 2001, Vol. 37
2001, Vol. 37文章信息
- 刘金福, 洪伟.
- Liu Jinfu, Hong Wei.
- 福建三明格氏栲林物种多度分布格局研究
- STUDY ON SPECIES ABUNDANCE DISTRIBUTION OF CASTANOPSIS KAWAKAMII NATURAL FOREST IN FUJIAN SANMING
- 林业科学, 2001, 37(专刊1): 200-204.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 200-204.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


格氏栲(Castanopsis kawakamii Hayata)属国家二级保护植物, 是中国中亚热带南缘特有的壳斗科常绿阔叶大乔木、珍稀濒危植物之一。其自然分布范围较窄, 主要分布于福建、台湾、广东、广西、江西等地区, 多零星生长在海拔200~1000 m的丘陵地带的常绿阔叶林中。但在福建三明小湖地区有近700 hm2以珍稀濒危植物格氏栲占优势的林分, 多由百年以上的大树组成, 是中国目前罕见的天然群落类型。
格氏栲林以号称“凤毛麟角”的独特自然森林景观, 引起许多生态专家、学者的极大关注:1958年郑万钧教授建议建立格氏栲自然保护区(章浩白, 1993);1962年阳含熙对天然格氏栲解析木进行调查研究(王本楠, 1987);1979年阳含熙等对格氏栲林植物群落展开数量分类的研究(阳含熙等, 1979);1986年林鹏等对格氏栲林群落结构特征进行分析(林鹏等, 1986);1989年邱喜昭等定性分析格氏栲属闽中部亚地带的区域代表树种(邱喜昭等, 1989);1994年游水生等探讨格氏栲自然保护区植被类型的划分(游水生等, 1994);1997~2001年刘金福、洪伟等系统开展格氏栲种群生态学研究工作, 揭示格氏栲种群数量增长动态、生态位、生命过程、空间格局、种内种间竞争、密度调节等方面规律(刘金福、洪伟等, 1998a~1998c;1999a~1999g;2001) 1); 2000年樊后保进一步分析格氏栲群落结构特征(樊后保, 2000);2000年吴承祯等提出格氏栲种群数量种群增长新模型(吴承祯等, 2000), 等等。
1)刘金福.格氏栲种群动态变化规律的数量分析研究.硕士毕业论文, 1997.
考虑到格氏栲林古老性和稀有性, 以及格氏栲天然更新越来越困难, 研究格氏栲林生物多样性具有深远的生态意义和实用价值。但迄今为止, 有关格氏栲林物种多样性研究仅局限于采用多样性指数简单描述群落的变化(林鹏等, 1986;樊后保, 2000), 忽略了格氏栲林富集种和稀有种对群落多样性贡献的差异, 而采用群落种—多度的观测频度分布寻找某种适合的理论分布来预测种—多度分布形式, 即格氏栲群落内部物种多度分布格局的研究尚属空白。笔者试图在前人研究基础上, 采用物种多度分布格局模型讨论明格氏栲林群落的物种多样性, 以此拓宽格氏栲林物种多样性研究的数量分析方法。
