 2002, Vol. 38
2002, Vol. 38文章信息
- 田国忠, 张志善, 李志清, 郭加虎, 申艳普.
- Tian Guozhong, Zhang Zhishan, Li Zhiqing, Guo Jiahu, Shen Yanpu.
- 我国不同地区枣疯病发生动态和主导因子分析
- DYNAMIC OF JUJUBE WITCHES' BROOM DISEASE AND FACTORS OF GREAT INFLUENCE AT ECOLOGICALLY DIFFERENT REGIONS IN CHINA
- 林业科学, 2002, 38(2): 83-91.
- Scientia Silvae Sinicae, 2002, 38(2): 83-91.
-
文章历史
- 收稿日期:2000-12-19
-
作者相关文章


2. 山西省农科院园艺研究所 太原 030023;
3. 河南省濮阳市林业科学研究所 濮阳 457000;
4. 陕西省清涧县科技局 清涧 718300
2. Research Institute of Horticulture, Shanxi Academy of Agricultural Science Taiyuan 030023;
3. Puyang Research Institute of Forestry Puyang 457000;
4. Bureau of Science and Technology of Qingjian County Qingjian 718300
枣疯病是我国枣树(Zizyphus jujuba Mill.)上的毁灭性病害, 多数感病品种幼苗当年即枯死, 幼树1 ~ 2a死亡, 成年结果树枣果产量逐年下降, 病枝上开出变态花器, 不结果, 或结不成熟的畸形果, 完全失去食用价值, 病树会在3 ~ 5 a逐渐衰退死亡, 因而给我国众多枣产区的红枣生产构成了巨大威协, 并使农民蒙受了很大经济损失(田国忠, 1998)。多年来, 国内外已在枣疯病的发生和危害、传播方式、发病因子调查、抗生素治疗、PCR检测病原、枣疯病原系统分类研究, 以及枣树组培脱毒等方面开展了许多研究(中国科学院上海生物化学研究所, 1974;孙淑梅等, 1988;王焯等, 1983;王祈楷等, 1981;陈子文等, 1984;何放亭等, 1996;田砚亭等, 1993; Kim, 1965; La et al., 1984; La, 1987; Pandey et al., 1976; Tian et al., 2000)。然而, 进入20世纪80年代中后期, 随着我国农村生产方式、体制的改变和枣树栽培面积、范围的扩大, 枣疯病在许多枣产区的发生和为害呈现出不断加重的趋势。迄今, 由于我国以往的枣疯病调查研究多局限于某一特定地区, 因而对我国不同地域和不同生态环境条件下病害发生特点一直缺乏系统的比较研究, 特别是近期对我国枣疯病的发生动态、发病因子变动资料采集和分析更是空白, 从而严重阻碍了对此病害的流行预测和防治策略的制定。
有鉴于此, 作者自1997 -11至2001 -11分别对北京、河南、陕西、山西、安徽、浙江、山东、河北等省市的部分枣树分布区开展了枣疯病的普查和典型调查研究, 以期全面了解当前我国枣疯病的发生和为害状况, 分析我国不同地区枣疯病的发生历史、现状和特点, 以及病害的流行趋势, 为我国枣疯病的危害评估、流行预测预报、持续控制策略研究和为制定针对性强的防治措施奠定必要的基础。
