 2004, Vol. 40
2004, Vol. 40文章信息
- 陈立新, 杨承栋.
- Chen Lixin, Yang Chengdong.
- 落叶松人工林土壤磷形态、磷酸酶活性演变与林木生长关系的研究
- The Succession of Various Types of Phosphorus, Phosphatase Activity, and the Relationship with the Tree Growth in Larch Plantations
- 林业科学, 2004, 40(3): 12-18.
- Scientia Silvae Sinicae, 2004, 40(3): 12-18.
-
文章历史
- 收稿日期:2003-02-12
-
作者相关文章


2. 中国林业科学研究院林业研究所 北京 100091
2. Research Institute of Forestry, CAF Beijing 100091
长白落叶松(Larix olgensis)素有“北方杉木”的美称,是我国东北地区最主要的造林树种,也是北方针叶树种中最为速生丰产的树种之一。然而,众多的研究结果表明:落叶松人工林林地土壤质量存在着不同程度的下降趋势(浅田廃夫,1981;王秀石,1982;张慧亮等,1987;刘世荣等,1993;阎德仁等,1997;潘建平等,1997;陈立新等,1998a),尤其是进入近熟林以后,林地缺磷现象明显,成为林木生长的限制因子(高雅贤,1983;陈立新等,1998b),这一问题已引起了学术界的普遍关注。为了揭示落叶松人工林土壤质量的演变机理,寻找影响林木树高、胸径生长量的主要磷形态,本文对处于相似立地条件下不同发育阶段的落叶松人工林根际与非根际土壤磷形态、土壤磷酸酶活性的演变规律进行了研究,揭示了影响林木生长的主要土壤磷形态。
1 材料与方法 1.1 试验地概况试验地位于吉林省长春市九台土们岭铁路林场,北纬44°06′,东经126°02′。本区是长白山向西部松辽平原过渡的丘陵地带,最高山峰马虎头山海拔为542m。属大陆性季风气候,最高气温为42℃,最低气温为-36℃,年平均气温为5℃,年降水量200~849mm,无霜期约为130~140d。原始植被为红松(Pinus koraiensis)阔叶混交林,经过人为活动形成了天然蒙古栎(Quercusmongolica)次生林和各种人工林植被。地带性土壤为暗棕壤,微酸性。
1.2 样地设置和土壤样品的采集在相似立地条件的落叶松人工幼龄林、中龄林、近熟林、成熟林各选择3块固定标准地,对标准地内的林木进行每木检尺,每块标准地选择一株优势木作为解析木,求得优势木胸径、树高定期平均生长量和连年生长量。同时在每块标准地内选择标准木3株,并在每一株标准木的周围按不同方位设置5个采样点,挖出表层细根,采用抖落法收集根际土壤混合样品。在各标准地内,沿S形曲线进行布点,采集0~20、20~40、40~60cm的非根际土壤,带回室内进行分析。