目前, 最常用的分布模型有五类(Magurran, 1988;谢晋阳, 1993;洪伟, 2000) :Whittacker提出的几何分布模型(The geometric series model, 1964);MacArthur提出的断棍分布模型(The broken-stick model, 1957);Fisher等提出的对数分布模型(The log series model, 1943);Preston提出的正态对数分布模型(The log-normal distribution model, 1947, 1962);吴承祯等提出的物种多度分布的Weibull分布模型(吴承祯等, 1997)。由于对数分布可反映物种以无规则的时间间隔侵入生境, 并反映出一个或几个物种在群落中占优势, 表明随着演替的进行, 环境条件逐渐改善。因此, 生态学者认为:成熟的自然群落物种多度分布呈对数分布(Magurran, 1988;谢晋阳, 1993)。根据格氏栲林生态特征分析(林鹏等, 1986), 格氏栲林是具有相对稳定性较大的天然群落, 故本文运用对数分布模型来反映格氏栲林的生物多样性—物种多度分布规律, 探索其变化规律, 为我国珍稀濒危植物格氏栲林的群落演替、稳定性评价、物种多样性保护、开发利用和丰富的基因资源合理经营管理提供科学依据。
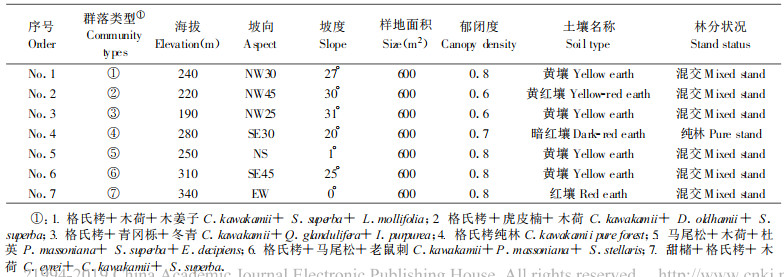
1 调查区自然概况格氏栲林位于三明市郊西南部, 面积近700 hm2, 地处26°07′~26°10′ (N)、117°24′~117°27′ (E), 该区海拔高度为180~604 m, 属福建武夷山东伸支脉地带, 其东南方为戴云山脉; 该区气候属中亚热带季风型气候, 年均温度19.5℃, 极端最低气温为-5.5℃, 最高气温40℃, > 10℃年积温为6215℃; 年平均降雨量为1500 mm, 3~8月的降雨量约为全年的75%;年平均相对湿度79%;年平均风速1.6 m·s-1; 土壤类型主要为暗红壤, 其次为红壤和紫色土, 土层较厚, 土层腐殖质丰富, 水肥条件均较好, 植物种类丰富, 群落类型多样, 林冠层呈波浪状, 郁闭度高达0.8左右, 格氏栲树冠常年浓绿, 冠幅十分庞大, 树形优美通直, 树皮呈灰褐色, 林相整齐, 呈黄绿色, 整个群落浓郁苍翠, 形成中亚热带常绿阔叶林所特有的貌特征。由于海拔低, 无明显垂直分布带, 格氏栲多数分布在海拔450 m以下(刘金福等, 1999a)。从整体上看, 格氏栲天然林植被自然性较强, 格氏栲林生态功能发挥较充分, 生态系统无明显的结构变化, 生境基本完好(刘金福等, 1999g)。
2 研究方法 2.1 野外调查在以格氏栲占优势的林分中, 调查7种主要群落类型, 各设置1块20 m×30 m的典型样地, 测定每块样地海拔、坡向、坡位坡度、土壤条件和群落类型等因子及记录乔木树种的种名、树高、胸径(起测径阶≥4 cm); 在每块样地内采用相邻格子法设置面积为5 m×5 m的小样方24个, 统计每个样方样方内所有乔木个体(胸径4 cm以下)、灌木、幼苗的种类和个体数; 在各样地内设置4个1 m×1 m的小样方调查草本分布、数量。调查资料整理后, 编制成森林植被调查报告, 并建立数据库输入计算机存档。调查样地概况见表 1。
|
|
(1)物种多度分布的对数分布模型的计算 对数分布模型是由物种在每一多度级以上的物种数量即物种频率(Sn)给出:

