 2004, Vol. 40
2004, Vol. 40文章信息
- 张明如, 翟明普, 王学勇, 贾黎明, 沈应柏.
- Zhang Mingru, Zhai Mingpu, Wang Xueyong, Jia Liming, Shen Yingbai.
- 火炬树克隆植株生长和生物量特征的研究
- A Study on the Characteristics of the Growth and the Biomass of Clonal Ramets in Rhus typhina
- 林业科学, 2004, 40(3): 39-45.
- Scientia Silvae Sinicae, 2004, 40(3): 39-45.
-
文章历史
- 收稿日期:2003-04-11
-
作者相关文章


2. 河北林业科学研究院 石家庄 050061
2. Hebei Academy of Forestry Shijiazhuang 050061
引进外来树种的初衷是为了丰富当地的物种资源、提高生物生产力、更有效地改善环境的生态质量,但是由于少数树种的生物学、生态学特性独特,在缺乏与当地植物、动物和非生物环境协同进化的前提下,很难形成和谐的生态关系,往往演变为外来入侵种,排挤当地的植物种,导致生物多样性锐减,危及自然生态系统的结构和功能(徐承远等,2001)。在我国引种的桉树(Eucalyptus spp.)、大米草(Spartina alterniflora)、水葫芦(Eichhornia crassipes)、薇甘菊(Mikania micrantha)等产生的生态后果(李振宇等,2002;解炎,2002)就是具体的佐证。目前太行山低山丘陵区正在进行退耕还林还草的生态工程建设,在实践中存在树种单一、大比例栽植外来树种、整地整齐划一等问题,其中大量种植推广的木本植物之一就有外来种火炬树(Rhus typhina)。
火炬树原产美国和加拿大南部,耐旱、耐瘠薄、喜光,幼期生长迅速,繁殖材料极易解决,被认为是水土保持的优良树种,秋季形成的景观优美独特(孙立元等,1997),因此火炬树在荒山植被恢复中被广泛选用。目前火炬树已在主要引种栽植地区如北京、河北和山东等不断向四周蔓延扩散,占据灌丛草地,并向农田侵入。火炬树克隆繁殖形成居于绝对优势地位的单优群落已经影响当地种构成落叶灌木层片和草本层片在群落中分布,导致物种的饱和度降低。
目前已有对火炬树苗木的耐旱性、苗木培育、分泌道的发育解剖学、腺毛的形态结构和发育、组织培养、侧柏火炬树混交林水源涵养功能、蚂蚁为火炬树传粉和幼苗幼树的分布格局等的研究(李吉跃等,1992;马淑英等,1997;毛学文等,1997;段新玲等,2000;庞福生等,2001;Werner et al., 1982;Gelderloos, 1988)。在自然条件下具有克隆生长(即无性繁殖相伴的营养生长过程)生物学过程的植物被称为克隆植物(clonal plant)(董鸣,1996)。自然条件下利用根系器官完成克隆繁殖过程的木本植物有山杨(Populus davidiana)(徐化成等,1994)、刺槐(Robinia pseudoacacia)、酸枣(Zizyphus setiva var.spinosus)、荆条(Vitex chinensis)、珍珠梅(Sorbaria sorbifolia)、漆树(Rhus verniciflua)、构树(Broussonetia papyrifera)、沙棘(Hippophe rhamnoides)(李根前等,2001)、火炬树和竹类(Phyllostachys spp.)(宋明华等,2002)等。国内外有关学者都注意到火炬树根蘖扩散能力极强的现象(潘志刚等,1994;Donald et al., 1998;James et al., 2001),但尚未研究火炬树克隆繁殖的特点。本文测定研究其根系生长发育基本特点和生物量积累时空格局,并分析与生境类型或群落类型的关系,揭示外来树种火炬树克隆繁殖扩散的基本规律。
