinicae, 2004, 40(3): 60-65.
-
文章历史
- 收稿日期:2003-04-01
-
作者相关文章


2. 冈山大学农学部 日本冈山 700-0803;
3. 中国科学院植物研究所植被数量生态学重点实验室 北京 100093
2. Faculty of Agriculture, Okayama University Okayama700-0803, Japan;
3. Laboratory of Quantitative Vegetation Ecology, Institute of Botany, CAS Beijing 100093
近些年来,叶绿素荧光测定已广泛应用于植物生理学、植物生理生态学研究的各个领域。杨胜铭等(2001)对光合作用中激发能分配的调节与光破坏防御的关系,陈晓英等(2002)对类囊体的空间结构及其对激发能分配的调节,吴长艾等(2001)对叶黄素循环及其调控特性进行了综述报道。张守仁(1999)对叶绿素荧光动力学参数的意义、陈贻竹等(1995)对叶绿素荧光技术在植物环境胁迫研究中的应用作了专项报道,荧光动力学参数的生态学意义及参数计算方法趋于一致,增强了测定结果的可比性。随着各种荧光测定仪器的出现,人们不仅能迅速地运用叶绿素荧光测定方法研究各种环境胁迫对植物光合作用的影响(李敦海等,2000;林植芳等,1999;2000;何维明,2000;何维明等,2000;肖春旺等,2001;姜闯道等,2002;苏行等,2002;冯建灿等,2002;张教林等,2002),而且,叶绿素荧光动力学技术在测定叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用。因此,叶绿素荧光动力学技术被称为测定叶片光合功能的快速、无损伤探针(Genty et al., 1989;Schreiber et al., 1994;张守仁,1999)。
毛乌素沙地位于中国西部半干旱区(38°57′~39°01′N,109°02′~109°17′E),是一个脆弱生态系统(张新时,1994)。该区年均气温6.5℃,多年平均降水量360 mm,多集中在7—9月,约占年降雨量的60%~70%,年均蒸发量2 300 mm。风大沙多,地带性土壤为栗钙土,其上发育有风沙土,流动沙丘约占65%。所以,在毛乌素沙地,风沙活动与风沙土的水分含量成为该区的重要生态因子和限制因子,确定了该区植物的优势生活型是耐风蚀与干旱的灌木。臭柏(Sabina vulgaris)又名叉子圆柏、沙地柏,是一种重要的克隆植物,在中国主要分布在青海、新疆、甘肃、宁夏、内蒙古、陕西等海拔1 100~2 200 m的地区,也是毛乌素沙地唯一的天然常绿灌木。臭柏的匍匐茎与地面接触后,能萌发形成不定根系,从而实现其扩展和繁殖,并使沙丘固定。因此,臭柏对毛乌素沙地生态环境的稳定与保护起着十分重要的生态作用,引起国内外生态学者的关注,并对其生理生态学特性、天然更新、根系分布、生存环境等进行了广泛研究(张国盛等,1999;2001;王林和等,1998;2002;山中典和等,2001;温国胜等,2000;Hirobe et al., 2001)。近几十年来,由于环境恶化,特别是人类及牛、羊践踏干扰频繁,导致臭柏匍匐茎受损或断裂,群落衰退,面积锐减。然而,关于自然条件下践踏干扰对臭柏生育的影响程度、匍匐茎所产生的不定根对消除这种干扰的作用的报道很少。本文采用了切断匍匐茎模拟践踏干扰的方式,研究了这种干扰对臭柏叶绿素荧光特性的影响。
