 2001, Vol. 37
2001, Vol. 37文章信息
- 陈礼光, 郑郁善, 邱尔发, 周小华, 吴擢溪.
- Chen Liguang, Zheng Yushan, Qiu Erfa, Zhou Xiaohua, Wu Zhuoxi.
- 米槠和苦槠LN2保存材料细胞膜代谢特征研究
- STUDY ON METABOLIZATION CHARACTERS OF CELL MEMBRANE OF CRYOPRESERVATION MATERIALS OF CASTANOPSIS CARLESII AND C. KAWAKAMII
- 林业科学, 2001, 37(专刊1): 137-142.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 137-142.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


米槠(Castanopsis carlesii)和苦槠(C. kawakamii)是亚热带壳斗科常见建群种, 其种子是亚热带典型的顽拗性种子, 极不耐脱水(Rorberts, 1973)。由于其种子成熟过程中不经成熟脱水阶段, 仍保持有较旺盛的代谢活动, 使种子代谢方向很快地由休眠转向萌发方向发展, 脱水敏感性剧增, 种子的耐贮藏性丧失(Farrant et al., 1988; 王贵禧等, 1999)。在种质资源日益受重视的今天, 如何有效地保存和长期占有种质资源在1个国家的国民经济增长与发展中均占有重要的战略地位, 愈来愈受到国内外种质资源工作者的关注。米槠和苦槠种子不耐贮藏特性, 导致了其种质资源的长期保存困难性。这类种子的常规贮藏, 一般采用层积法贮藏(孙时轩, 1990), 但目前有研究报道指出, 采用-196℃超低温保存(又称液态氮保存, Liquid nitrogen, 简作LN2)是长期保存这类种子的最有发展前途的1种途径(陆旺金等, 1998; 王君晖等, 1998)。通过对米槠和苦槠两树种种子LN2保存后代谢特征, 主要是酶活性的研究, 以期为长期保存这两个树种种子的种质提供和积累有益的科学参考依据。
1 研究材料和方法 1.1 研究材料研究材料为米槠和苦槠两树种种子及其离体胚, 统称为保存材料。
1.2 研究方法 1.2.1 含水量(MC)将米槠和苦槠的种子, 在其种胚部不伤及胚的前提下, 切取约5 mm×5 mm×5 mm大小的胚方, 用30℃+硅胶条件下使其失水至相对含水量为20%、15%、10%、8%、5%、2%等各含水量梯度, 不失水的为自然含水量(NMC), 以不经液态氮处理的为对照.种子的各含水梯度处理与离体胚的类似。
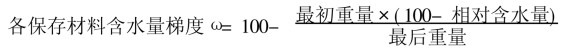
|
对材料进行各种冷冻处理, 然后进行LN2保存, 各种解冻处理后(图 1), 进行发芽率、电导率、SOD活性和MDA含量测定。

|
图 1 冷冻和解冻流程 Fig. 1 Freezing and thawing process 冷冻方式(FM), 解冻方式(TM), 停留时间(STm), 快冻(QF), 缓冻(MF), 慢冻(SF), 快解(QT), 缓解(MT), 慢解(ST). Freezing methods (FM), Thawingmethods (TM), Staying time (STm), Quick freezing (QF), Mild freezing (MF), Slowfreezing (SF), Quick thawing (QT), Mild thawing (MT), Slow thawing (ST). |