1 研究方法 1.1 调查地点和方法重点调查地点包括北京、河南、陕西、山西、浙江、安徽4省1市。其中北京主要调查市区居民院内散栽的枣树, 树龄10a至几百年不等, 品种杂; 市郊调查地位于娘娘府的一块20a左右树龄的枣林, 及海淀区周家巷村从山西交城引种的梨枣和骏枣品种枣园。河南濮阳市调查地点在市郊的太行村、杜家庄等地枣园, 主栽品种为扁核酸, 枣树周年多与小麦、花生等间作。陕西省设清涧县和佳县两个调查地区, 枣树多分布于黄土高原丘陵沟壑区, 佳县调查地点包括打火店苗圃、泥河沟乡和县城郊的黄河滩地、地处黄土高坡的小叶峰村和牛圈沟村, 木枣和油枣为主栽品种; 清涧县调查地为1998年在秀延河边平地新建的2个苗圃和沿无定河流域栽植的枣树, 重点对高杰村镇的李家崖、野桥畔、王家洼村和东拉河村开展了定点调查, 其主栽品种为方木枣和长木枣。山西省调查地点为临县枣分布区。安徽省调查地点为黄山市歙县杞梓里乡和宣州市水东镇枣区。浙江省调查地包括义乌市郊和东阳市水东枣园。
调查内容涉及枣树栽培历史和方式、枣疯病发生历史考察、现场普查、定点调查、随机抽查等。定点调查时间为每年6月份和10月份共两次。具体项目包括发病株率、症状严重度、树龄、品种、育苗和田间栽培管理方式、周围生态环境等。普查地区则涉及北京、河南、陕西、山西、安徽、山东、河北、浙江、江西等红枣主产区或零星分布区, 特别是枣疯病发生和为害区。
1.2 病菌检测方法用DAPI荧光显微镜, 检查枣树不同部位韧皮部的植原体特异性荧光(金开璇等, 1989)。先将枣树幼茎或叶柄鲜材料徒手切片, 迅速放入5 %戊二醛溶液中固定, 经DAPI(4′, 6-二脒基-2-苯基吲哚盐酸)染色后, 用Olympus落射荧光显微镜检查韧皮部植原体—DNA特异荧光, 所用紫外线激发滤光片波长为365nm, 阻断滤光片波长为420nm。植物自发荧光观察切片未用DAPI染色。
以植原体16S rRNA基因保守序列设计的两对引物R16mF2/R2和R16F2/R2进行直接PCR和Nested-PCR(田国忠等, 1999)。引物序列为R16mF2 : 5′ -CAT GCA AGT CGA ACG GA-3′, R16mR2 : 5′ -CTT AAC CCC AAT CAT CGA-3′; R16F2 : 5′ -ACG ACT GCT AAG ACT GG-3′, RI: 5′ -TGA CGG GCG GTG ACA AAC CCC G-3′。
进行Nested-PCR时, 取1uL直接PCR产物稀释40倍后作DNA模板, 用第二对引物R16F2/R2进行扩增, 其它条件同直接PCR。扩增产物用1 %~ 2 %琼脂糖凝胶电泳, 然后于紫外灯下检查1.5kb(直接PCR产物)和1.2kb(Nested-PCR)的特异扩增片段的有无。
2 结果与分析 2.1 枣疯病分布和为害现状及趋势调查结果显示枣疯病已遍布我国各大枣树栽培区, 但其分布状况和为害程度差异较大(见表 1)。
|
|
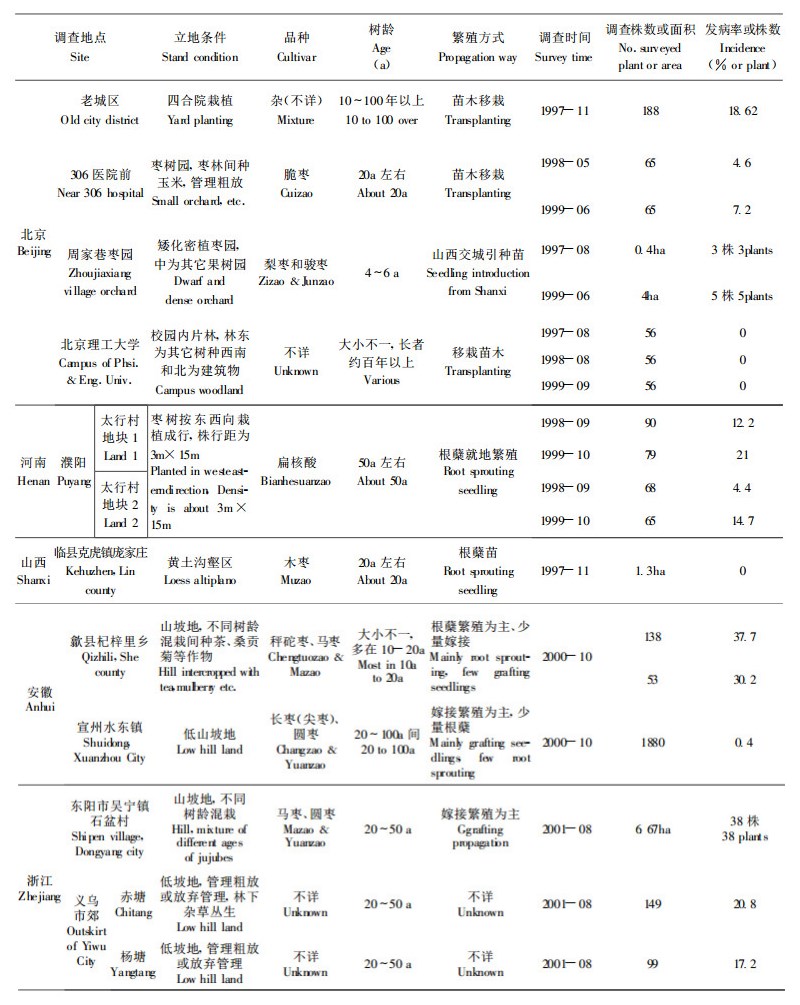
在调查的北京、河北、河南、陕西、山西、浙江和安徽都存在病害较严重发生和为害区或受害枣园。从病害分布范围看, 许多发病历史较长的老枣园, 由于病株基数大, 每年新发病株比例也明显高于病株率较低或仅出现零星病株的枣园。不同地区、同一地区的不同枣园间的病株率和为害程度差异很大。所调查的河南濮阳市、陕西清涧县、安徽歙县等地严重发病区, 特别是重病枣园, 并呈现出为害程度逐年加重的趋势, 在这些地区, 枣疯病已严重威胁到现存成年及中幼龄枣树的生存、枣果产量和质量, 成为这些地区红枣发展的主要限制因子。从本次调查结果看, 病害的分布范围基本与1989年的调查结果基本吻合(林业部野生动物和森林植物保护司, 1990), 即原来有枣疯病记载的地区, 病害多未被铲除或有效控制, 许多重病枣园的老枣树多已因病死亡, 或已造成整个枣园毁灭。少数病区, 象安徽宣州水东镇, 病害的扩展势头却不明显。此外, 也发现了原来未有枣疯病记载的枣树栽培区县已有零星病害发生, 如陕西佳县、浙江东阳市和山西临县等(表 2)。对这些地区出现的个别或零星分布病株的起源问题尚待进一步调查研究。