1 材料与方法 1.1 材料选择及准备试验地位于内蒙古鄂尔多斯市乌审旗图克苏木境内,毛乌素沙地开发整治研究中心北试验区的天然臭柏分布区(38°18′N,109°38′E,海拔1 320 m)。首先在天然臭柏灌丛中,选择一个生长良好的灌丛作为测定样丛,样丛与丘间地(有季节性积水)的相对高差为4 m,灌丛大小为37 m×46 m,灌丛中央高度为110 cm。从样丛外缘向中央的5 m范围内,选择明显可辨的、产生于同级匍匐茎,中部受沙埋,长40~50 cm,并产生不定根的2个相邻分枝作为一组测定枝,一个分枝进行切断处理,另一个分枝不切断作为对照,共选择3组。在每个测定枝上,从梢部向基部区分为:梢部(无不定根区)、中部(有不定根区)和基部(连接上一级匍匐茎的无不定根区)3个测定样段,在各测定样段选择2个长势良好的小枝用作荧光测定,并用数字胶带标记。切断处理于2002年8月15日19:00进行,切口位于与上级匍匐茎分枝处,切断后立即用凡士林涂抹切口。
1.2 荧光参数的测定2002年8月14日,对标记的测定小枝,在每个小枝顶端向下约2 cm处选择直径约2 mm、带有刺状或鳞片状叶的光合区作为荧光参数测定区,并小心地用23 mm宽的荧光胶带标记出测定区。在样丛内安放温湿度自动仪(Thermo Recorder RS-11,Tabai ESPEC Corp),开始记录测定期间的温度和湿度变化。使用德国Walz公司生产的MINI-PAM荧光测定仪,分别于8月15日(切断前)、16日、24日进行测定。其中,8月15日和16日的测定开始于9:00,然后每隔3 h测定一次,直到18:00结束;8月24日的测定,于8:30开始,9:00以后每隔1 h测定一次,直到19:00结束。最大荧光强度(Fm)和最小荧光强度(Fo)在8月19日黎明前约4:00测定。每次测定时,使探头位于紧靠荧光胶带的上部,尽量小心避免小枝受到损伤。同时,在8月24日,用Li-1600(Li-COR生产)测定标记小枝的气孔导度(gs),测定于8:30开始,10:00以后每隔2 h测定一次,直到19:00结束。8月15日、16日、24日3 d的温湿度变化趋势基本相同(图 1)。

|
图 1 8月15日、16日和24日试验地温湿度状况 Fig. 1 Temperature and relative humidity in experimental area on August 15, 16 and 24 |
在黑暗条件下(黎明前约4:00)测定的Fm和Fo分别表示PSⅡ反应中心处于完全开放时的最大荧光产量和最小荧光产量,Fv/Fm是PSⅡ最大的光能转化效率(其中Fv=Fm-Fo);在光照条件下测定实际荧光值(F)和最大荧光值(F′m),用ΔF/F′m表示PSⅡ实际光化学量子产量,它反映PSⅡ反应中心在有部分关闭情况下的实际原初光能捕获效率,其中ΔF=F′m-F。ETR是表观光合电子传递速率,即ETR=ΔF/F′m×PAR×0.5×0.84,其中PAR为光合有效辐射(μmol·m-2s-1)。用非光化学猝灭(NPQ)反映PSⅡ天线色素吸收的光能不能用于光合电子传递而以热形式耗散掉的光能部分,即NPQ=Fm/F′m-1(张守仁,1999)。
对照与处理样枝以及匍匐茎不同部位样枝之间各叶绿素荧光参数的比较均采用One-way ANOVA方差分析和LSD多重比较方法(SPSS10.0 for Windows软件,1999)进行分析。
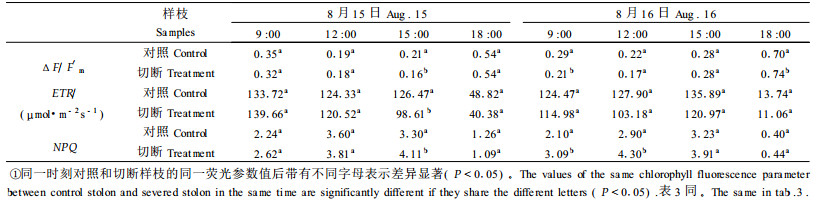