|
(1) |
式中:Sn为多度n的物种数量, a、X为参数。为了得到群落的对数分布模型, 就必须得到参数a和X的值。参数X的值可由下列公式求得:

|
(2) |
式中:S为群落中物种的总数; N为群落中个体的总数。X的值比0大而小于1, 如果S/N < 0.05, 则X > 0.99 (Magurran, 1988)。第二个参数a则由参数X得来(Magurran, 1988) :
|
(3) |
这样, 得出了参数a和X的值, 就可得到物种多度分布的对数模型。
(2)多度分布的检验 按Thomas和Shatteck (1986)的方法, 对多度的观察值进行分级, 并选择以2为底的对数(即种群多度的加倍), 作为各个多度级的上限值, 然后各上限值再加0.5以使各级之间的界限更明显了(Magurran, 1988)。首先对各个种的个体数用对数级数进行预测, 再对预测的结果进行检验。①预测对数级数分布以下式预测具有一定个体数的物种数目:

|
(4) |
aX为具有1个个体的物种数目, aX2/2为具有2个个体的物种数目, aX3/3为具有3个个体的物种数…, aXn/n为具有n个个体的物种数目。其中参数a和X据(2)式和(3)式推算。将参数a和X代入(4)式中便可计算出各个体数的物种数目预测值。然后, 据公式:
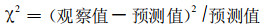
|
(5) |
求得χ2值及∑χ2值。②检验按自由度=多度级-1, 查卡平方(χ2)检验表, 估计精度取0.95 (α=0.05), 检查观察值与预测值之间的差异显著性, 以检验其分布。
(3)图解分析方法 “多度/频度”图解是由Preston (1948、1962、1963)提出的, 是用物种的数量作y轴(频度), 而以每一个种的多度级为x轴。x轴的多度级可以采用一个合适的量值, 且对轴进行以2为底的对数转换。为了从直观上说明格氏栲林物种多度分布格局, 本文提出采用“多度/累积概率”图解来表达格氏栲林的群落物种多度的对数模型更为直观一些, 绘制出每一种群的“物种多度—累积概率”图。
首先将样本数据按多度由小到大排列:X=(X1, X2, …, Xn)。这样分布函数定义为:Tn(X)=(X中小于或等于Xi, 的物种个数)/n, 然后在概率纸上做出曲线, 其数学形式为:

|
(6) |
它是随机变量分布函数p(X)的近似形式, 当n→∞, 有Tn(X)→p(X)→1。
3 结果与分析 3.1 对数分布模型的计算应用对数分布模型对格氏栲林7种群落的乔木层、灌木层和所有木本植物的物种进行物种多度分布的对数模型的计算, 将各群落中的乔木树种、灌木树种所有木本植物的有关数据分别代入(2)式和(3)式, 求得各群落的种群多度分布参数a和X(见表 2)。
|
|

将a与X值代入(4)式计算出各个体数的预测物种数目, 并据(5)式计算得到∑χ2值, 再查出χ0.052的临界值, 检验结果见表 3。
|
|

经分析, 7种群落乔木层物种分布的不均匀性较为明显, 物种间个体数的差异显著, 其多度级由4级至5级, 其中4级的占了57%;而各群落灌木层物种分布的不均匀性更为明显, 物种间个体数的差异更加显著, 其多度级由6级至7级, 其中6级的占了57%。从整体上看, 各群落所有木本植物物种分布的不均匀性极为明显, 物种间个体数的差异更大, 其多度级具有7级的比例占了85.7%, 7种群落占6种。无论何种层次, 经卡方(χ2)分布检验, 格氏栲林7种群落乔木层、灌木层、所有木本植物的物种多度分布的观察值和预测值之间均无明显差异(p < 0.05), 物种多度分布均很好适合于对数分布。
可见, 格氏栲林各群落不同层次物种多度分布明显差异, 乔木层物种分布相对来说较灌木层、所有木本植物物种分布的适合性差点, 以物种的个体数量来衡量, 乔木层多度级数相对小, 只介于4~5级, 物种个体数不均匀分配程度相对较弱, 而灌木层、所有木本植物的非均匀程度特别明显, 即群落物种均匀度相对较小, 表明这些层次主要由少数种群所控制, 与樊后保多样性指数分析结果是一致的(樊后保, 2000), 这可能与格氏栲种群多由百年以上的老年个体组成, 树冠开始变稀有关。
3.4 图解分析利用对数分布的分布模型, 并求出与Xi相对应的Sn (Xi)值, 进行累积, 然后根据(6)式在概率纸做出对数曲线, 见图所示(限于篇幅, 只绘制一个群落即格氏栲+青冈栎+冬青群落的乔木层、灌木层及所有木本植物物种的“多度分布/累积概率图”), 可直观地看出格氏栲林的物种多度分布格局符合对数分布。

