 2004, Vol. 40
2004, Vol. 40文章信息
- 施积炎, 丁贵杰, 袁小凤.
- Shi Jiyan, Ding Guijie, Yuan Xiaofeng.
- 不同家系马尾松苗木水分参数的研究
- Studies on Water Parameters in Pinus massoniana Seedlings of Different Families
- 林业科学, 2004, 40(3): 51-55.
- Scientia Silvae Sinicae, 2004, 40(3): 51-55.
-
文章历史
- 收稿日期:2002-10-25
-
作者相关文章



2. 浙江中医学院生命科学系 杭州 310053
2. Life Science Department of Zhejiang TCM Hangzhou 310053
研究树木对干旱胁迫的适应性,探索耐旱机理,进而培育耐旱且高产品种、家系或无性系,是提高树木生产潜力,摆脱干旱胁迫的最基本途径。李吉跃(1988)和郭连生(1989)将P-V技术引入树木耐旱性研究,标志着我国树木耐旱性研究进入了一个新的阶段。树木耐旱性方面作了大量研究,但大都比较不同树种间的耐旱性差异,对于种内耐旱性遗传变异研究较少。马尾松(Pinus massoniana)是优质建筑、纤维、松香(脂)材,也是我国南方主要工业用材和造林灭荒树种。本文应用P-V技术对不同家系马尾松的渗透势(φstlp)、充分膨胀的渗透势(φssat)、膨压为0时的相对含水量(RWCtlp)、膨压为0时的相对渗透含水量(ROWCtlp)、细胞最大弹性模量(εmax)和膨压随叶水势下降而下降的速率(b)等水分参数进行了研究,并对各家系渗透调节和保持膨压的能力作了综合评价,探讨马尾松的耐旱适应性及种内不同家系耐旱性遗传变异,筛选耐旱性相对较强的优良家系,从而为马尾松造林的家系选择、立地选择和培育技术提供理论依据和现实指导。
1 材料与方法 1.1 试验材料1999年1月选用1个贵州本地台江种源(作对照)和9个广西优良家系(家系号分别为20、23、38、75、83、127、186、456和464)马尾松1a生苗(同一种苗木其形态基本一致)在温室中进行盆栽。盆栽土壤为熟土(70%)+黄土(20%)+火烧土(10%)混匀而成,密度为1.08Mg·m-3。盆子规格为:高30cm,直径32cm。待苗长至2000年5月开始进行试验。
1.2 试验方法 1.2.1 供水良好时不同家系水分参数在供水良好时(土壤含水量SWC均高于26%),应用ZLZ-4型植物水分状况测定仪(压力室)测定叶水势和制作P-V曲线,各指标测定3次。具体方法详见Tyree(1972)和王万里(1984)。最大弹性模量εmax的计算采用沈繁宜等(1994)的新方法。采用SPSS软件进行数据分析。
1.2.2 干旱胁迫下水分参数的变化选择83和464家系进行研究。马尾松苗木在干旱处理前,充分浇水3d(土壤水分基本处于饱和状态),以后不再浇水,使其自然干燥,每隔3d取土样测定土壤含水量和取枝叶进行指标测定,每个处理3个重复。叶水势测定和P-V曲线的制作同上。
|
|
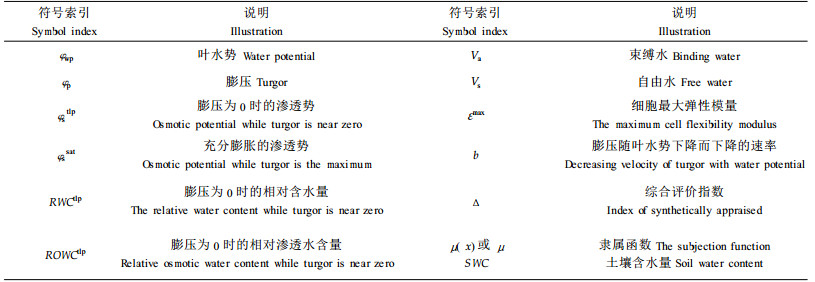
春夏之际是树木展叶和嫩枝生长、耐旱能力最弱的时期,这一时期耐旱性强弱对树木当年的生长和存活有决定性的作用,因此本试验重点对这一时期不同家系马尾松苗木的水分参数进行了研究。
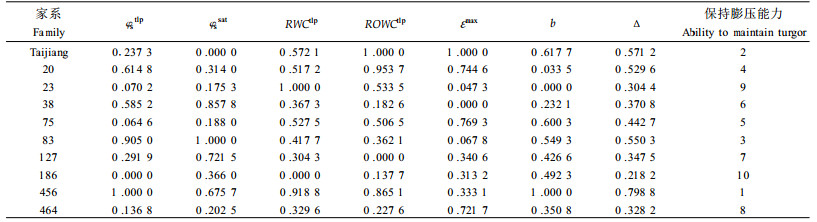