1.3 土壤样品的分析土壤全磷用硫酸-高氯酸酸溶, 钼锑抗比色法测定。土壤无机P用张守敬和Jackson提出的分级方法测定(中国土壤学会农业化学专业委员会,1983);有机P用灼烧-0.2mmol·L-1H2SO4浸提法测定;有效P用0.03mmol·L-1NH4F-0.025mmol·L-1HCI浸提,然后用钼锑抗比色法测定(中国土壤学会农业化学专业委员会,1983);酸性、中性磷酸酶采用Hoffmann法测定(关松荫等,1986;郑洪元,1982)。
2 结果与分析 2.1 不同发育阶段落叶松人工林土壤磷形态有效性的演变 2.1.1 土壤全磷含量的演变磷是植物生长的主要营养元素之一,土壤中95%的磷是以迟效性状态存在,而且不同磷形态其有效性不同。全磷含量高时并不意味着磷素供应充足,而全磷含量低于0.8~1.0g·kg-1时,土壤常出现磷供应不足。有机磷只有少部分是有效的,绝大部分有机磷必须在土壤微生物和磷酸酶作用下,将磷酸肌醇、磷脂等含磷有机化合物转化为植物可利用的无机磷酸盐(Molla et al., 1984),释放出生物有效磷。研究结果表明,各年龄阶段土壤全磷含量根际土壤大于非根际土壤。从幼龄林到中龄林,根际土壤全磷含量增高,并达到最大值,到近熟林全磷含量降低到最小,近熟林到成熟林略有提高,但成熟林与幼龄林相比土壤全磷含量降低14.67%;从幼龄林到中龄林非根际土壤全磷含量增高,中龄林到近熟林全磷含量降低,近熟林到成熟林全磷含量提高,非根际土壤全磷含量最高值出现在成熟林。对不同发育阶段落叶松人工林的根际与非根际土壤全磷含量进行t检验,结果显示,幼龄林、中龄林、近熟林根际与非根际土壤间土壤全磷含量差异均达到极显著和显著水平[幼龄林t=5.28>t0.01(1,16)=2.92;中龄林t=3.33>t0.01(1,16)=2.92;近熟林t=2.91>t0.05(1,16)=2.12]。成熟林根际与非根际土壤全磷含量差异不显著[t=1.50<t0.05(1,16)=2.12]。落叶松人工林各年龄阶段间根际、非根际土壤全磷含量差异均达到显著水平(表 1)。
|
|
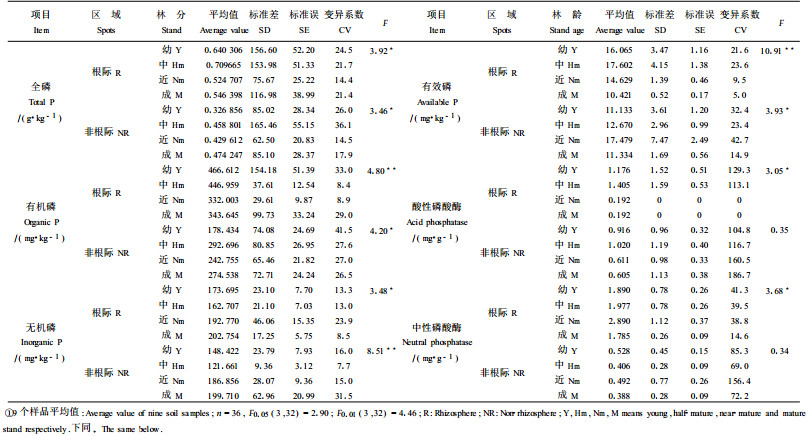
落叶松人工林不同发育阶段土壤有机磷含量与全磷含量变化规律并不完全一致(表 1)。从幼龄林到近熟林,根际土壤有机磷含量持续降低,到了成熟林,由于抚育间伐,林地光照条件的改善,林下植被生长旺盛,有机磷含量有所提高。各年龄阶段土壤有机磷含量根际大于非根际。差异性检验结果显示,幼龄林、中龄林、近熟林根际土壤与非根际土壤间土壤有机磷含量差异均达到极显著水平[幼龄林t=5.05>t0.01(1,16)=2.92;中龄林t=5.19>t0.01 (1,16)=2.92;近熟林t=3.37>t0.01(1,16)=2.92]。成熟林根际与非根际土壤有机磷含量差异不显著[t=1.68<t0.05(1,16)=2.12]。落叶松人工林各年龄阶段间根际、非根际土壤有机磷含量差异达到极显著、显著水平。