|
图 1 观察曲线与对数分布曲线的比较 Fig. 1 Comparison between observed curve and log series distribution curve A:乔木层Tree layer; B:灌木层Shrub layer; C:所有木本植物All woody species.观察曲线observed curve; ----------对数分布曲线log series distribution curve. |
从林业持续发展和生物多样性保护原则出发, 物种多样性应是计量评价格氏栲林综合效益的重要指标之一, 而物种数量与物种多度的关系是描绘格氏栲林生物多样性均匀度水平的特征状态的一种重要度量方法.本研究运用物种多度的对数分布模型首次描述格氏栲林的生物多样性, 从而丰富了格氏栲林物种多样性的研究结果, 也为格氏栲濒危机理及保护提供一些可借鉴的统计信息。研究结果表明, 格氏栲林不同主要群落的乔木层、灌木层及所有木本植物的物种—多度分布均很好符合对数分布, 表明用对数分布模型描述格氏栲林的物种—多度格局是可行的; 同时采用“多度/频度”图解方式来探讨物种多度分布的对数分布, 收到满意效果。
格氏栲林是介于中亚热带与南亚热带的一种常绿阔叶林过渡类型, 物种丰富, 结构复杂, 但优势种群的作用较明显, 优势乔木种群格氏栲控制整个群落环境, 导致格氏栲林群落中物种个体数间的差异较大, 优势种的个体数会明显多出一般种而使整个群落具有较低的均匀度, 即格氏栲林整体上物种均匀度相对较小, 组成中稀有种较多, 因而格氏栲林各层次物种-多度格局明显表现对数分布。同时研究表明格氏栲林为相对发育成熟的森林群落, 具有较大的稳定性和较高的生物多样性。
樊后保. 2000. 格氏栲群落的结构特征. 林业科学, 36(2): 6-12. DOI:10.3321/j.issn:1001-7488.2000.02.002 |
洪伟. 2000. 闽江流域森林生态研究. 厦门:厦门大学出版社, 50. |
林鹏, 丘喜昭. 1986. 福建三明瓦坑的赤枝栲林. 植物生态学与地植物学学报, 10(4): 241-253. |
刘金福, 洪伟, 李家和. 1998a. 格氏栲群落生态学研究Ⅱ .格氏栲林主要种群的竞争研究. 福建林学院学报, 18(1): 24-27. |
刘金福, 洪伟, 李家和, 等. 1998b. 格氏栲种群生态学研究Ⅲ .格氏栲种群优势度增长动态规律研究. 应用生态学报, 9(5): 453-457. |
刘金福, 洪伟, 李茂瑾. 1998c. 格氏栲群调节模型研究. 热带亚热带植物学报, 6(4): 309-314. |
刘金福, 洪伟. 1999a. 格氏栲群落生态学研究.格氏栲林主要种群生态位的研究. 生态学报, 19(3): 30-36. |
刘金福, 洪伟. 1999b. 格氏栲种群个体年龄与胸径的时间序列模型研究. 植物生态学报, 23(3): 283-288. |
刘金福, 洪伟. 1999c. 格氏栲种群增长动态预测研究. 应用与环境生物学报, 5(3): 247-253. |
刘金福, 洪伟, 陈清林. 1999d. 格氏栲种群生态学研究Ⅴ .格氏栲种群空间格局及其动态的研究. 福建林学院学报, 19(2): 118-123. |
刘金福, 洪伟. 1999e. 格氏栲种群生态学研究Ⅵ .格氏栲种群空间格局分布的 Weibull 模型研究. 福建林学院学报, 19(3): 212-215. |
刘金福, 洪伟. 1999f. 格氏栲种群生态学研究Ⅶ .格氏栲种群分布格局的强度与纹理分析研究. 中南林学院学报, 19(1): 59-63. |
刘金福, 洪伟. 1999g. 福建三明格氏栲自然保护区评价. 吉林林学院学报, 15(2): 70-73. |
刘金福, 洪伟. 2001. 格氏栲种群优势度增长改进模型的研究. 植物生态学报, 25(2): 225-229. DOI:10.3321/j.issn:1005-264X.2001.02.013 |
邱喜昭, 林鹏. 1989. 福建中亚热带常绿阔叶林壳斗科树种的水平分布特点. 植物生态学与地植物学学报, 13(1): 36-42. DOI:10.3321/j.issn:1005-264X.1989.01.007 |
王本楠. 1987. 崔-Lawson 种群模型的显示解及拟合实例. 生态学杂志, 6(2): 27-30. |
吴承祯, 洪伟. 1997. 观光木群落物种多度分布的Weibull 模型研究. 福建林学院学报, 17(1): 20-24. |
吴承祯, 洪伟, 陈辉, 等. 2000. 珍稀濒危植物青钩栲种群数量特征研究. 应用生态学报, 11(2): 173-176. DOI:10.3321/j.issn:1001-9332.2000.02.004 |
谢晋阳. 1993. 物种多样性指数与物种多度分布格局. 植物科学综论: 222-233. |
阳含熙, 卢泽愚, 杨周南. 1979. 植物群落数量分类的研究. 林业科学, 15(4): 244-255. |
游水生, 郭振庭. 1994. 模糊聚类探讨福建三明格氏栲自然保护区植被类型的划分. 武汉植物学研究, 12(4): 333-340. |
章浩白. 1993. 福建森林. 北京: 中国林业出版社, 107-116.
|
Magurran A E. 1988. Ecological diversity and its measurement .Princeton University Press. Sydney : Croom Helm, 1-79.
|