1 材料与方法 1.1 调查地区概况研究地区位于河北省平山县太行山丘陵低山区,海拔150~400 m,分布有宽度不等的沟谷。丘陵地势较缓,而山地地势相对陡峻。地带性气候属暖温带半干旱半湿润季风型大陆性气候。土壤以褐土为主,沟谷和坡下部土层深厚,随着坡位上升土层变薄;pH值为6.5~7.5,有机质含量为1%左右,土壤速效氮约21.4~80 mg·kg-1, 土壤速效磷含量约5.4~16 mg·kg-1, 土壤速效钾约60~103 mg·kg-1,成土母岩以片麻岩为主。
天然植被因人为干扰程度和地形因子的复合影响而发生逆行演替,现存植被主要由荆条灌丛、酸枣灌丛和分布于缓坡沟谷地带梯田的农作物、经济林以及散生树木、沟谷的阔叶林带、果园构成。由于人为过度利用,灌丛与杂草群丛低矮稀疏,部分地段表土侵蚀严重。
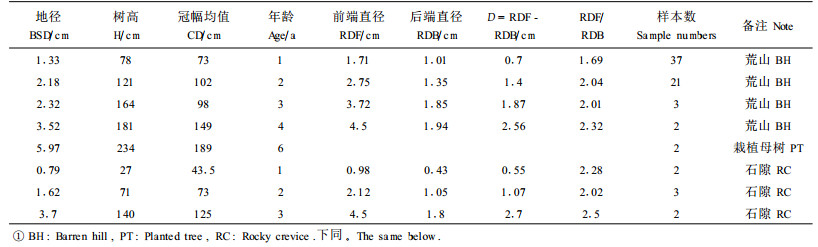
1.2 研究方法研究区于1997年开始栽植火炬树,多数克隆子株为1~2 a,且呈群团状分布。2002年7—9月分别生境类型和年龄,随机选择火炬树克隆繁殖子株测定其前后端水平侧根直径(67株)、单位长度(以10 cm为单位)水平侧根鲜重(31株)和采用全树收获法(30株)分干、枝、叶、皮、粗根、中根、小根、细根(董鸣,1997)称取鲜重以及测定调查地径、树高、枝下高、冠幅、年龄、胸径和起源;将水平侧根样品和全树样品带回室内在85℃烘箱内烘48 h, 然后在精度0.001 g天平称重。其中火炬树克隆子株前端水平侧根和后端水平侧根定义为火炬树水平侧根上子株成苗处的前部侧根和后部侧根(图 1);由于火炬树克隆子株一级水平侧根长度的不确定性,为了便于对比,本文采用子株前后端10 cm长的水平侧根作为比较测定指标。

|
图 1 火炬树克隆子株前端水平侧根直径(B)与后端水平侧根直径(A)对比 Fig. 1 Comparison the front part of the lateral root diameter(RDF, B) and the back part of the lateral root diameter(RDB, A)of the Staghorn sumac clonal ramets |
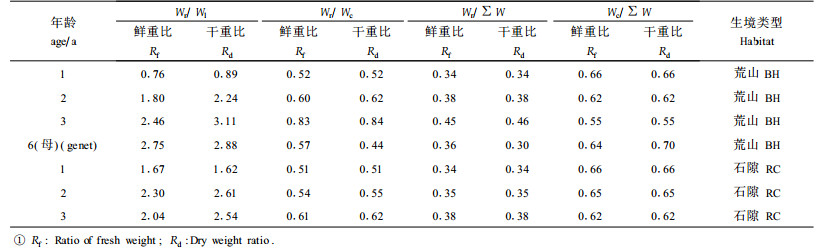
在调查测定的基础上,进一步计算火炬树克隆子株前端水平侧根直径(RDF)与后端水平侧根直径(RDB)比值Rd、克隆子株前端水平侧根直径与后端水平侧根直径的差值为分化度D、克隆子株前端单位长度水平侧根生物量(W1)与后端单位长度水平侧根生物量(W2)比值为Rw、克隆子株前端单位长度水平侧根生物量(W1)与后端单位长度水平侧根生物量(W2)的差值为分化度Dw、克隆子株根系生物量(Wr)与树冠生物量(Wc)的比值(即Wr/Wc)和克隆子株根系生物量(Wr)与叶片生物量(Wl)的比值(Wr/Wl)。而LRBF和LRBB分别表示克隆子株前端和后端单位长度水平侧根生物量的干重。
2 结果与分析火炬树在原产地具有典型的克隆繁殖特征,引种到我国温带和暖温带地区同样表现出克隆繁殖的特点。火炬树栽植1年后,开始出现克隆子株。自然条件下克隆子株可生长分布在荒山草坡、石质山地和农田等生境,而且随时间推移,克隆子株数量迅速增加,向周围扩散并占据临近空间成为优势种。
2.1 火炬树根系形态特征火炬树栽植后其主根生长缓慢最终停止,而水平侧根不断向前延伸,通常分布在土壤表层10 ~20 cm的范围内。根系分布浅利于水平侧根不定芽萌动出土。克隆子株前端水平侧根通常为1条,有时分叉为2~3条或更多。独特的是克隆子株前后端水平侧根直径差异显著(图 1),前端水平侧根直径远远超过后端水平侧根直径,克隆子株前后端水平侧根直径呈现明显的不对称性。随着火炬树水平根继续向前生长,产生多级克隆子株;与后端水平侧根直径相比,每一子株前端水平侧根直径明显增大,这为水平侧根向前延伸和子株不断形成奠定营养基础。火炬树克隆子株前后端水平侧根直径差异显著,在荒山草坡、农田、石隙、路旁和平缓、上坡、下坡地段及随火炬树年龄变化几乎表现完全一致的形态变异格局。