φstlp值反映了树木维持最低膨压的极限渗透势。Tyree(1972)曾指出植物耐旱特性表现在组织的细胞出现质壁分离时具有一个大的整体渗透势(φstlp)和一个高的整体弹性模量(ε)。φstlp越低,表明植物维持膨压的能力越强,所以φstlp值的大小被公认为是衡量植物耐旱性强弱的最佳指标之一(张建国等,1994a)。根据φstlp的排序(表 2),供试家系保持膨压的能力为:456>83>20>38>127>台江>464>23>75>186。不同家系φssat大小排序与φstlp的排序有差异,总体看,保持膨压能力强的家系,φssat值较低,而保持膨压能力弱的家系φssat值较高。植物细胞的φssat主要决定于2个方面:一是细胞中可溶性溶质的数量和种类;二是细胞体积的变化,主要是指细胞中自由水与束缚水的变化(张建国等,1994b)。
|
|
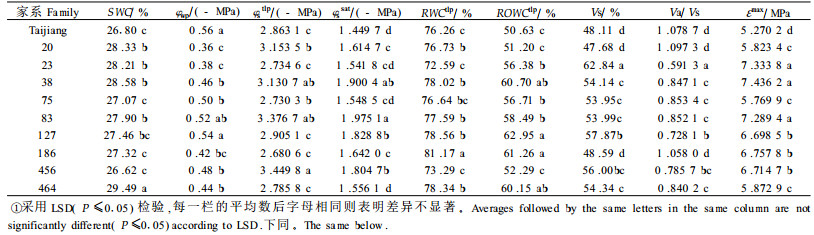
RWCtlp、ROWCtlp是组织细胞初始质壁分离时的相对含水量和相对渗透水含量。RWCtlp、ROWCtlp值越低,表明细胞在很低的含水量下才发生质壁分离。所以这2个指标在一定程度上反映了组织细胞对脱水的忍耐能力。从表 2可以看出,23家系与456家系的RWCtlp值最低,分别为72.59%和73.29%,186家系RWCtlp值则相对较高,为81.17%。但仅根据RWCtlp值还不能准确判定不同家系苗木组织细胞对脱水的忍耐能力大小,植物或家系不同,束缚水和自由水的含量比例不同,其组织细胞初始质壁分离时的相对含水量也就不一致,又因为植物组织细胞代谢所利用的水主要为渗透水,所以认为ROWCtlp值的大小能客观地反映植物组织细胞对脱水的忍耐能力。台江种源、家系20和家系456具有较低的ROWCtlp值,因此其组织细胞具有较强抗脱水能力。而186、464和38家系ROWCtlp值较高,其组织细胞抗脱水能力也相对较弱。
Va/Vs表示苗木束缚水和自由水含量之比。各家系的Va/Vs都很高,除23家系Va/Vs为0.591 3外,其余家系Va/Vs都在0.728 1以上,比一般阔叶树高很多。一般认为Va/Vs越高,表明细胞原生质粘滞性及原生质胶体的亲水性越强,因而越有利于树木吸水和保持体内水分,对提高树木的耐旱能力有重要意义(李吉跃等,1997)。因此,从Va/Vs看,这些家系具有很强的保水能力,但在干旱胁迫下Va/Vs的提高究竟对耐旱性起多大的作用,很难准确定论。此外,Va/Vs还是影响渗透调节的一个因素,所以与植物保持膨压有关。
2.3 不同家系细胞弹性模量(εmax)的比较树木叶组织的细胞弹性大小影响着细胞膨压的变化。叶组织的细胞弹性可以用细胞弹性膜量(ε)来表示。ε值越大,表示细胞壁越坚硬,弹性越小,反之,则说明细胞越柔软,弹性越大。通常ε被定义为组织在失去一部分水分后细胞部分压力势的变化:ε=V×(dP/dV)。式中,V是细胞自由水体积,P是压力势(李吉跃等,1997;刘友良,1992)。
由于ε不是一个常数,所以在分析中一般取最大体积弹性模量εmax表示细胞弹性大小来揭示树木在干旱胁迫下的适应能力。研究表明,在正常水分条件下,各马尾松家系εmax都很小,为5.270 2~7.333 8 MPa,说明马尾松细胞有比较大的弹性,因而保持膨压的能力比较强,其中台江种源的最小,为5.270 2 MPa,显示出细胞有比较大的弹性。而38家系的值最大,为7.333 8 MPa,细胞弹性相对较小。