2.1.3 土壤无机磷形态的演变落叶松人工林不同发育阶段根际和非根际土壤无机磷总量的变化规律与根际土壤全磷含量的变化规律相反(表 1)。从幼龄林到中龄林降低,中龄林以后持续增高;成熟林时,无机磷含量超过了幼龄林的16.7%。表明中龄林以前以有机质积累为主,有机磷的矿化作用较弱,中龄林以后根系分泌有机酸增多和微生物活性增强,对磷的活化作用也随之增强,根际微域的有机磷被矿化为无机磷,因此表现出幼龄林到中龄林土壤全磷含量和有机磷含量增加,无机磷含量减少,中龄林以后根际土全磷含量和有机磷含量减少,无机磷含量增加。各年龄阶段土壤无机磷含量根际大于非根际,检验结果显示,幼龄林和中龄林根际与非根际土壤无机磷含量差异达到显著和极显著水平[幼龄林t=2.29>t0.05(1,16)=2.12;中龄林t=5.33>t0.01(1,16)=2.92]。近熟林和成熟林根际与非根际土壤无机磷含量差异不显著[近熟林t=0.33<t0.05(1,16)=2.12;成熟林t=0.14<t0.05(1,16)=2.12]。
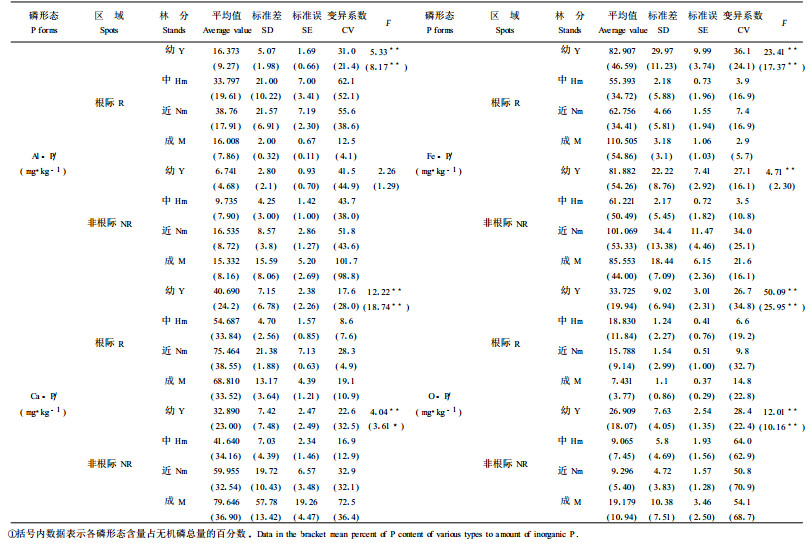
无机磷分组测定结果表明, 不同发育阶段以Fe-P(铁结合态的磷酸盐)占无机磷总量的百分比最高,达34.41%~54.86%,Ca-P(钙结合态的磷酸盐)为其次,占23.00%~38.55%,O-P(闭蓄态磷酸盐)为再其次,占3.77%~19.94%,Al-P最少,仅占4.68%~19.61%。无机磷各组分变化规律为Al-P(铝结合态的磷酸盐)含量从幼龄林到近熟林出现增加趋势,近熟林以后降低。Fe-P含量从幼龄林到中龄林降低,中龄林以后出现增加趋势。Ca-P含量随林龄的增加而增加,Ca-P含量的增加与土壤风化程度有关。O-P含量则相反。这是由于随林龄的增加O-P不断矿化释放出有效磷——Fe-P和Al-P,使土壤Al-P含量和Fe-P含量增加,最终使土壤无机磷总量增加。另外,在中龄林和近熟林时,Fe-P含量根际小于非根际,出现Fe-P亏缺现象,在成熟林时出现O-P和Ca-P的亏缺,说明落叶松不同发育阶段对无机磷形态的吸收是有选择性的,中龄林和近熟林林木以吸收Fe-P为主,成熟林林木以吸收Al-P和Ca-P为主。因此,表现出中龄林和近熟林出现Al-P含量和Ca-P含量增加,成熟林Fe-P含量增加,O-P含量、Al-P含量减少的变化规律。方差分析表明,各年龄阶段间根际土壤、非根际土壤Ca-P含量、Fe-P含量、O-P含量差异均达到极显著和显著水平,非根际土壤Al-P含量差异未达到显著水平(表 2)。
|
|
从幼龄林到中龄林,根际土壤有效磷含量有所增加,中龄林以后降低,到成熟林达到最低;从幼龄林开始到近熟林,非根际土有效磷含量一直处于增加的趋势,近熟林以后开始降低(见表 1)。近熟林和成熟林非根际土壤有效磷含量大于根际土壤,表明近熟林和成熟林在根际微域出现磷的亏缺。这是因为进入中龄林林木进入速生期,对养分需要量增大,磷将成为土壤微生物和植物生长的主要限制因子,因而,促进了微生物和植物根系分泌胞外磷酸酶,加快了有机磷的水解,释放出大量的生物有效磷,此时树木对磷的吸收量尚不是最大,中龄林以后,随着林龄增大,林木对磷的吸收量增多,到成熟林时林木对磷的吸收量达到最大。另外,近熟林和成熟林在根际微域出现有效磷的亏缺现象与Fe-P含量、Ca-P含量、O-P含量在近熟林和成熟林出现亏缺有关。所以,此阶段应注意磷的供应。