2 结果与分析 2.1 切断匍匐茎对叶绿素荧光参数日进程的影响臭柏匍匐茎上不论是对照还是切断处理小枝的ΔF/F′m、ETR及NPQ 8月15, 16, 24日的日进程(表 2、3)随着PAR(表 1)的变化而变。ΔF/F′m随着PAR增强,逐渐下降,在午间达到最低值,午后开始缓慢上升;而ETR和NPQ的变化与Δ F/F′m相反,表现出与PAR相似的特征(表 2、3)。8月24日的日进程(表 3)表明,对照和切断处理小枝的ΔF/F′m从8:30起随着PAR增高,逐渐下降,在1 2:00—16:00,PAR保持在1 400 μmol·m-2 s-1左右的高光下,形成一个低值区。NPQ日进程与ΔF/F′m相反,从8:30起逐渐上升,最高值出现在15:00,随后下降。对照和切断处理的ETR日进程略有差异,对照ETR在10:00左右达到全天最大值(108.7 μmol·m-2 s-1),17:00形成第二个峰值(82.2 μmol·m-2 s-1)。切断处理样枝ETR在8:30出现第一个峰值(90.1 μmol·m-2 s-1),16:00出现第二个峰值(65.3 μmol·m-2 s-1)。ETR日进程略呈“双峰”曲线的趋势。
|
|

|
|
|
|
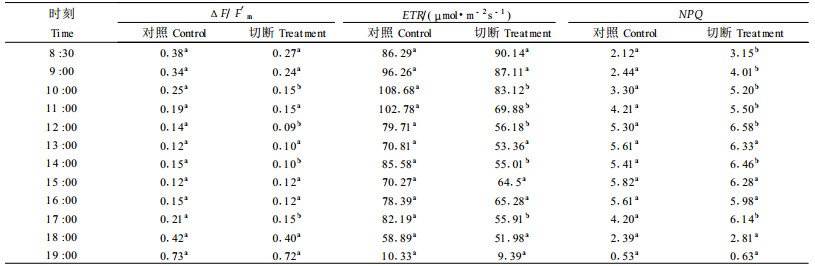
匍匐茎被切断前(8月15日)的测定结果(表 2)表明,对照与准备进行切断处理的小枝的ΔF/F′m、ETR及NPQ值,除在15:00差异显著(P<0.05), 差值皆为20%左右外,其它时段差异不显著(P<0.05),差值平均为8%左右。经切断匍匐茎处理后(表 2、3),被实施切断处理的小枝的ΔF/F′m、ETR及NPQ皆受到影响。8月16日,切断处理的第一天,9:00和18:00切断处理小枝的ΔF/F′m与对照间的差异显著(P<0.05),9:00较对照平均低约25%;ETR与对照间的差异虽不显著,但平均降幅达13 %,午后差值减小;NPQ在9:00和12:00与对照间的差异显著(P<0.05),午前平均增幅高达47%,午后平均增幅减至15%(表 2)。8月24日即切断处理一周,切断处理小枝的ΔF/F′m与对照小枝之间差异显著(P<0.05)率约为33%,ETR的差异显著(P<0.05)率约为43%,NPQ的差异显著(P<0.05)率约为58%。13:00之前,切断匍匐茎处理小枝的ΔF/F′m和ETR比对照平均低29%和19%。1天中同一时刻切断处理小枝与对照相比,ΔF/F′m的最大降幅出现在10:00,降低40%;ETR的最大降幅出现在1 4:00,降低36%。NPQ午前较对照平均高40%。14:00后,除ETR保持与午前相同的降低率(平均为19%)外,ΔF/F′m的平均降低值由午前的29%减小为15%;NPQ平均增高值由午前的40%减小为19%(表 3)。
对照和切断匍匐茎叶绿素荧光特征参数值的One-way ANOVA方差分析和LSD多重比较结果(表 2、3)表明,随着切断处理后时间的延长,切断匍匐茎上的小枝的ΔF/F′m、ETR和NPQ与对照的差异有增大的趋势。但方差分析结果(表 4)表明,切断处理后,匍匐茎各部位的Fv/Fm值与对照间差异不显著(P>0.05)。这表明,虽然臭柏匍匐茎遭到了切断干扰胁迫,由于不定根的存在,在早晨仍然能保持与对照相同的光能转化效率,而在光下遭受切断干扰胁迫的臭柏小枝,维持高的光能转化效率的能力下降。
|
|
臭柏小枝8月24日气孔导度的日进程(表 5)表明,对照气孔导度在12:00前的变化迅速,8:3 0处于最高值,随后开始下降,12:00后变化趋于平缓,直到19:00均没有回升。切断处理样枝的气孔导度虽然也表现出与对照相似的变化趋势,但其强度和下降速度均低于对照。在8: 30切断处理的气孔导度仅为对照的30%左右,以后随着时间的推移两者的差距逐渐缩小,12: 00切断处理的小枝(叶)的气孔导度约为对照的36%,12:00后切断处理的样枝的气孔导度较对照平均约低39%。对照和切断匍匐茎处理小枝的气孔导度的差异性检验结果(表 5)表明, 18:00前二者差异显著(P<0.05),切断处理明显地抑制了臭柏刺叶的气孔导度。