|
|
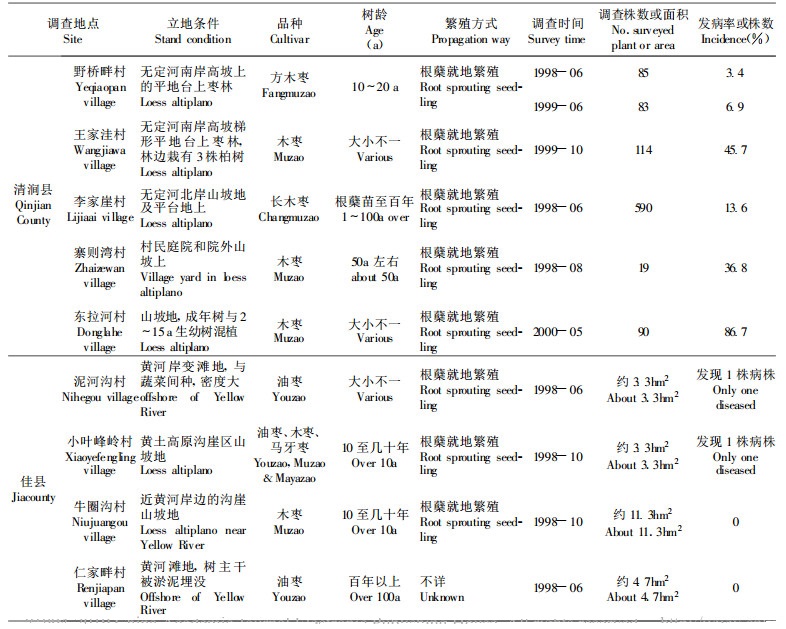
从调查结果看, 枣疯病传播存在三类典型代表方式, 即以陕西清涧为代表的黄土高原传播类型———以根蘖苗传播为主方式, 以河南濮阳为代表的平原农区传播类型———以介体叶蝉传播为主类型, 以及以新建枣园为代表的嫁接传播类型。
在清涧县的各重病枣园, 不同树龄的母、子树先后发病(包括母子根系相连和母树断根长出的根蘖苗两种类型)所占比例很高。大量的断根萌生的根蘖苗即表现为典型的全株丛枝症状。在以根蘖苗繁殖方式为主的枣园, 病树多呈团簇状分布, 病株数少至几株, 多至几十株, 其间仍有少数尚未发病的健株存在。在对该县北山里村的约2m距离的两株病树进行挖根调查发现, 其根系相连, 为典型的母子传病类型。安徽歙县杞梓里乡重病枣园、河北唐县等也存在此传播方式。
在河南濮阳太阳村重病地块, 介体昆虫传播(叶蝉)明显占主导地位, 表现为相似树龄的成年结果树的发病率稳定升高, 累计死株数目逐年增加; 病株簇状或条块分布明显, 病株基数大的地块, 新发病株增加也快, 表现为发病率加速增高的趋势。
从表 3中也可以得出, 嫁接育苗也是枣疯病传播的另一重要途径。由于已知酸枣(Z. spinosa Hu.)种子不传病, 在清涧县苗圃内, 当年嫁接苗即出现病苗显然是接穗带菌所致。其中夏二十里铺乡营田村用取自病区的枣树枝条作为接穗时, 因带病接穗比例较高而导致数量较多的嫁接苗发病; 而且, 既使取自无症状的枣树也未能避免接穗的带菌。在当地枣疯病很轻的佳县苗圃, 归圃育苗和采自本地的接穗嫁接苗皆未出现发病株, 这显然与接穗不带菌直接相关。
|
|
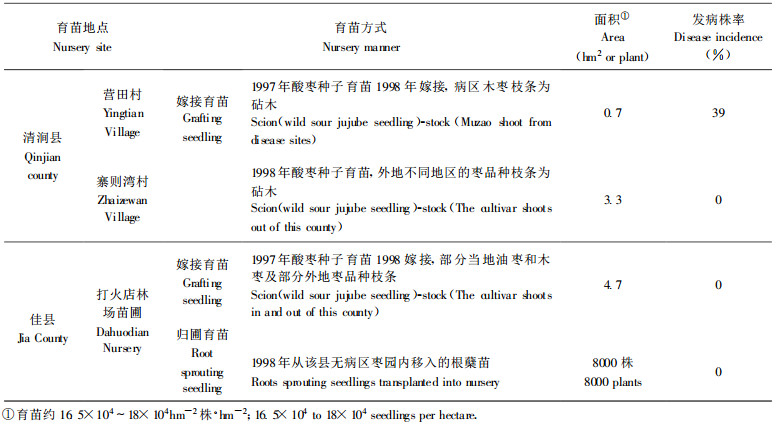
在安徽宣州市水东镇卢柴冲村, 自20世纪80年代开始, 当地即开始了有计划地用酸枣种子苗为砧木就地取成年枣树枝条作接穗进行嫁接育苗, 由于原有的病株率较低、毒源基数低, 至今, 嫁接的枣树(多为20a左右树龄)总体发病率仍较低, 原来的多数老枣树多生长和结果正常。
从病害侵染的毒源植株调查情况来看, 除了发病的栽培枣树外, 野生酸枣也是许多枣疯病发生区的不可忽视的病害侵染来源, 在河北唐县、陕西清涧县和北京密云县和海淀区的枣园周围皆发现酸枣丛枝病株存在。其中, 唐县郓城枣园周围的山坡上酸枣树发生丛枝病株的比率较高。另外, 在陕西清涧县也观察到与松柏树靠近的枣园枣疯病严重发生的现象。
2.3 枣树的田间抗病性及抗性丧失问题从不同品种自然感病率来看(表 4), 枣树品种间对枣疯病的自然抗性有明显差异, 其中梨枣表现出对枣疯病的高度感病, 而婆婆枣和洪赵小枣表现出对枣疯病的高度抗性。梨枣作为一种优质鲜食品种, 如何避免其高度的感病特性带来的枣疯病威胁需要引起足够的重视。在陕西清涧, 也观察到品质优的方木枣, 其感病程度重于长木枣, 表现为发病树病原扩展迅速、死亡较快。因而可能在枣果品质和对枣疯病的抗病性方面存在一定矛盾。河南濮阳的扁核酸是当地的主载培品种, 从田间发病率、染病树的症状发展和病树死亡进程看, 也为不抗病品种之一(参见表 2)。
|
|
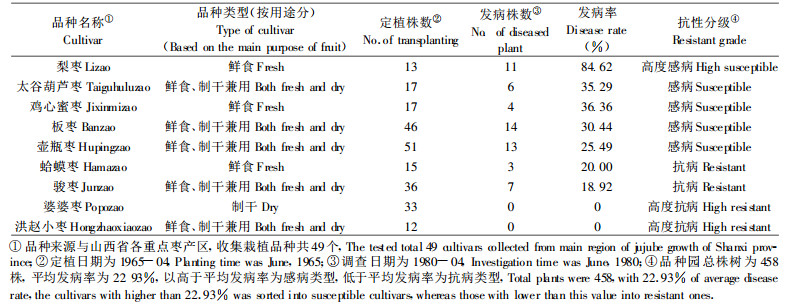