2.2 火炬树克隆子株前后端水平侧根直径的变化规律根据植物生长与繁殖是依赖于净初级生产力的原理(孙儒泳等,1993),本文推测火炬树克隆子株前后端水平侧根直径差异是由于光合有机物质在水平侧根前后端积累不平衡的结果。显然火炬树克隆子株前后端水平侧根直径与其地径、冠幅、年龄和树高存在一定的相关性。
2.2.1 火炬树地径与水平侧根直径的关系随地径增加,克隆子株前后端水平侧根直径必然会增大。在调查范围内火炬树克隆繁殖产生的子株大多数为1 a或2 a,所以仅分析火炬树1 a和2 a子株地径与其水平侧根直径的关系。由表 1可知, 无论是1 a还是2 a克隆子株,子株前端地径与前端水平侧根直径的复相关系数均大于地径与后端水平侧根直径的复相关系数,说明火炬树的光合产物用于根茎和子株前端水平侧根的生长远超过子株后端水平侧根的生长,所以地径和前端水平侧根直径较大,而后端水平侧根直径较小。
|
|

通过统计相关分析,结果同样发现(表 1),1 a幼苗和2 a幼树的冠幅与前端水平侧根直径复相关系数大于冠幅与后端水平侧根直径的复相关系数,前者复相关系数达到0.736 9和0.739 2,后者复相关系数仅为0.502 7和0.511 1。
2.2.3 火炬树树高与水平侧根直径的关系火炬树树高与其前后端水平侧根直径相关性较弱。取样数据经过统计相关分析发现1 a幼苗和2 a幼树树高与其前端水平侧根直径复相关系数分别为0.628 9和0.676 9,而与其后端水平侧根直径复相关系数分别为0.564 0和0.515 2(表 1)。
2.2.4 火炬树克隆子株前后端水平侧根直径时间序列特点将火炬树克隆子株前后端水平侧根直径的调查数据按照年龄求出相应的平均值,然后根据克隆子株的年龄和前后端水平侧根直径的关系绘制直方图。由(图 2A)可知,生长分布于荒山生境的火炬树,随年龄增加克隆子株后端水平侧根直径增加幅度较小,根径由1.01 cm增大至1.94 cm,增加了0.93 cm, 相比较克隆子株前端水平侧根直径增加的幅度非常大,根径由1.71 cm增大至4.5 cm, 增加了2.79 cm。火炬树克隆子株前端水平侧根直径由1 a生到4 a生增大了2.63倍,后端水平侧根直径仅增大了1.92倍,结果出现了火炬树克隆子株1 a生幼苗前端水平侧根直径与4 a生幼树后端水平侧根直径相差很小的现象。由(图 2B)可知,通过克隆繁殖生长于石隙生境的火炬树,克隆子株的年龄分别属于1 a、2 a和3 a时,前端水平侧根直径分别为0.98、2.12和4.5 cm,增加了3.52 cm,与此同时后端水平侧根直径分别为0.43、1.05和1.8 cm,增加了1.37 cm。3 a的前端水平侧根直径是1 a的4. 59倍,而3 a后端水平侧根直径是1 a的4.19倍。可见随着树龄的增加,前端水平侧根直径增加倍数和幅度均大于后端水平侧根直径。

|
图 2 荒山(A)、石隙(B)火炬树克隆子株前后端水平侧根直径的年龄序列 Fig. 2 Age series of RDF and RDB of the R. typhina clonal ramets on barren hills(A) and in rocky crevice(B) LRD: lateral root diameter; □前端直径RDF; ■后端直径RDB. |
比较表 2还可知道,生长分布在荒山草坡的火炬树子株,年龄由1 a增至4 a,前后端水平侧根直径的差值由0.7 cm增加到2.56 cm, 前后端水平侧根直径比值从169%增加至232%;同样生长在石隙生境的火炬树克隆子株,年龄由1 a增至3 a,前后端水平侧根直径差值亦逐渐增加,由0.55 cm增加到2.7 cm, 前后端水平侧根直径比值从228%增至250%。
|
|
优势种或建群种的生物量是度量生态系统能量固定和物质生产储量以及营养积累的主要数量指标。对于广大的温带暖温带半湿润森林分布区,落叶木本植物的生物量在一定程度上成为下一个生长季继续生长和繁殖的能量、物质和营养的重要蓄库。因此研究火炬树母株生物量的分配特点和克隆子株生物量的时空变化格局可以说明火炬树的生态对策、对空间资源的利用和占据特点、对克隆繁殖的影响程度。
