 2004, Vol. 40
2004, Vol. 40文章信息
- 陶建平, 臧润国.
- Tao Jianping, Zang Runguo.
- 海南霸王岭热带山地雨林林隙幼苗库动态规律研究
- Studies on the Dynamics of Seedling Bank in Gap of Tropical Montane Rain Forest in Bawangling, Hainan Island
- 林业科学, 2004, 40(3): 33-38.
- Scientia Silvae Sinicae, 2004, 40(3): 33-38.
-
文章历史
- 收稿日期:2002-09-16
-
作者相关文章


森林天然更新是森林生态系统自我繁衍恢复的手段,林隙更新是森林天然更新的一种重要方式。林隙面积大小、林隙形成的季节(Brokaw, 1985;Denslow,1987;Canham, 1990)以及林隙年龄长短(Whitmore, 1989;Runkle, 1989)等都对种子散播和林隙小气候有强烈影响,从而直接影响着更新状况及其与周围林分结构的差异。传播到一定生境的种子萌发成为幼苗后,常常形成幼苗库,作为缓冲机制,幼苗库在幼苗补充为幼树或大树的过程中具有重要的“保险”作用。林隙内种子库和幼苗库是森林植物种群动态和森林更新的一个重要环节,是森林生物多样性维持机制的一个重要方面。林隙更新过程中,林隙内幼苗的种类和数量随着林隙年龄和林隙面积的动态改变而发生着变化,最后只有少数种类的小部分个体进入林冠层,完成林隙树种的更新(臧润国等, 2001; Poulson et al., 1989),因此,研究林隙更新动态对揭示林隙树种的更替规律和森林循环的动态规律都有重要的科学意义(Brokaw, 1985; Kneeshaw et al., 1998)。
从20世纪70年代至今,林隙已日益成为生态学研究的热点,在我国红松(Pinus koraiensis)阔叶林和亚热带常绿阔叶林中,关于林隙更新的研究已较为充分(李旭光等, 1997;臧润国等, 1998;1999a;王周平等, 2000;2001)。海南岛热带山地雨林结构复杂,种类繁多,物种多样性丰富,这导致了对其研究的难度,有关林隙树种更新动态规律的研究,特别是涉及林隙树种幼苗动态的研究,在此类型森林中还鲜有人涉足(臧润国等, 1999b)。本文拟通过对不同大小和不同年龄林隙内树种的组成和数量特征进行分析,探讨不同特征林隙内树种更替的数量变化规律,为热带森林的更新、保护和持续经营提供参考。
1 研究地点与研究方法 1.1 研究地区概况调查地点设在海南岛霸王岭国家级自然保护区保护站旧址东北侧的热带山地雨林内,北纬18°50′~19°05′,东经109°05′~109°25′,属于原始潮湿热带常绿略带轻微季节性的热带山地雨林。坡度大部分在5~10°左右,山地砖红壤。热带季风气候,年平均温为23.6℃,年平均降水量为1 500~2 000 mm。主要是以陆均松(Dacrydium pierrei)和线枝蒲桃(Syzygium araiocladum)等为优势的热带山地雨林群落。