参照1996年国际种子检验规程的规定(ISTA编, 1999)。并计算种子的发芽指数和活力指数。
发芽指数:GI=∑Gt/Dt。Gt:在时间t日的发芽数, Dt:相应的发芽日数
活力指数:VI=GI×L。L:幼根平均长度
1.2.4 电导率测定DDS-11A型电导率仪测定法(黄学林等, 1990)。
电导率(mΩ·cm-1g-1) = (电导测定值-空白) ×电极常数
然后换算为25℃时电导率(张宪政, 1992) :25℃电导率=电导率×[1+0.02× (t-25) ]
计算相对电导率:相对电导率Sr (%) =S1/S2×100。式中:S1为25℃电导率, S2为绝对电导率
1.2.5 超氧化物歧化酶(SOD)活性参照Giannoplitis和Reis (1977) (朱广谦等, 1990)。利用SOD抑制氯化硝基氮蓝四唑(NBT)在光照下的还原作用。3 mL反应系统中含0.013 mol·L-1蛋氨酸、63×10-6 mol·L-1 NBT、100×10-6 mol·L-1 EDTA、1.3×10-6 mol·L-1核黄素、0.05 mol·L-1 pH=7.8含1%PVP的磷酸缓冲液。4000 Lx的光强下照光15 min后, 测OD560值。SOD活性大小以每10胚抑制NBT光还原50%为一个酶活性单位。
1.2.6 丙二醛含量的测定硫代巴比妥酸法(朱广谦等, 1990)。含量以每10胚中含丙二醛mol数表示。
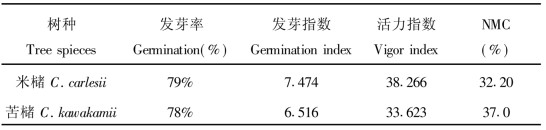
2 结果与分析 2.1 自然质量特性对米槠和苦槠两树种的种子, 进行发芽试验。各试验4个重复, 每重复50粒。计算出各树种的发芽率、发芽指数和活力指数(表 1)。新采收的两树种种子的原始发芽率较高, 分别为79%和78%, NMC均大于30%, 自然状态下2树种种子质量特性较相近。
|
|
在0~20%MC的范围内, 米槠种子的发芽率随含水量的降低而呈先升高后降低的规律, 其中12%MC的发芽率最高, 达50%, 10%~15% MC区间内, 发芽率差异并不显著; 在不同的冷冻和解冻处理中, 其中MF-QT方式比其他两处理的发芽率要高得多(图 2a)。苦槠的发芽率在15%MC MF-QT方式最高, 达48%, 其次是12%MC, QF-QT方式, 达43% (图 2b)。

|
图 2 LN2保存后米槠(a)和(b)苦槠发芽率 Fig. 2 Germination of C.carlesii (a) and C.kawakamii (b) after cryopreservation |
种子活力的大小与电导率的高低一般呈负相关, 活力高的种子, 其膜修补能力强, 电解质外渗量少, 积累速率慢, 活力弱的种子则反之。
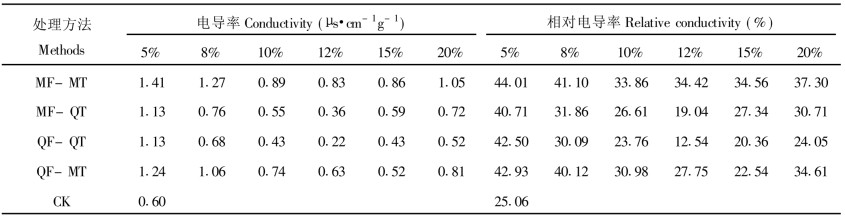
新鲜的米槠种子浸泡液的电导率每8 h为0.60 μs·cm-1g-1, 超低温保存后, 20%MC处理的种子相对电导率较高, 则说明冷冻和解冻过程会对20%MC米槠种子造成伤害。当米槠种子降低到12%MC, QF-QT的处理中出现一个电导率最低值, 而且MF-QT处理电导率也较低, 说明12%MC米槠种子基本未受到伤害而发生膜系统破坏。当米槠种子降低到8%MC时各处理的电导率有较大幅度地上升, 这表明米槠种子已受到较为严重的脱水伤害。采用12%MC和快解方式(45℃温水解冻)的效果最好(表 2)。