2.3.1 火炬树母株和子株生物量的分配格局调查地段火炬树生长分布的生境类型属于荒山草坡,干旱贫瘠是其主要胁迫因子。在干旱贫瘠的条件下,植物光合有机产物较大比例积累于地下器官根系(孙儒泳,1993; 宇万太等,2001;尚玉昌, 2001),结果根系的生长超过树冠生长,从而增强了对干旱贫瘠生境的适应性。从收获法测得的火炬树地上各器官和地下各级根系的鲜重和烘干重(表 3)可知,说明火炬树栽植母株和子株生物量的分配特点。火炬树母树地上鲜重为5 425 g, 地下鲜重仅为3 115 g, 地上与地下鲜重比约为1.74;地上干重为2 987 g, 地下干重仅为1 300 g, 地上与地下干重比约为2.29;叶片与根系鲜重比为0.36,干重比约为0.35。地上部分鲜重的顺序为树干>树皮>叶片>树枝,而干重的顺序是树干>树枝>树皮>叶片;地下各级根系无论是鲜重还是干重,均是粗根>中根>小根>细根,而且粗根的鲜重和干重占根系鲜重和干重的比例分别为0.54和0.55,中根的鲜重和干重约是小根鲜重和干重的2倍。栽植母株(表 4)地上与全株生物量的比值高于根系与全株生物量的比值;根系鲜重生物量约占地上鲜重生物量0.57,而干重仅占0.25,说明根系的水分含量高于地上部分;与叶片相比,根系生物量的鲜重和干重分别为叶片的2.75倍和2.88倍。对于火炬树克隆子株而言,荒山和石隙生境的生物量随年龄增加相差较小,而且地上和地下生物量的分配比例及其变化趋势随着树龄增加在两种生境上亦基本相同(表 4)。随年龄增加,荒山和石隙生境克隆子株的根冠比、根与全株重量比值增加而且鲜重干重比值大致相近,相反冠占全株重量的比值减小但鲜重干重比值相同。克隆子株的根叶干重比随着年龄增加而增大,但石隙生境克隆子株3 a的根叶鲜重比略有下降。作为贮藏、吸收和繁殖器官的根系,其干重和鲜重均超过光合器官叶片。无论是荒山或石隙生境,地上与全株的生物量比值高于根系与全株,只是随年龄增加其比值略有减小。本研究认为地上生物量积累量超过地下根系积累量意味着火炬树初期(1~6 a)生长迅速,通过快速高生长和树冠生长尽早获得竞争的优势地位,从而成功排挤同一层片当地灌木和草本植物种;其次地下根系生物量主要集中于粗根和中根,粗根和中根是克隆子株的主要营养贮藏库。
|
|

|
|
火炬树克隆子株前端水平侧根生物量的大小关系到根系向前扩展延伸的能力和为新的子株形成提供的营养充足与否,直接影响火炬树克隆繁殖能力的强弱。本文选择测定火炬树前后端单位长度水平侧根的生物量,期望揭示不同年龄阶段克隆子株前后端水平侧根的特点。由图 3可知,随着火炬树克隆子株年龄的增加,其地径和水平侧根直径呈增大的趋势;而子株前后端水平侧根一级根的长度变化较为复杂,表现为由1 a生到2 a生先增大再到3 a生时稍稍下降。所以克隆子株间各级水平侧根长度呈多样化,为了直观准确地反映火炬树子株前后端水平侧根的生长规律,本文分析了单位根长生物量的变化特点。由图 4可以看出,克隆子株前后端单位根长水平侧根的生物量随年龄增大而增加,不同的是前端水平侧根的生物量增加的幅度更大。后端水平侧根生物量由2.69 g增加至10.136 g, 前端水平侧根由9.42 g增加至66.847 g,前端水平侧根增加幅度大约是后端水平侧根的8.5倍。相同条件下克隆子株前端水平侧根生物量积累的速度远大于后端水平侧根,从而更有利于火炬树快速拓展占据新的生长位点和保证子株的成活与初期快速生长。

|
图 3 火炬树克隆子株地径水平侧根数量特征 Fig. 3 Base stem diameter and lateral root characteristics of clonal ramets in R.typhina |

|
图 4 火炬树克隆子株单位根长生物量比较 Fig. 4 Biomass of 10 cm length lateral root of clonal ramets in R.typhina |