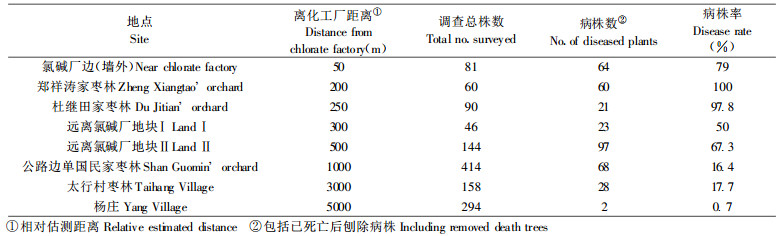
从表 5的调查结果中可以推断环境因素对某一品种抗病性的重要影响。离化工厂近的枣林, 丛枝病发病率和死株率明显增加, 这与此工厂曾出现过的氯气等有害废气排放导致枣树树体抗病性降低有直接的关系。植株本身固有的对植原体遗传抗病性的下降导致了植株抗侵染能力的下降, 或造成原来已被感染但尚处于潜伏期的带菌树体抗性丧失, 病原得以迅速大量繁殖而提早发病; 有害气体对病树的进一步的伤害, 又加速了树势衰退和病树死亡。
|
|
不合理的施肥也对枣树抗病性的丧失或病害的发展起明显的促进作用。在陕西清涧县和安徽歙县山地栽植的枣树, 普遍存在连年偏施纯氮肥(碳酸氢氨和尿素等)的问题, 这可能是造成枣树营养供应不全或不足和土壤有机质不断下降而导致枣树抗病性下降的重要因素(孙曦, 1997)。在安徽水东的枣园, 病害的扩展速度较低, 可能与当地一直重视施用土杂肥, 避免了偏施氮肥的管理传统不无关系。
2.4 田间枣树样品的带菌检测结果采用DAPI荧光显微镜技术可以从采自北京、河南濮阳、安徽歙县和宣州、陕西清涧和佳县、河北唐县等地的表现典型丛枝症状的叶柄、叶脉、变态花梗、幼枝等部位的韧皮部位筛管内检测到大量兰绿色植原体—DNA特异荧光(图 1A), 而健康对照则无此特异性荧光。其中, 病株丛枝部位的植原体—DNA荧光最强; 从纵切片中, 可以观察到许多被植原体—DNA荧光充满的细长筛管细胞, 在其它薄壁细胞中观察不到此特异性荧光。在病树上的黄化枝条中检出植原体的机率很低。但此技术对尚未表现丛枝症状的无症带菌枝条, 检测准确性也较低。

|
图 1 田间采集枣树样品的植原体检测结果 Fig. 1 Detection of phytoplasma from the field-collected samples of jujube trees A : DAPI荧光显微镜检测DAPI fluorcscence microscopy A1:枣疯病树丛枝部位幼茎组织横切片, 韧皮部具植原体特异荧光云Young stem cross cection of witches' broom part, showing specific phytoplasma fluorescence in the phloem; A2:健康枣树幼茎横切片, 韧片部无特异荧光Healthy control; A3:枣疯病树幼茎纵切片, 筛管内发植原体特异荧光Longitudinal section of young stem of witches' broom part, showing emitted fluorescence from sieve tubes. P —韧皮部; x —木质部, s—筛管; n—细胞核; p—phloem, x—xylem, s—sieve tube, n—cell nucleus B:枣树无症状部位的PCR检测结果(1998年9月河南濮阳太行村枣园取样); B : PCR detection to the symptomless trees or part of diseased trees(collected from Taihang village, Puyang, Henan on September, 1998) B1, 11:用枣疯病叶片组织抽提DNA为模板, 经直接PCR扩增出1.5kb片段(B11)和Nested-PCR扩增出1.2kb片段(B1) From leaf DNA templates of diseased part showing witches' broom symptoms, 