1.2 研究方法调查在中山大学于1983年所设的5块50 m×50 m的热带山地雨林永久样地内进行。在固定样地内随机选取林隙27个,对每个林隙都辩认林隙形成木的种类,测量其胸径和高度,根据在当地林区工作多年的专业人员的经验并结合保护区周围采伐后树木的腐烂情况,估测每个倒木出现的年龄(臧润国等, 1999a), 结合固定样地的调查记录(每5 a一次), 推测林隙的发育阶段。依据林隙的两种定义(Runkle, 1982; 臧润国等,1999a)对每个林隙的大小进行精确测定,即将林隙周围胸径在18 cm以上的树木作为扩展林隙(EG)的边界,精确测定EG周围树木的胸径和树高,并对每棵树木做树冠投影图,用罗盘仪导线测量法,测定不同的周围树木之间的距离以及不同树木连线之间的夹角,最后绘制出扩展林隙的多边形平面图,依据多边形平面图,就可以精确计算扩展林隙的面积。对扩展林隙内所有的乔灌木都进行种名辨认、测量高度,并对高度在1.5 m以上的树木测定胸径。此处,规定胸径≤1 cm的树木为幼苗,而1~10 cm为幼树,>10 cm者为大树。
2 结果与分析 2.1 林隙内树木的种类组成和数量在霸王岭热带山地雨林内共调查林隙27个,面积6 831 m2。在林隙中,共有乔木树种116种,其中黄叶树(Xanthophyllum hainanense)、乌心樟(Cinnamomum tsangii)、线枝蒲桃(Syzygium araiocladum)、白背槭(Acer decandrum)、厚壳桂(Cryptocarya chinensis)、侯氏新木姜(Nephelium topengii)、龟背灰木(Symplocos andenophylla)、厚皮香八角(Illicium ternstroemioides)、油丹(Alseodaphne hainanensis)和香楠(Cinnamomum rigidissimum)等的个体数量较多,它们仅占总种数的8.47%,但却占到个体总数的47.03%;其它一些分布不均匀的树种,比如山杜英(Elaeocarpus sylvestris)、土密树(Bridelia blansae)、番椒槁(Listsea baviensis)、海南肖榄(Platea hainanensis)、海南油杉(Keteleeria hainanensis)和大叶新木姜(Neolitsea levinei)等的个体数量很少,58个种(占总种数的50%)的个体总数仅占所有个体总数的6.02%。可见,热带山地雨林林隙中树种的种类组成相当丰富,但个体数量分布却极不均匀,种类间数量差别很大。由于林隙形成时间长短不同,树种的分布和数量也存在较大差异。在林隙形成初期,林隙中光照条件优越,物种间竞争较小,此时林隙中一些树种比如油丹(Alseodaphne hainanensis)、竹叶松(Podocarpus annamiensis)、黄叶树、白背槭、白椎(Castanopsis kawakamii)等树种的个体数量,特别是幼苗(幼树)的个体数量较多,密度较大。随着林隙年龄的增加,林隙内光照条件的改变,竞争加强,这些树种的个体数量迅速减少。相反,一些耐荫树种比如厚壳桂、线枝蒲桃、灰木(Symplocos spp.)、乌心樟、尖峰润楠(Machilus monticola)等则在年龄较大的林隙中有较高的个体数量。
2.2 林隙特征对林隙幼苗和幼树种类和数量的影响 2.2.1 林隙面积对林隙内树种数量的影响对13个年龄在20~30 a的林隙内个体数量特征的分析表明:单位面积树木幼苗种数(种密度)和单位面积树木幼苗个体数量(个体密度)随林隙面积增大而迅速减小,但幼树和成树的个体密度和种密度的值随林隙面积则变化不大。整个林隙内树木的个体密度和种密度的变化主要决定于幼苗数量和种类的变化,而幼树和成树的种类和数量变化对总体树木的密度影响不大(图 1a,b)。幼苗和幼树每个体拥有的种数(species per stem)在不同面积林隙间波动不大,成年树木每个体拥有的种数随林隙面积的增大而减小,其值远高于幼苗和幼树每个体拥有的种数(图 1c)。随着林隙面积的增加,树种的种密度虽然逐渐降低,但这只是一种相对数量的减小,并不意味着大林隙内树种数量少于小林隙,相反,林隙面积越大,林隙内的树种越多。统计表明,当林隙面积达到300 m2时,其中的树木种类达到75种,并有继续上升的趋势。