差异性检验结果显示,近熟林和成熟林根际与非根际土壤有效磷亏缺现象并没有达到显著水平[近熟林︱t︱=1.13<t0.05(1,16)=2.12;成熟林︱t︱=1.55<t0.05(1,16)=2.12]。幼龄林、中龄林根际土壤有效磷浓度大于非根际土壤,差异显著[幼龄林t=2.96>t0.05(1,16)=2.12;中龄林t=2.90>t0.05(1,16)=2.12]。落叶松人工林根际土壤、非根际土壤有效磷含量各年龄阶段间差异达到极显著、显著水平。
2.2 土壤磷酸酶活性的演变不同发育阶段落叶松人工林根际与非根际土壤酸性磷酸酶活性的变化规律一致(表 1)。从幼龄林到中龄林,酸性磷酸酶活性增加,中龄林以后酸性磷酸酶活性降低,到成熟林达到最低。落叶松人工林幼龄林和中龄林根际土壤酸性磷酸酶活性大于非根际土壤,近熟林和成熟林土壤酸性磷酸酶活性非根际大于根际,表明近熟林和成熟林在根际微域出现酸性磷酸酶活性的下降,是造成近熟林和成熟林在根际微域出现有效磷亏缺的另一个主要原因。差异性检验结果显示,幼龄林、中龄林、近熟林和成熟林根际与非根际土壤酸性磷酸酶活性差异不显著[幼龄林t=0.43<t0.05(1,16)=2.12;中龄林t=0.58<t0.05(1,16)=2.12;近熟林︱t︱=1.27<t0.05(1,16)=2. 12;成熟林︱t︱=1.10<t0.05(1,16)=2.12]。经方差分析,落叶松人工林各年龄阶段之间根际土壤、非根际土壤酸性磷酸酶活性差异达到了显著或不显著水平。
不同发育阶段落叶松人工林根际与非根际土壤中性磷酸酶活性的变化为幼龄林到近熟林,根际土壤中性磷酸酶活性增加,近熟林以后中性磷酸酶活性降低,到成熟林达到最低;幼龄林到中龄林,非根际土壤中性磷酸酶活性降低,中龄林到近熟林中性磷酸酶活性提高,近熟林以后非根际土壤中性磷酸酶活性降低,并降低到最低值。各年龄阶段落叶松人工林根际土壤中性磷酸酶活性大于非根际,根际土壤分别是非根际土的3.58、4.87、5.87和4.6倍。这促进了根际土壤中有机磷化合物被树木的吸收利用(Hedley et al., 1982)。差异性检验结果显示,幼龄林、中龄林、近熟林和成熟林根际与非根际土壤中性磷酸酶活性差异极显著[幼龄林t=4.54>t0.01(1,16)=2.92;中龄林t=5.27>t0.01(1,16)=2.92;近熟林t=6.28>t0.01(1,16)=2.92;成熟林t=11.14>t0.01(1,16)=2.92]。经方差分析,落叶松人工林各年龄阶段之间根际、非根际土壤中性磷酸酶活性差异达到显著或不显著水平。
综上所述,不同发育阶段落叶松人工林根际土壤磷酸酶总活性与有效磷含量变化趋势一致。从幼龄林到中龄林,根际土壤磷酸酶总活性增加,中龄林以后降低,到成熟林达到最低;而在非根际土壤中并不存在这一规律,非根际土壤有效磷含量从幼龄林到中龄林升高,中龄林以后降低,到成熟林达到最低,而磷酸酶总活性从幼龄林开始一直处于降低趋势,到成熟林达到最低。
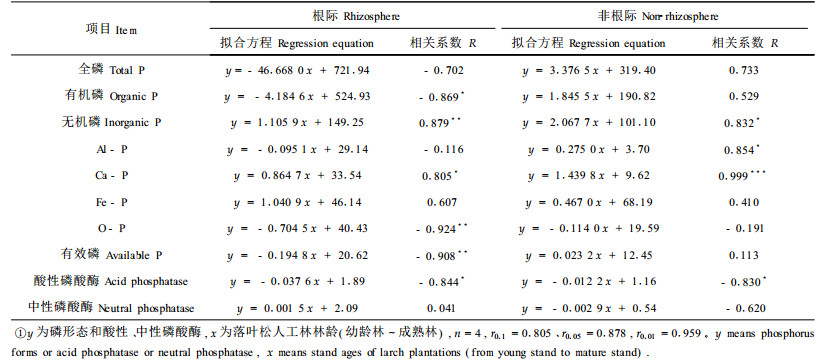