|
|
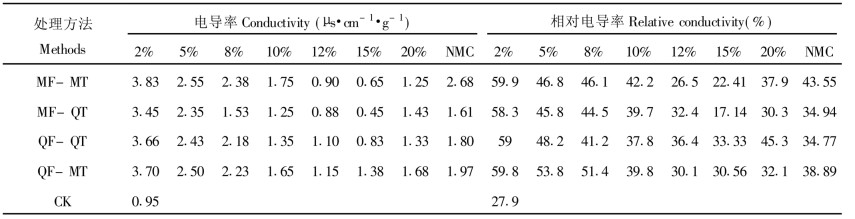
新鲜的苦槠种子电导率每8 h为1.79 μ·cm-1g-1, 苦槠种子超低温保存后, 其电导率值随含水量降低而下降, 到15%MC时达最小值, 而后又上升。MF-MT、MF-QT2处理的电导率要比QF-QT和QF-MT低, 冷冻方式对种子的超低温保存效果影响较解冻方式来得大, 这可能是冷冻过程中, 细胞产生保护性脱水, 而使解冻过程中受到的伤害减少到最小。经超低温处理的各种含水梯度的苦槠种子相对电导率随着含水量的变化规律是先降低后升高。经过液氮贮存的苦槠种子相对电导率都有下降的趋势。从中可以看出10%~15%MC的苦槠种子经液氮贮存后相对电导率与CK比变化不大。且经MF-QT处理的效果好于MF-MT和MF-QT (表 3)。
|
|
超氧化物歧化酶(SOD)是需氧生物中普遍存在的一种含金属的酶。它在防御活性氧或其它过氧化物自由基对细胞膜系统的伤害, 防止细胞的衰老过程中起着重要的作用。
米槠离体胚LN2保存后SOD活性及其50%光还原需酶量见图 3。

|
图 3 LN2保存后米槠离体胚SOD活力分析 Fig. 3 SOD activity of C.carlesii excised embryos after cryopreservation
酶活性Enzyme activity  MF -MT; MF -MT;  MF -QT; MF -QT;  QF -QT; QF -QT;  MF-ST. MF-ST. 需酶量Needed enzyme volume —○— MF -MT; —●—MF -QT; —△—QF -QT; —▲—MF-ST. |
SOD酶活性随含水量降低是成抛物线形变化规律, 12%MC米槠离体胚经MF-QT其酶活性最大, 达244 U·h-1, 其次是15%MC, MF-ST方式, 为215 U·h-1 (图 3)。
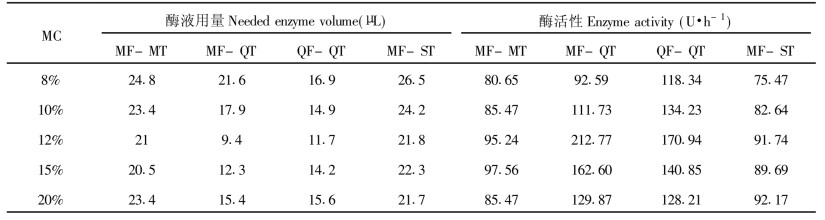
采用QF-QT处理后的苦槠离体胚SOD活性均比其他3种处理方法高。说明苦槠离体胚采用MF-QT法处理对细胞膜的伤害较轻, 即胚的抗逆性和抗衰老能力较强, 活力较高。随含水量的变化, 酶活性的变化趋势呈抛物线形, 峰值位于12%MC与MF-QT交叉点上, 达212.77每10个胚酶活性单位·h-1。不同的冷解冻方式之间酶活性差异也较大, QF-QT方式酶活性最大为170.94每10个胚酶活性单位·h-1 (表 4)。
|
|
米槠种子的MDA含量也是随含水量的降低而呈抛物线变化趋势, 米槠种子当含水量大于10%时, 其MDA含量是比较接近, 也比较低的, 最小的是12%MC, 其MF-QT方式处理的MDA含量仅为0.194 μmol·mL-1, 其他冷解冻处理的MDA含量也为最低。当米槠种子含水量小于10%时, 其膜脂过氧化产物MDA含量迅速增加, 已经对细胞造成极大膜损伤。而且, 不同冷冻解冻处理之间也存在较大的差异, 其中以MF-QT法MDA含量最低, 膜脂过氧化程度最轻(图 4a)。

|
图 4 LN2保存后米槠(a)和苦槠(b)离体胚MDA含量变化 Fig. 4 MDA content of excised embryos of C.carlesii (a) and C.kawakamii (b)after cryopreservation
 MF -MT; MF -MT;  MF-QT; MF-QT;  QF -QT; QF -QT;  MF-ST. MF-ST.