|
图 1 不同面积林隙对林隙树种数量的影响 Fig. 1 Effects of different gap size on the number of gap fillers —◇—幼苗Seedlings;—□—幼树Saplings;—△—成树Trees;—x—总体Total.图例下同。The cutlines are same below. |
对12个面积在100~200 m2的林隙内个体数量特征的分析表明:树木不同发育阶段的个体密度、种密度以及每个体拥有的种数随林隙年龄变化有不同表现,幼苗的个体密度在小于40 a的林隙中变化不大,然后在40 a以上的成熟林隙中略有上升。幼树和大树的个体密度在不同年龄林隙中变化不大(图 2a)。幼树和幼苗的种密度在中等年龄林隙(20~40 a)中较小,而在幼年林隙(<20 a)和成熟林隙(>40 a)中都较大,种密度随林隙年龄有先下降而后上升的过程(图 2b)。林隙内树木总体的个体密度和种密度的变化主要决定于幼苗的数量和种类(图 2a,b)。早期林隙较大的种密度可能是由于这些林隙内存在有较多先锋树种幼苗的缘故,而成熟林隙内种密度和个体密度的升高则可能是由于顶级耐荫树种大量发生的缘故。不同年龄林隙内幼苗每个体拥有的种数变化不大,幼树和成年树木每个体拥有的种数则随林隙年龄的增加先降低然后升高(图 2c)。

|
图 2 不同林隙年龄对林隙树种数量的影响 Fig. 2 Effects of different gap age on the number of gap fillers |
将所有树木个体作为一个整体,按高度和径级分组,分析林隙树种的更新动态(图 3)。在小林隙和幼年林隙中, 高度小于100 cm或胸径小于1 cm的树木幼苗的密度要明显高于大林隙或成熟林隙,这表明,林隙形成的干扰环境有利于种子的萌发和幼苗的发生,这可能与林隙形成初期的优越光环境和林隙内较小竞争的小环境条件有关。大林隙和成熟林隙中的幼苗向幼树的转化率要明显高于小林隙和幼年林隙(表 1),在树高100 cm的个体向150 cm的个体的转化过程中,小于10 a的林隙中的转化率为0.51,大于40 a的林隙的转化率为0.63;面积小于50 m2的林隙的转化率为0.40,而面积大于250 m2的林隙的转化率为0.76。与其它径级相比,径级1~5 cm向5~10 cm的转化率最低,这表明,5~10 cm径级的幼树的死亡率较高或是胸径增长较快,这一阶段幼树的生存环境和更替状况可能对森林树种的数量特征和种类组成有重大影响,是树木更新的一个关键阶段。

|
图 3 林隙内不同大小级树木的数量变化 Fig. 3 Number of trees in different size classes in different gaps Ⅰ:H≤100 cm;Ⅱ:100 cm<H≤150 cm, D≤1 cm;Ⅲ:1 cm<D≤5 cm;Ⅳ:5 cm<D≤10 cm;Ⅴ:10 cm<D≤15 cm;Ⅵ:D>15 cm;H树(苗)高Height of trees;D胸径Diameter at breast hight.下同The same below. |
|
|
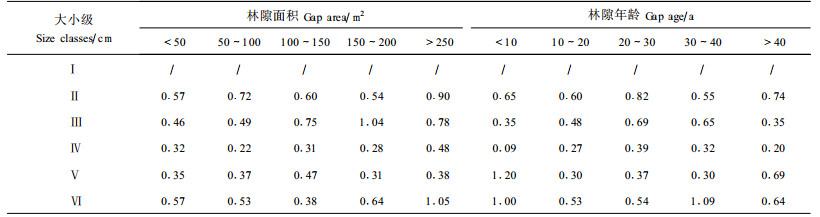