2.3 落叶松人工林土壤磷形态及磷酸酶活性与林龄的关系落叶松人工林不同发育阶段土壤磷形态及磷酸酶活性的演变规律,可以用线性方程进行拟合(表 3)。结果显示,根际土壤有效磷和O-P随林龄增加呈显著的减少趋势,根际土壤全磷含量、有机磷含量、酸性磷酸酶活性随林龄增加有明显的减少趋势。根际土壤无机磷随林龄的增加呈显著的增加趋势,其中Ca-P含量、Fe-P含量有明显的增加趋势。非根际土壤无机磷总量、Al-P含量和Ca-P含量随林龄的增加呈明显或极显著的增加趋势,酸性磷酸酶活性随林龄增加有明显的减少趋势。
|
|
应用SPSS统计软件的Stepwise逐步回归的方法,以林木树高、胸径平均、连年生长量作为因变量,土壤磷形态、有机质、速效钾、水解氮作为自变量,在落叶松不同发育阶段中,建立林木树高、胸径生长量对土壤磷形态、有机质、速效钾、水解氮的逐步回归模型,筛选出了对树种生长影响较大的土壤养分因子(表 4)。结果显示,在幼龄林影响林木胸径生长的主要因子是全磷、有效磷、无机磷总量和速效钾,影响林木树高生长的主要因子是水解氮、有机质、有效磷、有机磷和Ca-P,复相关系数均达到了显著或极显著水平;在中龄林影响林木胸径生长的主要因子是无机磷总量、Fe-P、全磷和速效钾,影响林木树高生长的主要因子是水解氮、有机质、Ca-P、有机磷和有效磷。但复相关系数仅树高连年生长量达到了显著水平,其它胸径平均生长量和连年生长量以及树高平均生长量复相关系数均没有达到显著水平。这可能是因为此阶段影响林木生长的因子中,除考虑的土壤养分因子外,还有其它因子。如土壤物理性质、土壤微生物和土壤酶的活性等;在近熟林影响林木胸径生长的主要因子是速效钾、有机磷、有效磷、无机磷总量、水解氮和Fe-P,影响林木树高生长的主要因子是有效磷含量、有机质、全磷、速效钾、水解氮和无机磷总量;在成熟林影响林木胸径生长的主要因子是速效钾、Fe-P、有机质和水解氮,影响林木树高生长的主要因子是速效钾、水解氮和无机磷总量。依据偏向关系数的大小及显著性检验,以上结果显示,不同发育阶段影响林木生长量的主要土壤因子是有机质、速效钾、水解氮以及磷形态的全磷、有机磷、有效磷、无机磷总量。林木生长量与Ca-P和Fe-P也相关,但未达到显著水平。
|
|

从表 4还可以看出,幼龄林胸径和树高平均生长量与土壤有效磷呈极显著或显著负相关,与全磷、有机磷呈极显著或显著正相关,表明树木胸径和树高的生长需要消耗大量的有效磷,而每年林木吸收的养分的一部分又以凋落物等有机物的形式进入土壤,因而呈现出土壤有效磷含量随着林木的生长而显著降低,而土壤全磷、有机磷则相反,在幼龄阶段呈现出积累的趋势;在中龄林时,胸径和树高生长量与土壤全磷、Ca-P和Fe-P呈一定的负相关,说明随着林木的生长,土壤有效磷的消耗,造成土壤有效磷含量的降低,当成为林木生长的限制因子时,根系将分泌大量的有机酸,根际土壤酸性、中性磷酸酶活性增强(表 1),促进磷的矿化,土壤无机磷总量增加,此阶段林木增强了对Ca-P和Fe-P的吸收;在近熟林时,胸径和树高生长量与土壤全磷、有机磷、Fe-P呈极显著或显著或不显著负相关关系,与无机磷总量、有效磷含量呈极显著或显著或不显著正相关关系,说明近熟林阶段林木生长以吸收土壤Fe-P为主,而且随着林木的生长,土壤全磷、有机磷不断矿化转化成无机磷和有效磷,因此,土壤全磷、有机磷、Fe-P随着林木生长量的增加而减少,土壤无机磷总量、有效磷含量则相反。