|
|

植物叶片捕获的激发能,一部分用于CO2的固定,另一部分以荧光或热的形式耗散掉。因此植物发射的叶绿素a荧光成为一种反映光合作用活性和行为的指标(赵平等,2000;陈贻竹等,1995)。在叶绿素a荧光参数中,叶绿素荧光的可变部分(Fv)与最大荧光值(Fm)的比值(Fv/Fm),反映了开放的PSⅡ反应中心捕获激发能的效率,是研究植物胁迫的重要参数,任何影响PSⅡ效能的环境胁迫均会使Fv/F m降低(李敦海等,2000)。在未受环境胁迫的条件下,高等植物叶片的Fv/Fm为0.80~0.83(Bjökman et al., 1987)。臭柏匍匐茎切断处理4 d后Fv/Fm(8月19日测定)平均值为0.781±0.009,对照小枝的Fv/Fm(8月19日测定)平均值为0.782±0.008,表明遭受切断干扰的臭柏匍匐茎的PSⅡ反应中心仍能保持良好的最大光能转化效率。ΔF/F′m是反映在光下PSⅡ反应中心有部分关闭时的实际光能捕获效率的指标,弱光下的植物因叶面入射光量少而限制其对光能的吸收,但其较高的ΔF/F′m值被认为可以部分地减少光能不足的不利效应(林植芳等,2000)。臭柏匍匐茎切断处理和对照枝的ΔF/F′m在中午时降低,出现光抑制,而后随光强减弱又恢复,表明这种调节是可逆的。NPQ是反映非光化学猝灭如热耗散的有效指标,高的NPQ可以防止过量的光能造成光合器官失活或破坏,已有研究表明NPQ与叶黄素循环有关(Demmig-Adams, 1990;姜闯道等,2002)。8月24日臭柏匍匐茎切断处理和对照枝在上午8:30的PSⅡ电子传递的量子效率较高(ΔF/F′m分别约为0.27、0.38),非光化学猝灭(NPQ)较低,通过PSⅡ的电子传递速率高达86.3和90.1 μmol·m-2s-1(表 3),表明吸收的光能用于光化学反应的比例高,PSⅡ已处于活跃的状态,这种状态保持到10:00左右。中午,NPQ的上升和ETR有所下降,表明叶绿体吸收的光能用于实际的光化学转换的比例减少,而用于非光化学反应的耗散能量的比例增大。臭柏匍匐茎切断处理样枝较对照样枝受到的光抑制作用较大。
干扰通过改变资源可利用性影响植物的许多功能,从而导致植物行为发生明显改变(Bazzaz, 1996)。切断匍匐茎干扰处理的实质是阻断了母株向子(分)株的水分、矿质元素等的运输,改变原有的资源利用方式,对匍匐茎地上小枝的PSⅡ功能产生了明显影响。在干扰处理的24 h内,这种影响已在反映PSⅡ功能特性的荧光参数变化中得到体现,表现为ΔF/F′m和ETR下降,NPQ上升。同冯建灿等(2002)报道的自然干旱胁迫下,刺槐(Robinia pseudoacacia)叶绿素a荧光参数Fv/Fm下降,NPQ上升,施用土壤保水剂可以减少叶绿素a荧光参数的下降程度的结果一致。同何维明等(2000)报道的水分梯度对臭柏光能吸收、传递、最大转化效率的影响均不显著的结果不一致,主要原因可能是土壤含水率差异大所致。何维明等(2000)报道的水分梯度中土壤的最低含水率为4.08%,而本研究测定时沙地风沙土的含水率为2.68%。但这些研究结果得出一个共同结论是土壤水分条件,首先直接影响植物叶绿素a荧光参数,进而影响植物的生长发育。在切断匍匐茎干扰处理9 d后(表 3),臭柏匍匐茎切断处理样枝光合电子传递速率(ETR)的日均值为对照的79.76%,ΔF/F′m为对照的81.56%,NPQ为对照的125.20%,表明此时切断匍匐茎干扰处理的小枝受到较对照枝更强的水分胁迫。切断与上一级分株的连接后,匍匐茎上小枝荧光参数的这种变化,有利于该匍匐茎上小枝在受到水分胁迫时有效地保护光合机构不致遭受严重伤害,也是分株在生理特性上对切断干扰的一种响应。