为了进一步了解树木个体更新的特点,本文按大小级对树木个体的更新动态进行了分析。从图 4可以看出,林隙树木更新过程中,个体数量差异主要出现在幼苗(幼树)阶段,当胸径大于10 cm时,个体数量趋于稳定。依据树木幼苗数量的变化形式,可以将林隙填充者分为以下类型:①急剧下降型,这一类型树种多属阳性树种,幼苗生长对光照要求苛刻,往往幼苗数量极多,但死亡率也很高,主要分布在幼年林隙或大林隙中。黄叶树、油丹、竹叶松、鸡毛松(Podocarpus imbricatus)、海南油杉等属于此类型,其中鸡毛松只存在高度在60 cm以下的幼苗,是此类中的极端类型。②“Z”型,这一类型树种也属于阳性树种,但相对耐荫,树木幼苗的数量在林隙郁闭到一定程度时才急剧下降。厚皮香八角、虎皮楠(Daphniphyllum oldhamii)、山茶(Caellia japonica)、鱼骨木(Canthium dicoccum)、侯氏新木姜等属于此类型。③均匀下降型,此类树种的幼苗在不同大小级的死亡率相差不大,属于喜阳耐荫的种类,它们能在较荫蔽的环境生长,对林隙的年龄和大小选择性不强。荔枝叶红豆(Qrmosia semicastrata)、白背槭、华润楠(Machilus chinensis)、多香木(Polyosma cambodiana)等属于此类型。④波动下降型,此类型中又分两类,一是波动中快速下降的喜阳耐荫树种,如乌心樟、龟背灰木、香楠、白椎、柏拉木(Blastus cochinchinensis)、尖峰润楠等,它们对林隙的选择性也不强;另一类是波动中缓慢下降的荫性树种,如线枝蒲桃、厚壳桂(Cryplocarya chinensis)、毛叶嘉赐树(Casearia membranacea)、灰木等,它们一般在成熟或郁闭度较大的林隙中生长,并且有较多的成年个体存在。

|
图 4 不同林隙树种数量动态的几种类型 Fig. 4 Quantitative dynamics of different gap fillers |
目前有关林隙干扰和林隙更新的研究已相当充分,但在热带森林林隙更新的研究中,一般只测定高度大于1.5 m的幼树和成树, 往往忽略幼苗的作用。由于幼苗的发生和成活状况将对森林群落的种类组成和演替方向产生重要的影响,甚至直接决定树木种群未来的命运(班勇等,1995),因而,林隙幼苗动态是森林群落动态和森林更新的一个重要环节。本研究表明,高度小于1.5 m的树木个体在热带林隙个体数量中占有较大的比重,它们在不同年龄和不同大小的林隙中有不同的种类组成、数量特征和行为表现,在林隙形成初期,由于有较优越的光照条件和较小的竞争环境,林隙中有较多的个体数量和较大的密度,此时,占据林隙生境的主要是一些喜光的先锋树种的幼苗,幼苗发生率高,死亡率也相对较高。在成熟林隙中,由于郁闭度增大,植株间资源竞争增强,林隙中的幼苗则主要是一些竞争能力强而相对耐荫的种类。林隙年龄和大小的差异不但影响林隙内个体的数量,也影响树种组成,不同大小和年龄林隙具有不同的林隙群落特征。
林隙面积直接影响林隙内光照状况(Canham, 1990),进而影响林隙内的温度和湿度(臧润国, 1999a),不同大小林隙具有不同的林隙小生境,因而影响其中树木幼苗的发生和生长。林隙面积对树木更新有较大影响,许多树种都需要特定大小的林隙来完成更新(Brokaw, 1987;Brokaw et al., 1989; Whitmore, 1989)。海南岛热带山地雨林林隙大小对林隙更新树种的种类组成和数量的影响主要表现在幼苗上,而成树和幼树在不同面积林隙中的种类和数量差异不大。林隙幼苗种密度和个体密度都随林隙面积增大而减小,这是由于幼苗个体的数量主要取决于单位林隙面积内繁殖体(种子或其他营养繁殖体)的数量和幼苗发生率的大小的缘故。树木种子成熟后主要散布于母树周围较近的地面上(胡玉佳等, 1992),因此,较大的林隙由于单位面积上的种子数量相对较少,即使优越的光环境和较小的竞争有利于幼苗发生,但面积较大的林隙内的幼苗密度和数量仍然相对较小。之所以说这种密度的减小是一种相对数量的减少,一方面是由于林隙内种类的绝对数目随林隙面积的增大而显著增加,另一方面则是由于随着林隙面积的增加,林隙树种幼苗每个体拥有的种数并无明显变化,这说明,个体密度的减小是由于种类相对减少造成的。