1.5kb fragments was amplified by direct PCR(B11) and 1.2kb fragment was amplified by Nested-PCR(B1); B2, 4, 5, 7, 8和9:分别为不同病树上的无症状部位随机取样叶片DNA的Nested—PCR结果, 其中B4和B7扩增出1.2kb片段, 其余为阴性4, 5, 7, 8 and 9: nested-PCR results of symptomless leaves collected at random from different diseased trees respectively, 1.2kb DNA fragments were amplified from B4 and B7, others negative; B3和B6为靠近病树的无症状树的取样检测结果, 其中B6扩增出1.2kb片段, 此无症状树于2000年7月份发病; B3 and B6: nested-PCR results of symptomless trees very close to diseased trees, 1.2kb DNA fragment were amplified from B6, in addition, this tree expressed typical witches' broom symptoms in June, 2000; B10:未产生丛枝症状、仅表现单纯为全株叶片黄化症状的枣树用直接和Nested-PCR皆未检测到植原体侵染No phytoplasma infection was detected from the jujube tree showing yellow leaves of whole plant and without witches' broom symptoms; B12: PCR-DNA分子量标准, B12: PCR-DNA markers |
采用R16mF2/R2引物对进行的直接PCR可扩增出上述不同病样中的枣疯病植原体的1.5kb片段DNA, 而用R16F2/R2引物对进行的巢式PCR可扩增出1.2kb片段。其中巢式PCR可检测出用直接PCR不能检测出的极低浓度的植原体感染, 为无症带菌材料的检测提供了新的有效工具, 并很好地弥补了DAPI技术的缺陷(图 1B)。
在对北京病枣树(编号为J1)的不同部位或器官取样检测结果发现, 用直接PCR和巢式PCR皆可从丛枝部位检测出1.5kb特异扩增带; 用直接PCR未能从病树上的一无症花枝检测出1.5kb扩增带, 而进一步用巢式PCR则检测出1.2kb扩增带, 说明此部位已存在低浓度的植原体; 一无症花枝上的小花样品中未检测出病原; 另外病树上采集的无症根蘖苗枝条和一无症营养枝中也未检测出病原。与J1病枣树相邻的三株未发病枣树样品也未检测出植原体感染。对陕西清涧县李家崖一取样地点的病树周围无症树的随机取样检测结果发现, 12个样品皆未检测出植原体的感染; 另一样地的一株病株四周的四株无症树中有一株检测到植原体感染。
用PCR对1999年9月20日取自河南濮阳太行村病株无症枝条和与病树相邻的无症状枣树上的叶片进行的检测的结果显示, 所有10个样品用直接PCR皆未扩增出植原体特征带, 当进一步进行巢式PCR时, 有2株病树上的无症状枝条样品检测出植原体的感染, 其中一株为与病株相邻的无症株。至2000年7月份对无症带菌株发病情况的追踪调查显示, 原来的一株无症带菌株已发病, 并表现出典型丛枝症状, 从检测到病原至表现症状的潜育期达1年半以上。这是枣树存在无症带菌株的更直接证据(图 1B)。另外, 用PCR确证陕西佳县泥河沟村和小叶峰村的各一株可疑的枣疯病病树的病原, 从而校正了过去佳县未有枣疯病记载的历史。