在成熟林时,胸径和树高生长量与土壤无机磷总量呈负相关关系,与Fe-P含量呈正相关关系,说明此阶段林木生长主要吸收Fe-P以外的其它无机磷形态如Al-P、Ca-P。综上所述,落叶松不同发育阶段影响林木胸径、树高生长的主要土壤磷形态随林龄的变化而变化,但各年龄阶段影响林木生长的主要磷形态是全磷、有机磷、无机磷总量、有效磷、Ca-P和Fe-P,进一步说明落叶松主要吸收利用有效磷的形态是Al-P、Ca-P和Fe-P,这一规律与无机磷形态变化规律相吻合。
3 结论与讨论落叶松人工林在整个生长过程中,根际土壤有效磷含量与根际土壤磷酸酶总活性、酸性磷酸酶活性变化规律一致。从幼龄林到中龄林,根际有效磷含量、磷酸酶总活性和酸性磷酸酶活性增高,中龄林以后持续降低;从幼龄林到中龄林根际与非根际土壤全磷含量、非根际有机磷含量增高,中龄林到近熟林降低,近熟林以后又有所恢复。
对幼龄林和中龄林而言,酸性磷酸酶对根际土壤磷形态具有较强的活化作用。随着林龄增大,酸性磷酸酶对磷形态的活化作用减弱,即到了近熟林和成熟林时,对根际土壤磷形态活化作用不明显。而到了近熟林和成熟林,中性磷酸酶对根际土壤无机磷、Fe-P和O-P活化作用明显增强,中性磷酸酶对各年龄阶段落叶松人工林根际土壤O-P具有显著的活化作用。
落叶松人工林土壤磷形态及磷酸酶活性与林龄相关性显示,根际土壤有效磷含量、O-P含量、全磷含量、有机磷含量、酸性磷酸酶活性随林龄增加呈显著或明显的减少趋势,根际土壤无机磷总量、Ca-P含量、Fe-P含量随林龄的增加呈显著或明显的增加趋势。非根际土壤Ca-P含量随林龄的增加呈极显著的增加趋势。
尽管近熟林和成熟林阶段有机磷矿化作用增强,无机磷总量增加,但仍表现出非根际土壤有效磷大于根际土壤,在根际微域出现磷的亏缺现象,这与酸性磷酸酶活性、Fe-P、Ca-P、O-P含量在近熟林和成熟林出现亏缺、林木对有效磷的吸收能力增强有关。但经方差分析,有效磷亏缺现象并没有达到显著水平。
在落叶松人工林不同发育阶段,影响林木胸径、树高生长的主要土壤养分因子随林龄的变化而变化。不同发育阶段影响林木生长量的主要养分因子是土壤有机质、速效钾、水解氮以及土壤磷形态的全磷、有机磷、有效磷、无机磷总量,其次是Ca-P和Fe-P;土壤磷形态的这一规律与其含量的变化相吻合。在逐步回归方程之中,自变量水解氮、速效钾等的偏相关系数值显示明显负影响效应,这很可能与林木速生过程,对水解氮、速效钾等需求量大,引起土壤中这些养分迅速下降有关,结果导致偏相关系数值呈现明显的负影响效应。
陈立新, 陈祥伟, 史桂香. 1998a. 提高落叶松人工林林地质量的研究. 东北林业大学学报, 26(13): 6-11. |
陈立新, 陈祥伟, 段文标. 1998b. 落叶松人工林凋落物与土壤肥力变化的研究. 应用生态学报, 9(6): 581-586. |