林隙年龄也影响着林隙更新状况及其与周围林分结构的差异,决定着林隙在森林循环中的地位(Runkle, 1989;Whitmore, 1989)。林隙种密度和个体密度对林隙年龄的反应也各不相同(图 2)。早期林隙中较大的幼苗种密度和个体密度主要是由于林隙初形成时具有较好的光照条件和较小的竞争,因而大量阳性树种幼苗和一些喜阳耐荫树种幼苗大量发生的缘故,随着林隙年龄的增加,郁闭度和竞争都相应增强,幼苗的发生和成活都随之减少。在成熟林隙内幼苗密度有所增加则可能是由于阴性树种幼苗的种类和数量增加的缘故。成树和幼树的竞争能力较强,受光照限制相对较小,因而在不同年龄林隙中的密度变化不大。
林隙形成了具有边缘效应的过渡地带,这种地带内存在有多种生态条件(刘济明, 2000),为具有不同竞争能力的树种提供了异质性的环境资源(Schnitzer et al., 2001), 这种异质环境有利于森林树种幼苗的定居(establishment)和个体密度的增加。有研究表明,林隙个体密度的增加导致了林隙物种多样性的增加,但这种多样性的增加会随着个体快速稀疏(死亡)而很快消失(Denslow, 1995;Stevens et al., 1999)。这种现象可能因为树种的生物学特性的差异而在不同大小和不同面积的林隙中有所不同,阳性先锋树种的幼苗在林隙中大量发生后个体数量会很快下降,而一些耐荫树种幼苗的发生速度和消亡速度都要缓慢得多,因而,具不同生物学特性的树种的物种多样性变化在不同林隙中可能表现出不同的形式。Schnitzer和Carson(2001)的研究就表明,对耐荫树种幼苗的物种多样性而言,林下和林隙内无显著差异,而先锋树种和藤本幼苗在林隙内的多样性则显著高于林下的多样性。本研究的结果还表明,林隙内树木的个体密度、种密度以及每个体拥有的种数在树木的不同发育阶段差异较大,因而对林隙物种多样性的变化必须要因测定对象范围的差异而作出不同的解释,只有这样才能够真实反映不同林隙内的物种多样性状况。
热带森林的种类组成非常丰富,霸王岭热带山地雨林林隙中的乔木树种达116种,但分布不均匀,密度差异较大。有研究表明,在霸王岭热带山地雨林中,达到主林层高度的树种只有30余种,密度也只有183株·hm-2,胸径达到50 cm以上的树种只有18种,仅占群落树种总数的13.04%。林隙更新后能进入主林层的树种也只是少数种类的少数个体(臧润国等, 2001),这些少数种类的少数个体是群落内的优势种,是整个热带山地雨林生态系统的主体。目前,对这些优势种的研究还缺乏动态资料,因此,有必要就林隙特征对这些优势树种更新的影响,特别是林隙特征对这些树种的种子萌发、幼苗发生以及幼苗消亡过程的影响作连续动态的研究,这对于热带山地雨林生态系统功能的维持以及生物多样性的保护都有重要意义。
班勇, 徐化成. 1995. 大兴安岭北部原始老龄林内兴安落叶松幼苗种群的生命统计研究. 应用生态学报, 6(2): 113-118. DOI:10.3321/j.issn:1001-9332.1995.02.006 |
胡玉佳, 李玉杏. 1992. 海南岛热带雨林. 广州: 广东高等教育出版社, 72-81.