分布于荒山草坡和石隙二种生境的火炬树,随着树龄增加生物量的积累量大致相近(图 5、6),2 a生长于荒山草坡的克隆子株全株鲜重和干重分别为1 230 g和544.344 g,同样为2 a生长于石隙生境的克隆子株全株鲜重和干重分别为1 153 g和513.669 g。

|
图 5 荒坡火炬树基株子株生物量比较 Fig. 5 Biomass comparison between R.typhina's ramet and its ramets on barren hills |

|
图 6 石隙火炬树子株生物量比较 Fig. 6 Biomass comparison of the clonal ramets of R.typhina growing in rocky crevice |
火炬树在我国引种历史不长,由于其水平侧根形态和生长发育的特殊性,可以认为火炬树具有极强的入侵潜力,入侵行为正在表达。从形态和生长发育的角度出发,幼期树冠生长迅速,既可同草本层和灌木层的当地种竞争而获得优势,形成单优群落,又可保证单株叶面积的快速增加,提高光合强度,从而为水平侧根生长提供大量的光合有机产物;火炬树克隆子株前后端水平侧根直径大小的不对称性应该视为光合有机产物积累不平衡的结果,火炬树克隆子株具有光合有机产物在水平侧根趋前贮藏的特征,为其快速克隆繁殖准备最小空间间隔的相对充足的营养库。已有研究指出,水平侧根可作为芽库进行克隆繁殖(Maun et al., 1984), 还可作为物质传输的路径。通过对火炬树的调查和研究认为,水平侧根亦可作为有机营养物质的贮藏库,为不定芽萌动、子株初期快速生长提供充足的营养物质。
火炬树克隆子株前后端水平侧根直径与地径的相关系数最大,冠幅、树高较小;其中克隆子株前端水平侧根直径与上述三者的复相关系数都高于后端水平侧根直径的复相关系数。说明克隆子株前端水平侧根的直径与基径和冠幅的生长具有较高程度的同步性。
荒山和石隙生境火炬树克隆子株单位长度水平侧根以及母株子株生物量的分析结果显示子株前端水平侧根高于子株后端水平侧根,全株生物量分配格局为植冠高于根系。因此火炬树生物量积累的空间格局主要表现为树冠高于根系,而根系生物量又以克隆子株水平侧根前端大于后端。显然火炬树生物量积累格局更有利于克隆繁殖和占据太行山低山丘陵灌草群落上层,而不是采取高比例生物量积累于根系的生态对策以适应干旱的低山阳坡生境。应该认为这是火炬树在干旱瘠薄生境和荆条酸枣灌丛的双重生态选择压力下所采取独特而有效的生态适应对策。
克隆植物克繁殖器官具有多样性的特点,如根状茎型、匍匐茎型、丛生型、球茎型、鳞茎型和根出条型(于飞海等,2001),其中根茎型克隆植物一般研究子株根茎的分枝数目、分枝角度和间隔子长度(宋明华等,2002),还应研究根茎长度(钟章成,2001)。对于火炬树, 子株前后端水平侧根直径是克隆器官重要的形态学参数。因此克隆植物繁殖对策的多样性是植物生长繁殖与生境干扰的时空异质性相互作用权衡的结果。
本文所进行的研究属于火炬树克隆繁殖基础的生态学问题,上述结论随着地域和时间尺度的扩大,必然会发生相应的变化。今后尚待深入研究的内容依然很多,比如火炬树克隆子株前后端水平侧根解剖结构、激素调节水平、基株与子株的生理整合特点和火炬树种群拓殖扩散的规律以及火炬树与侧柏(Platycladus orientalis)、酸枣、荆条的种间关系表现形式和调控途径。
董鸣. 1996. 资源异质性生境中的植物克隆生长:觅食行为. 植物学报, 38(10): 828-835. |
董鸣主编. 陆地生物群落调查观测与分析. 北京: 中国标准出版社, 1997: 59-72
|
段新玲, 任东岁, 赵树珍. 2000. 火炬树的组织培养及植株再生. 植物生理学通讯, 6: 535. |
李根前, 黄宝龙, 唐德瑞. 2001. 毛乌素沙地中国沙棘无性系种群林缘扩散规律. 南京林业大学学报, 25(2): 9-13. |
李吉跃, 何军. 1992. 火炬树苗木耐旱特性的研究. 北京林业大学学报, 14, supp. 2: 9-16. |
李振宇, 解炎. 2002. 中国外来入侵种. 北京: 中国林业出版社, 1-53.