切断匍匐茎后,匍匐茎上小枝的气孔导度下降,午前下降幅度较午后明显(表 4),与叶绿素a荧光参数的变化趋势相近似。切断分株的气孔导度下降,可能也是适应水分供给源减少的一种生理对策。8月19日测定的切断和对照枝黎明前的Fv/Fm值非常接近,表明承受切断处理分株在经过夜间的恢复后,PSⅡ活性可以达到相当于对照的水平,但白天随着光照、水分胁迫的加大及气孔开放程度的减小,受到比对照更加严重的胁迫,各荧光参数间的差异较大;从12:00直到19:00,二者的气孔导度差距变小,回升速度缓慢。
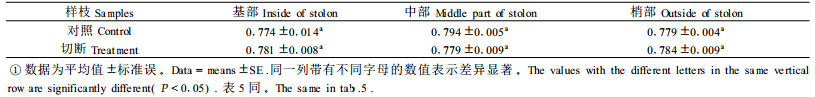
臭柏匍匐茎在遇到沙埋后可产生不定根,并以这种克隆生长的方式进行个体拓殖。在干扰频繁的环境中,克隆生长对克隆植物的生存有重要作用(董鸣,1999;何维明,2000)。相连的分株当处于异质性资源供应时,可发生资源传输,这种传输可以是向顶性的也可以是向基性的(Alpert, 1996;Headley et al., 1988)。切断臭柏匍匐茎与上级分株的连接后,匍匐茎资源供应状况发生改变,处于匍匐茎基部的地上小枝失去了来自上级分株的水分供给,可能会首先面临水分胁迫,但8月19日测定结果显示,切断处理样枝基部、中部、梢部的Fv/Fm值与对照没有显著差异(表 4),表明匍匐茎中部不定根吸收的水分在整个匍匐茎中实现了共享,这种共享既是向顶性的也是向基性的。资源异质性是自然环境的基本特征,克隆植物的资源共享特性可以缓冲局域的资源短缺与环境胁迫,这对臭柏群落的维持与拓展具有非常重要的生态学意义。
切断匍匐茎干扰处理后,由于匍匐茎中部有不定根的存在,被切断匍匐茎上小枝的ΔF/F′m、ETR在切断一周后仍能保持在对照的80%左右(各时刻平均),不致使光合机构被迅速破坏,不定根在消除切断匍匐茎所造成的不利干扰中,发挥了明显的作用。同何维明(2000)切断匍匐茎对沙地柏(Sabina vulgaris)子株存活率影响不显著的报道一致,表明不定根在维持臭柏群落稳定和拓展中具有重要作用。尽管如此,切断匍匐茎干扰处理还是导致了切断匍匐茎的PSⅡ反应中心功能特性的下降,切断匍匐茎的PSⅡ反应中心功能是否会因匍匐茎上不定根数量和空间的增加而恢复?多少不定根量能够抵御类似的干扰?这些问题仍需进一步研究。
陈贻竹, 李晓萍, 夏丽, 等. 1995. 叶绿素荧光技术在植物环境胁迫研究中的应用. 热带亚热带植物学报, 3(4): 79-86. |
陈晓英, 姜闯道, 邹琦, 等. 2002. 类囊体的空间结构及其对激发能分配的调控. 植物生理学通讯, 38(4): 307-311. |
董鸣. 1999. 切断根茎对根茎禾草沙鞭和赖草克隆生长的影响. 植物学报, 41(2): 194-198. DOI:10.3321/j.issn:1672-9072.1999.02.017 |
冯建灿, 胡秀丽, 苏金乐, 等. 2002. 保水剂对干旱胁迫下刺槐叶绿素a荧光动力学参数的影响. 西北植物学报, 22(5): 144-149. |
何维明. 2000. 切断匍匐茎对沙地柏子株生长和资源利用效率的影响. 植物生态学报, 24(4): 391-395. DOI:10.3321/j.issn:1005-264X.2000.04.002 |
何维明, 马风云. 2000. 水分梯度对沙地柏幼苗荧光特征和气体交换的影响. 植物生态学报, 24(5): 630-634. DOI:10.3321/j.issn:1005-264X.2000.05.021 |