从采自佳县和清涧县苗圃的54个无症嫁接苗样品中均未检测出带菌苗木, 这一结果可能意味着如果是带病接穗, 嫁接到酸枣种子苗砧木上后一般当年发病, 而未表现典型丛枝症状的嫁接苗的带菌几率很低; 也可能是本次试验所采集和测定的样本较少、尚不能完全代表整个苗圃的带菌状况或存在漏检等。
3 讨论从所调查的重病区来看, 自从20世纪80年代开始, 伴随着枣树繁育速度的加快和枣果产量的提高, 枣疯病的为害也随之加重。根据调查结果分析, 病害加重的原因可能主要涉及病害传播频率的增加和枣树抗病性的普遍降低两个方面。
首先, 就某一地区来看, 一直沿用至今的就地根蘖繁育方式和民间种苗交流方式, 不断地扩大着病园原有病株的基数, 并将病株带入原来无病的枣园。进一步的介体昆虫传播使零星发病区很快变成了轻病区或重病区; 原来未有枣疯病发生的枣园也出现了病株。以上传病过程, 经过多年的累加效应而达到迄今的严重为害程度。进入20世纪90年代, 全国各枣区间品种的引进和种源交流数量和频率明显增加, 使病害随种苗和其它营养繁殖材料(主要为接穗)进行长距离人为传播的几率增加, 导致了枣疯病的发生范围扩大和为害程度的进一步加重(田国忠, 1998; 1999)。因而, 如何更全面、系统地了解当前我国枣疯病的分布、危害程度和蔓延趋势是值得进一步开展的研究课题之一。
关于枣疯病的传病介体叶蝉, 国内外已有许多的报道(陈子文等, 1984;孙淑梅等, 1988;王焯等, 1981; 1983)。但象清涧、佳县、延川等黄土高原地区、安徽歙县、浙江义乌市和东阳市、湖南浦县等枣疯病发生较重的南方山区的传病介体及特性方面仍缺乏针对性研究。这在一定程度上影响了对在这类地区病害流行趋势的预测和具体防治措施的制定。本次调查已发现在清涧县枣园附近载种零星柏树的地块, 枣疯病危害最为严重, 故推测松柏树可能是该地区传毒叶蝉的越冬场所; 而且在黄土高原沟壑区, 介体昆虫的传播效率和流行速度可能会低于其它平原枣区(孙淑梅等, 1988;田国忠等, 2000)。
从本次调查结果来看, 枣树对枣疯病植原体的抗性不仅涉及品种间遗传抗病差异, 而且环境因子对品种抗性的影响。一方面, 从初步的自然抗病鉴定调查结果看, 我国作为枣树的起源地和主要栽培地, 记载的品种多达700余个, 且发现了许多表现明显的田间自然抗病的种质资源(曲泽洲等, 1993)。因而, 如何深入开展定向抗病育种工作, 培育将抗病种质与优良品质和栽培性状相结合的新品种具有重要意义。由于田间自然传播鉴定存在需要时间长、接种机率不均一、无法区分抗介体传播及抗病原扩展等不同抗病类型等缺陷, 决定了有必要对通过自然鉴定获得的抗病种质材料用嫁接、介体人工传毒、生化指标鉴定及抗病分子标记等途径进行进一步的诱发抗病鉴定及开展抗病机制等方面的研究1)(田国忠等, 1994; 1999)。另一方面, 环境胁迫会导致枣树抗病机能的明显丧失在枣疯病的发生和流行中所起的作用也不容忽视。某些生产上曾被普遍接受的或片面提高产量措施和栽培管理不善(比如, 树皮环剥、喷施提高座果剂、偏施肥料、过度伤根等等)常常会造成树势衰退、抗病性降低、进而导致病株率增加。特别是因局部或系统性空气、水体或土壤污染与诱发无症带菌枣树发病, 及与整个枣园病害流行之间的关系更是值得深入探讨的新问题。
1) 温秀军, 孙士学, 孙朝晖.抗枣疯病枣树品系的选择. 2000, 内部交流材料