|
马淑英, 胡正海. 1997. 火炬树分泌道的发育解剖学研究. 西北植物学报, 17(5): 112-117. DOI:10.3321/j.issn:1000-4025.1997.05.022 |
毛学文, 施文甫, 王弋博. 1997. 火炬树腺毛的形态结构和发育的研究. 西北植物学报, 17(6): 137-139. DOI:10.3321/j.issn:1000-4025.1997.06.028 |
潘志刚, 游应天主编. 中国主要外来树种引种栽培. 北京: 中国林业出版社, 1994: 525-529
|
庞福生, 刘孝智, 李晓伟, 等. 2001. 侧柏火炬树混交林蓄水保水效益的研究. 水土保持研究, 8(3): 14-15. DOI:10.3969/j.issn.1005-3409.2001.03.005 |
尚玉昌编著. 普通生态学(第二版). 北京: 北京大学出版社, 2001: 343-346
|
宋明华, 董鸣. 2002. 群落中克隆植物的重要性. 生态学报, 22(11): 1960-1967. DOI:10.3321/j.issn:1000-0933.2002.11.023 |
孙立元, 任宪威主编. 河北树木志. 北京: 中国林业出版社, 1997: 350-352
|
孙儒泳, 李博, 诸葛阳, 等. 1993. 普通生态学. 北京: 高等教育出版社, 211-229.
|
解炎. 2002. 恢复中国的天然植被. 北京: 中国林业出版社, 7-18.
|
徐承远, 张文驹, 卢宝荣, 等. 2001. 生物入侵机制研究进展. 生物多样性, 9(4): 430-438. DOI:10.3321/j.issn:1005-0094.2001.04.016 |
徐化成, 郑均宝主编. 封山育林研究. 北京: 中国林业出版社, 1994: 90-103
|
于飞海, 董鸣. 2001. 生态学研究热点——克隆植物生态学. 植物, 3: 32-33. |
宇万太, 于永强. 2001. 植物地下生物量研究进展. 应用生态学报, 12(6): 927-932. DOI:10.3321/j.issn:1001-9332.2001.06.030 |
钟章成. 2000. 植物种群生态适应机理研究. 北京: 科学出版社, 1-9.
|
Donald J Leopold, William C Mccomb, Robert N. 1998. Muller Trees of central hardwood forests of North America. Timber Press, Portland, Oregon: 375-377. |
Gelderloos O G. 1988. Pollination of staghorn sumac by ants. American nurseryman, 168(12): 65-67. |
James W Hardin, Donald J Leopold, Fred M White. Textbook of Dendrology (Ninth edtion) McGraw-Hill Higner Education, 2001: 460-461
|
Maun M A, Lapierre J. 1984. The effects of burial by sand on Ammophila breviligulata. Journal of Ecology, 72: 827-839. DOI:10.2307/2259534 |
Werner P A, Harbeck A L. 1982. The pattern of tree seedling establishment relative to staghorn sumac cover in Minchigan old fields. American Midland Naturalist, 108(1): 124-132. DOI:10.2307/2425301 |