姜闯道, 高辉远, 邹琦. 2002. 缺锰降低大豆叶片叶绿素荧光的高能态猝灭. 植物生理与分子生物学学报, 28(4): 287-291. |
李敦海, 宋立荣, 刘永定. 2000. 念珠藻葛仙米叶绿素荧光与水分胁迫的关系. 植物生理学通讯, 36(3): 205-208. |
林植芳, 彭长连, 孙梓健, 等. 2000. 4种木本植物叶片的光合电子传递和吸收光能分配特性对光强的适应. 植物生理学报, 26(5): 387-392. DOI:10.3321/j.issn:1671-3877.2000.05.003 |
林植芳, 彭长连, 林桂珠. 1999. C3、C4植物叶绿素荧光猝灭日变化和对光氧化作用的响应. 作物学报, 25(3): 284-290. DOI:10.3321/j.issn:0496-3490.1999.03.003 |
苏行, 胡迪琴, 林植芳, 等. 2002. 广州市大气污染对两种绿化植物叶绿素荧光特性的影响. 植物生态学报, 26(5): 599-604. DOI:10.3321/j.issn:1005-264X.2002.05.013 |
王林和, 刘美珍, 张国盛, 等. 2002. 毛乌素沙地臭柏不定根发生特性的研究. 林业科学, 38(5): 156-159. DOI:10.3321/j.issn:1001-7488.2002.05.026 |
王林和, 张国盛, 董智. 1998. 毛乌素沙地臭柏种子产量及更新的初步研究. 林业科学, 34(6): 105-112. DOI:10.3321/j.issn:1001-7488.1998.06.014 |
吴长艾, 孟庆伟, 邹琦. 2001. 叶黄素循环及其调控. 植物生理学通讯, 37(1): 1-4. |
肖春旺, 周广胜. 2001. 毛乌素沙地中间锦鸡儿幼苗生长、气体交换和叶绿素荧光特性对模拟降水量变化的响应. 应用生态学报, 12(5): 692-696. DOI:10.3321/j.issn:1001-9332.2001.05.011 |
杨胜铭, 高辉远, 邹琦. 2001. 状态转换对光合作用中激发能分配的调节及其与光破坏防御的关系. 植物生理学通讯, 37(2): 89-93. |
张国盛, 高润宏, 王林和, 等. 2001. 毛乌素沙地臭柏群落结构和生物多样性组成研究. 内蒙古农业大学学报, 22(4): 88-91. |
张国盛, 王林和, 李玉灵, 等. 1999. 毛乌素沙地臭柏根系分布及根量. 中国沙漠, 19(4): 378-383. DOI:10.3321/j.issn:1000-694X.1999.04.013 |
张教林, 曹坤芳. 2002. 光照对两种热带雨林树种幼苗光合能力、热耗散和抗氧化系统的影响. 植物生态学报, 26(6): 639-646. DOI:10.3321/j.issn:1005-264X.2002.06.001 |
张守仁. 1999. 叶绿素荧光动力学参数的意义及讨论. 植物学通报, 16(4): 444-448. DOI:10.3969/j.issn.1674-3466.1999.04.021 |
张新时. 1994. 毛乌素沙地的生态背景及其草地建设的原则与优化模式. 植物生态学报, 18(1): 1-16. DOI:10.3321/j.issn:1005-264X.1994.01.008 |
赵平, 孙谷畴, 曾小平, 等. 2000. 两种生态型榕树的叶绿素含量、荧光特性和叶片气体交换日变化的比较研究. 应用生态学报, 11(3): 327-332. DOI:10.3321/j.issn:1001-9332.2000.03.002 |