2.4 不同家系膨压(φp)与叶水势(φwp)关系当植物含水量下降而引起水势下降时,组织细胞能够保持膨压就是一种重要的耐旱机理(李吉跃等,1994),这对维持植物的生长和生存以及正常的生理功能都是至关重要的。研究表明,10个家系苗木膨压φp随叶水势φwp下降而下降(表 3),并且有良好的线性关系(φp=a+b φwp),而且这种线性关系十分稳定,不随土壤干旱胁迫的加剧而发生变化。在线性方程中,参数a值表示树木充分膨胀时叶子细胞能够达到的最大膨压,参数b值则反映树木渗透调节能力的大小,b值越小,表明随着水势的下降,膨压下降的速率越小,因而保持膨压的能力也就越强。b值为1时,表明树木没有渗透调节能力。
|
|
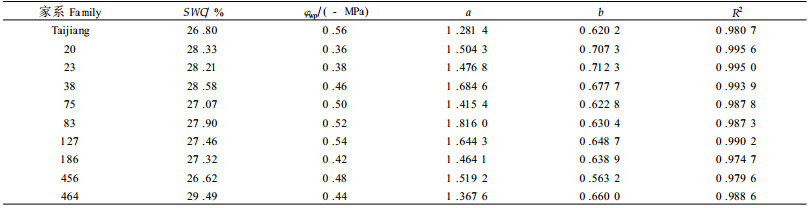
从表 3可以看出,各家系b值都小于0.712 3,说明所有供试马尾松家系都有一定的渗透调节能力,但家系间差异较大。依据b值的大小,10个家系渗透调节能力由大到小的顺序为:456>台江>75>83>186>127>464>38>20>23。应指出b值的排序(表 3)和φstlp的排序(表 2)并不一致,这是因为b值的大小除了与φstlp有关外,还与细胞弹性以及φssat等因素有关。
2.5 不同家系保持膨压能力综合评价为了较为准确地评价不同家系马尾松间耐旱性的差异,选取了φstlp、φssat、RWCtlp、ROWCtlp、εmax和b值6个水分参数指标,对供试家系苗木保持膨压能力进行了综合评价。具体方法是将水分参数隶属函数数值化,这样既可使水分参数无量纲化,又可对水分参数在保持膨压方面的贡献直接度量,由于参加评价的上述6个水分参数与保持膨压的能力呈负相关关系,所以用反隶属函数的方法来计算每个水分参数在保持膨压方面的贡献大小。其具体计算公式为(张建国等,1994b;陈贻源,1992):μ(xj)=1-(xj-xmin)/(xmax-xmin),式中,xj表示水分参数,μ(xj)∈[0,1];综合评价结果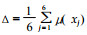
各家系保持膨压能力综合评判结果见表 4。由于树木耐旱能力的强弱不是一个绝对概念,其排序也只是相对而言,尤其对于同一树种的不同家系(或种源),它们的耐旱能力非常相近,因此,根据耐旱能力的强弱,把家系划分成若干类群更有实际应用价值。聚类分析结果(图 1)表明:所有家系可明显分成3类,第1类包括456、20家系和台江种源,是保持膨压能力最强的类群;第2类包括83、127、38家系,是保持膨压能力较强类群;第3类包括23、186、464和75家系,是保持膨压能力相对较弱类群。
|
|

|
图 1 不同家系保持膨压能力系统聚类图 Fig. 1 System cluster of φp capacity from different families |
通过83、464家系(限于篇幅,仅取2个比较)的对比研究(表 5)表明,干旱胁迫下,φstlp、RWCtlp、ROWCtlp和b值总体呈先下降后上升的趋势。说明在一定的干旱胁迫范围内,马尾松苗木的渗透调节能力随着干旱胁迫的发展渗透调节能力提高了。b值的下降无疑有助于苗木在叶水势降低的情况下保持膨压,维持正常的生理功能。从b值的变化可以看出,马尾松苗木渗透调节能力最强的时期可能出现在中度至严重胁迫范围(-1.6~-2.5 MPa)。2个家系εmax值在干旱胁迫下都有一定程度的上升,其中83家系升幅较小,而464家系在-2.42 MPa时上升幅度突然变大,说明83家系比464家系对干旱胁迫的适应能力相对较强。
|
|
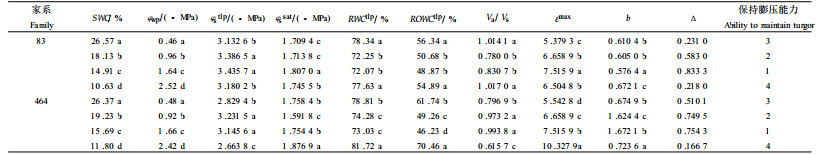