|
用较高的含水量进行超低温保存苦槠种子, 易造成低温伤害, 其最佳的含水量为12%MC, 经MF-QT后MDA仅为2.84 μmol·mL-1, MDA含量的变化趋势基本上呈倒抛物线形。但经过适当的处理后, 其超低温保存的成功含水量有可能会降低到8%或者更低(图 4b)。
米槠和苦槠2树种种子的MDA含量均随含水量的下降先下降后升高, 其最小值均出现在12%MC, MF-QT处理中。MDA含量越低, 说明对细胞膜的破坏就越小, 即种子的膜完整性保持地越好, 种子受到的伤害也越小, LN2保存更容易取得成功。因此, 由MDA测定结果表明:米槠和苦槠均是12%MC, MF-QT法。
3 讨论与分析LN2保存是当今迅猛发展起来的一种种质保存新技术, 有着广泛应用前景, 在超低温状态下, 细胞代谢基本上处于静止状态, 这将给保存材料遗传稳定性的长期维持开辟出一条引人入胜的崭新的途径。
新鲜的实验材料有着较好的质量特性, 米槠和苦槠现两树种发芽率分别高达79%和78%, 经LN2处理后, 其发芽率分别为50%和48%。引起这两树种发芽率降低的原因主要是实验材料失水过程中, 由于其不耐脱水而引起的。
细胞结构的完整性是种子活力的基础, 种子老化劣变很大程度上是由于细胞膜受到损伤甚至解体, 膜透性增大, 电解质外渗, 水浸液电导率上升, 不同活力的种子, 其膜修复能力不同(彭幼芬等, 1994; 张保恩等, 1999)。SOD、能够在逆境胁迫和衰老过程中清除种子体内过量的活性氧, 维持活性氧的代谢平衡、保护膜结构, 从而使顽拗性种子在一定程度上忍耐、抵抗逆境胁迫, 或延缓衰老进程(Norman E et al., 1989)。MDA含量高低则说明脂质过氧化的程度和对逆境条件反应的强弱。顽拗性种子在低温或脱水过程中易造成活性氧代谢系统的失调。对LN2保存后米槠和苦槠种子的电导率、SOD活性和MDA含量研究发现, 主要影响因素是含水量, 其次是不同的冷冻处理方式和解冻处理方式。最优的并非为最高或是最低含水量, 而是存在有最适含水区间, 过高的含水量则会在冷冻和解冻过程均会造成低温伤害, 而过低的含水量则会造成脱水伤害。适宜的含水量能够较好的维持细胞膜的完整性。研究结果还表明, LN2保存过程中, 米槠种子和离体胚最适含水量均为12%, 其发芽率最高, 细胞膜伤害最小, SOD活性最高, MDA含量最低, 细胞膜的完整性能够得到较好的保持; 而苦槠种子最适含水量为15%, 其发芽率最高, 其离体胚保存时, 其含水量可适当降低到12%, 此时细胞膜伤害最小, SOD活性最高, MDA含量最低。
国际种子检验协会(ISTA)编, 浙江大学种子科学中心译.1996国际种子检验规程.北京: 中国农业出版社, 1999, 15~ 18
|
黄学林, 陈润政. 1990. 种子生理实验手册. 北京: 农业出版社.
|
陆旺金, 傅家瑞. 1998. 顽拗性种子种质保存现状. 种子, (2): 37-40. |
彭幼芬, 王文章. 1994. 种子生理学. 湖南: 中南工业大学出版社, 158-183.
|
孙时轩. 1995. 造林学. 北京: 中国林业出版社, 63-64.
|
王贵禧, 梁丽松, 宗亦臣. 1999. 贮藏板栗休眠与萌芽的温度的调控. 林业科学, 35(3): 29-33. DOI:10.3321/j.issn:1001-7488.1999.03.006 |
王君晖, 黄纯农. 1998. 木本植物种质超低温保存的研究进展. 世界林业研究, (5): 6-11. |
张保恩, 黄学林. 1999. 种子吸胀期间的泄漏物与活力的关系. 植物生理学通讯, 35(3): 231-235. |
朱广谦, 钟诲文, 张爱琴. 1990. 植物生理学实验. 北京: 北京大学出版社.
|
Farrant J M, Pammenter N W, Berjak P. 1988. Recalcitrant -a current assessment. Seeds Sci.&Techno., (16): 155-166. |
Norman E, et al. 1989. Assay and electrophoresis of superoxide dismutase from red spruce (Picea rubens Sarg.), loblolly pine (Pinus teada L.), and sotch pine(Pinus sylrestris L.). Plant Physiol, 90: 742. DOI:10.1104/pp.90.2.742 |
Rorberts E H. 1973. Predicting the storage life of the seeds. Seeds Science and Technology.Ⅰ.: 499-514. |