山中典和, 张国盛, 王林和, 等. 2001. 中国内蒙古毛乌素沙地における看护植物の除去が臭柏幼树の生存におよぼす影响. 日本绿化工学会志, 26(1): 120-124. |
温国胜, 图子美和子, 坂本圭儿, 等. 2000. 慢性な水ストレス下での臭柏の生育经过. 日本绿化工学会志, 25(3): 242-248. |
Alpert P. 1996. Nutrient sharing in natural clonal fragments of Fragaria chiloensis. J Ecology, 84: 395-406. DOI:10.2307/2261201 |
Bazzaz F A. 1996. Plants in changing environments: linking physiological, populations, and community ecology. London: Cambridge University Press, 14-37.
|
Bjökman O, Demming B. 1987. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77K among vascular plants of diverse origins. Planta, 170: 489-504. DOI:10.1007/BF00402983 |
Demmig-Adams B. 1990. Carotenoids and photoprotection in plants: a role for the xanthophyll zeaxanthin. Biochim Biophys Acta, 1020: 1-24. DOI:10.1016/0005-2728(90)90088-L |
Genty B, Briantais J M, Baker N R. 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim Biophys Acta, 900: 87-92. |
Headley A D, Callaghan T V, Lee J A. 1988. phosphate and nitrate movement in the clonal plants Lycopodium annotinum L. and Diphasiastrum complanatum(L.) Holub. New Phytol, 110: 487-495. DOI:10.1111/j.1469-8137.1988.tb00287.x |
Hirobe M, Ohte N, Karasawa N, et al. 2001. Plant species effect on the spatial patterns of soil properties in the Mu-us desert ecosystem, Inner Mongolia, China. Plant and Soil, 234: 195-205. DOI:10.1023/A:1017943030924 |
Schreiber U, Bilger W, Neubauer G. Chlorophyll fluorescence: New instruments for special applications. In: Schulze E D, Caldwell M M. Ecophysiology of photosynthesis. Berlin: Springer-Verlag, 1994
|