过去, 由于枣树室内苗期嫁接接种病原试验不能完全模拟不同地区枣林介体昆虫的自然传播模式, 或无条件开展用各种树龄枣树进行系列接种试验, 因而对不同严重程度病园内存在的病株、无症带菌株和健康植株三者的比例, 枣树不同品种从被传毒至发病的时间(潜育期)、无症带菌株表现症状所需的条件等问题皆缺乏深入的了解。本研究已初步肯定了巢式PCR在无症带菌株调查中的价值和作用, 因而进一步对枣园内枣树无症带菌状况开展调查研究, 不仅可澄清一些长期悬而未决的理论问题, 而且对于枣疯病流行预测预报和指导病害的综合防治, 以及避免引种过程中病害的人为传播等都有深远的意义(Griffiths et al, 1994 : Jarausch et al, 1998; Martini et al, 1999)。
陈子文, 张风舞, 田旭东, 等. 1984. 枣疯病传播途径的研究. 植物病理学报, 14(3): 141-146. |
何放亭, 武红巾, 陈子文, 等. 1996. 几种植物类菌原体(MLOs)的分子检测及遗传相关性比较. 植物病理学报, 26(3): 251-255. |
金开璇, 田国忠, 汪跃. 1989. 组织化学技术快速检测泡桐丛枝病研究. 植物病理学报, 19(3): 185-188. DOI:10.3321/j.issn:0412-0914.1989.03.001 |
林业部野生动物和森林植物保护司主编.森林植物检疫对象和检疫技术.长春: 吉林科学技术出版社, 1990
|
罗飞, 朱水芳, 张成良等.泡桐丛枝病媒介昆虫体中植原体的检测和鉴定.见: 刘仪主编.植物病害研究与防治.北京: 中国农业科学出版, 1998, 280~ 283
|
曲泽洲, 王永惠主编.中国果树志: 枣卷.北京: 中国林业出版社, 1993
|
孙淑梅, 张风舞, 田旭东. 1988. 枣疯病的煤介昆虫—凹缘菱纹叶蝉生物学和防治研究. 植物保护学报, 15(3): 173-177. |
孙羲主编.植物营养原理.北京: 中国农业出版社, 1997
|
田国忠. 1999. 北京市木本植物植原体病害发生及防治对策. 北京农业科学, 17(6): 25-28. |
田国忠, 熊耀国, 汪跃, 等. 1994. 泡桐对丛枝病原MLO的抗性研究. 林业科学研究, 7(2): 155-161. |
田国忠, 张锡津, 罗飞. 1999. 抗病和感病泡桐无性系组培苗对嫁接传染植原体的不同反应. 林业科学, 35(4): 31-39. DOI:10.3321/j.issn:1001-7488.1999.04.006 |
田国忠, 张志善, 郭加虎, 等. 2000. 陕西清涧县枣疯病发生和为害调查及防治建议. 陕西林业科学, (2): 46-51. |
田国忠. 1998. 枣疯病的预防和治疗策略研究. 林业科技通讯, (2): 14-16. |
田砚亭, 王红艳, 牛辰, 等. 1993. 枣树脱除类菌原体(MLO)技术的研究. 北京林业大学学报, 15(2): 20-26. DOI:10.3321/j.issn:1000-1522.1993.02.001 |
王焯, 于保文, 周佩珍, 等. 1981. 枣疯病传毒昆虫研究I. 传毒昆虫—中国拟菱纹叶蝉.植物病理学报, 11(3): 25-29. |
王焯, 张承要, 周佩珍, 等. 1983. 枣疯病传播昆虫分布调查. 植物病理学报, 13(3): 174. |
王祈楷, 除绍华, 陈子文, 等. 1981. 枣疯病的研究. 植物病理学报, 11(1): 15-18. |
中国科学院上海生物化学研究所. 1974. 枣疯病病原体的电子显微镜研究. 中国科学, (6): 622-625. |