从不同水势梯度下保持膨压能力综合评价结果看(评价方法同上),2个家系苗木保持膨压的能力都表现为先上升后下降的趋势,说明在一定干旱胁迫范围内,马尾松苗木的渗透调节和保持膨压的能力逐渐得到提高,表现出较强的耐旱能力。这是马尾松苗木对干旱胁迫的一种适应性,也为干旱锻炼方法提高树木的耐旱能力提供了理论依据。目前已知许多木本植物都具有渗透调节能力,但不同植物及其品种渗透调节能力不同,参与渗透调节的物质种类不同。总之,渗透调节是一个很复杂的问题,受许多因素的影响,水分亏缺速率、发育状况、环境因子、水分亏缺的前期历史等都会影响到植物的渗透调节。植物以什么方式或途径适应干旱,还取决于植物本身的生理生态特性和具体的条件。
3 小结φstlp值的大小被公认为是衡量植物耐旱性强弱的最佳指标之一。根据φstlp的排序,供试家系保持膨压能力的排序为:456>83>20>38>127>台江>46 4>23>75>186。各马尾松家系εmax都很小,为5.270 2~7.333 8 Mpa,说明马尾松细胞有比较大的弹性。台江种源、20和456家系具有较低的ROWCtlp值,因此其组织细胞抗脱水能力较强。而186、464和38家系ROWCtlp值较高,其组织细胞抗脱水能力也相对较弱。各家系的Va/Vs都很高,显示出较强的保水能力。各家系苗木膨压φp随叶水势φwp下降而下降,并且有良好的线性关系(φp=a+b φwp)。各家系b值都小于0.712 3,说明所有供试马尾松家系都有较强的渗透调节能力。
综合评价表明:456、83、20家系和台江种源渗透调节和保持膨压的能力最强,75、38和127家系渗透调节和保持膨压的能力较强,而23、464和186家系渗透调节和保持膨压的能力则相对较弱。说明马尾松苗木耐旱性存在较大的种内遗传变异。
干旱胁迫下,对83家系与464家系的研究表明,在一定水势范围,b值出现一个低值,φwtlp也有一定程度的下降,εmax值有一定程度的上升。83家系对干旱的适应能力强于464家系。在一定的干旱胁迫范围内,马尾松苗木随着干旱胁迫的加剧渗透调节能力有所提高。
陈贻源编. 模糊数学. 武汉: 华中工学院出版社, 1992: 41-51
|
郭连生. 1989. 对几种阔叶树种耐旱性生理指标的研究. 林业科学, 25(5): 389-394. DOI:10.3321/j.issn:1001-7488.1989.05.004 |
李吉跃. 1988. 油松侧柏苗木抗旱性初探. 北京林业大学学报, 10(2): 23-30. |
李吉跃, 张建国, 姜金璞. 1994. 京西山区人工林水分参数的研究(Ⅱ). 北京林业大学学报, 16(2): 1-9. |
李吉跃, 张建国, 姜金璞. 1997. 侧柏种源耐旱特性及其机理研究. 林业科学, 33(sp1): 1-13. |
刘友良. 1992. 植物水分逆境生理. 北京: 农业出版社.
|
沈繁宜, 李吉跃. 1994. 植物叶组织弹性模量新的计算方法. 北京林业大学学报, 16(1): 35-39. |
王万里. 1984. 压力室(PRESSUS CHAMBER)在植物水分状况研究中的应用. 植物生理学通讯, (3): 52-57. |
张建国, 李吉跃, 姜金璞. 1994a. 京西山区人工林水分参数的研究(Ⅰ). 北京林业大学学报, 16(1): 1-12. |
张建国, 李吉跃, 姜金璞. 1994b. 京西山区人工林水分参数的研究(Ⅲ). 北京林业大学学报, 16(4): 46-54. |
Tyree M T. 1972. The measurement of the turgor pressure and water relations of plants by pressure-bomb technique. J Exp Bot, 23: 267-282. DOI:10.1093/jxb/23.1.267 |