程国玲, 唐立君, 郎福生. 2001. 水曲柳落叶松纯林与混交林根际土壤氮磷养分特点及变化. 东北林业大学学报, 29(1): 26-29. DOI:10.3969/j.issn.1000-5382.2001.01.007 |
高雅贤. 1983. 落叶松人工林土壤中水、肥动态的研究. 林业科技, (2): 9-13. |
关松荫等编著.土壤酶及其研究法.北京:农业出版社, 1986
|
姜培坤, 徐秋芳, 钱新标. 1995. 杉木檫树根际土壤磷素研究. 浙江林学院学报, 12(3): 242-246. |
刘世荣, 李春阳. 1993. 落叶松人工林养分循环过程与潜在地力衰退趋势的研究. 东北林业大学学报, 21(2): 19-24. |
潘建平, 王华章, 杨秀琴. 1997. 落叶松人工林地力衰退研究现状与进展. 东北林业大学学报, 25(2): 59-63. |
王秀石. 1982. 落叶松人工林土壤因子变化规律的研究. 吉林林业科技, (4): 1-11. |
阎德仁, 王晶莹, 杨茂仁. 1997. 落叶松人工林土壤衰退趋势. 生态学杂志, 16(2): 62-66. DOI:10.3321/j.issn:1000-4890.1997.02.014 |
张慧亮, 高雅贤, 向黄怀. 落叶松人工林下的土壤肥力状况. 见: 营林科技论文集. 北京: 中国林业出版社, 1987: 144-147
|
郑洪元. 1982. 土壤动态生物化学研究法. 北京: 科学出版社, 173-265.
|
张彦东, 白尚斌, 王政权. 2001. 落叶松根际土壤磷的有效性研究. 应用生态学报, 12(1): 31-34. DOI:10.3321/j.issn:1001-9332.2001.01.007 |
中国土壤学会农业化学专业委员会编.土壤农业化学常规分析方法.北京:科学出版社, 1983
|
浅田廃夫. 1981. カラマツ造林学. 东京: 农林出版株式会社.
|
Gerretsen F C. 1948. The influence of micro-organism on the phosphate intaken by the plant. Plant and soil, 1: 51-60. DOI:10.1007/BF02080606 |
Molla M A Z, Chowdhury A A. 1984. Microbial mineralization of organic phosphate in soil. Plant and soil, 78: 393-399. DOI:10.1007/BF02450372 |
Hedley M J, et al. 1982. Plant induced change in the rhizospher of rape(Brassica napus var. emerald) seedlings III: Change in L value, soil phosphate fraction and phosphatase activity. New phytologist, 91(1): 45-56. DOI:10.1111/j.1469-8137.1982.tb03291.x |