Griffiths H M, Sinclair W A, Davis R E, et al. 1994. Characterization of mycoplasmalike organisms from Fraxinum syringa and associated plants from geographically diverse sites. Phytopathol, 84(2): 119-126. DOI:10.1094/Phyto-84-119 |
Jarausch W, Lansac M, Saillard C, et al. 1998. PCR assay for specific detection of European stone fruit yellows phytoplasmas and its use for epidemiological studies in France. Eur. J Plant Pathol, 104(1): 17-27. |
Kim C J Witches' broom of jujube tree, Ziziphus jujuba Mill.var. enermis Rehd. Transmission by grafting. Korean J. Microbiol, 1965, 3(1): 1~ 6
|
La Y J.and Lee D J. Distribution of mycoplasma in witches' broom infected jujube tissue. J. Korean Forestry Soc, 1984, 67(1) : 28~ 30
|
La Y J. Current status of researches on mycoplasmal diseases of plants in Korea. In: Raychaudhuri S P & Rishi N ed. Mycoplasma Diseases of Woody Plants New Delhi : Malhotra Publisher House, 1987, 117~ 128
|
Martini M, Murari E, Mori N, et al. 1999. Identification and epidemic distriction of two flavescence doreerelated phytoplasmas in Veneto(Italy). Plant Dis, 83(10): 925-930. DOI:10.1094/PDIS.1999.83.10.925 |
Pandey P K, Singh A B, Nimbalkar M R, et al. 1976. A witches' broom disease of jujube from India. Plant Disease Reporter, 60(4): 301-303. |
Tian Guozhong, Zhishan Zhang, Fei Luo et al.An Investigation of jujube witches' broom caused by phytoplasma in different regions of China. In: Zhou Guanghe and Li Huaifang ed.Proccedings of the First Asian Conference on Plant Pathology. Beijing : China Agricultural Scientech Press, 2000, 177.
|