|
李旭光, 何维明, 董鸣. 1997. 缙云山大头茶种群林窗动态的初步研究. 生态学报, 17(5): 543-548. DOI:10.3321/j.issn:1000-0933.1997.05.015 |
刘济明. 2000. 茂兰喀斯特森林主要树种的繁殖更新对策. 林业科学, 36(5): 114-122. DOI:10.3321/j.issn:1001-7488.2000.05.019 |
王周平, 李旭光, 石胜友, 等. 2000. 缙云山森林林隙形成特征的研究. 西南师范大学学报(自然科学版), 25(3): 306-309. |
王周平, 李旭光, 石胜友, 等. 2001. 重庆缙云山针阔混交林林隙树木更替规律研究. 植物生态学报, 25(4): 399-404. DOI:10.3321/j.issn:1005-264X.2001.04.003 |
臧润国, 郭忠凌, 高文韬. 1998. 长白山自然保护区阔叶红松林林隙更新的研究. 应用生态学报, 9(4): 349-353. DOI:10.3321/j.issn:1001-9332.1998.04.004 |
臧润国, 刘静艳, 董大方. 1999a. 林隙动态与森林生物多样性. 北京: 中国林业出版社, 3-13.
|
臧润国, 余世孝, 刘静艳, 等. 1999b. 海南霸王岭热带山地雨林林隙更新规律的研究. 生态学报, 19(2): 151-158. |
臧润国, 杨彦承, 蒋有绪. 2001. 海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究. 植物生态学报, 25(3): 270-275. DOI:10.3321/j.issn:1005-264X.2001.03.003 |
Brokaw N V L. 1985. Gap-phase regeneration in a tropical forest. Ecology, 66: 682-687. DOI:10.2307/1940529 |
Brokaw N V L. 1987. Gap-phase regeneration of three pioneer tree species in a tropical forest. Journal of Ecology, 75: 9-20. DOI:10.2307/2260533 |
Brokaw N V L, Scheiner S M. 1989. Species composition in gaps and structure of a tropical forest. Ecology, 70: 538-541. DOI:10.2307/1940196 |
Canham C D. 1990. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forest. Journal of Forest Research, 20: 620-631. |
Denslow J S. 1987. Tropical rain forest gaps and tree species diversity. Annual Review of Ecology Systematics, 18: 431-451. DOI:10.1146/annurev.es.18.110187.002243 |
Denslow J S. 1995. Disturbance and diversity in tropical rain forest: the density. Ecological Applications, 5: 962-968. DOI:10.2307/2269347 |
Kneeshaw D D, Bergeron Y. 1998. Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology, 79: 783-794. DOI:10.1890/0012-9658(1998)079[0783:CGCATR]2.0.CO;2 |
Poulson T L, Platt W J. 1989. Gap light reg?mes influence canopy tree diversity. Ecology, 70: 553-555. DOI:10.2307/1940202 |
Runkle J R. 1982. Patterns of disturbance in some old-growth mesic forests of eastern North America. Ecology, 63: 1533-1146. DOI:10.2307/1938878 |
Runkle J R. 1989. Synchrony of regeneration gaps, and latitudinal differences in tree species diversity. Ecology, 70: 546-547. DOI:10.2307/1940199 |
Schnitzer S A, Carson W P. 2001. Treefall gaps and the maintenance of species diversity in a tropical forest. Ecology, 82: 913-919. DOI:10.1890/0012-9658(2001)082[0913:TGATMO]2.0.CO;2 |
Stevens S M, Carson W P. 1999. Plant density determines species richness along an experimental fertility gradient. Ecology, 80: 455-465. DOI:10.1890/0012-9658(1999)080[0455:PDDSRA]2.0.CO;2 |
Whitmore T C. 1989. Changes over twenty-one years in the Kolombangara rain forest. Journal of Ecology, 77: 469-483. DOI:10.2307/2260763 |