 2001, Vol. 37
2001, Vol. 37文章信息
- 洪伟, 柳江, 吴承祯.
- Hong Wei, Liu Jiang, Wu Chengzhen.
- 红锥种群结构和空间分布格局的研究
- A STUDY ON STRUCTURE AND SPATIAL DISTRIBUTION PATTERN OF CASTANOPSIS HYSTRIX POPULATION
- 林业科学, 2001, 37(专刊1): 6-10.
- Scientia Silvae Sinicae, 2001, 37(专刊1): 6-10.
-
文章历史
- 收稿日期:2001-01-08
-
作者相关文章


 、CA 5种聚焦度指标及Iwao
、CA 5种聚焦度指标及Iwao  回归测定种群空间分布格局和动态。结果表明:红锥种群结构存在增长和下降两种类型;聚焦度指标测定结果一致表明红锥种群的空间分布为随机型:Iwao 回归分析表明分布的基本成分是红锥个体;在种群成长过程中,分布格局从聚集型向随机型转化。 regression suggested that the basic component of distribution was individual. Population dynamics analysis showed the spatial distribution pattern changed from aggregate type to random type during the course of population growth.
回归测定种群空间分布格局和动态。结果表明:红锥种群结构存在增长和下降两种类型;聚焦度指标测定结果一致表明红锥种群的空间分布为随机型:Iwao 回归分析表明分布的基本成分是红锥个体;在种群成长过程中,分布格局从聚集型向随机型转化。 regression suggested that the basic component of distribution was individual. Population dynamics analysis showed the spatial distribution pattern changed from aggregate type to random type during the course of population growth.红锥(Castanopsis hystrix)又名刺栲、红栲,是福建省南亚热带优良阔叶树种之一,具有生长迅速,材质优良等特点,是家具和造船的重要用材,木材价格昂贵。红锥喜温暖湿润的气候,耐阴,多生于年降水量在1100~2000 mm,年均温在18℃~24℃的地区,主要分布在福建华安、南靖、平和、松溪、龙岩和漳平等地。目前,红锥资源日趋减少,亟待采取有效的保护和经营措施,然而有关红锥的研究鲜有报道。因此,本研究试图揭示红锥种群的结构及分布格局等种群学特征,为红锥的保护和合理经营提供理论依据。
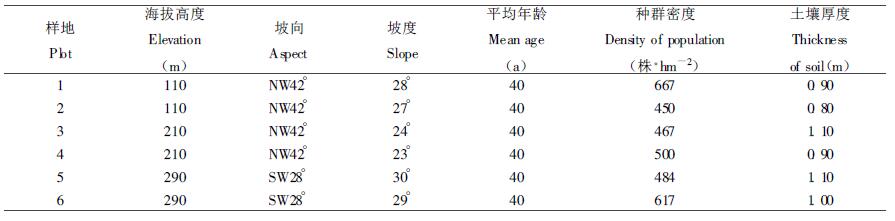
1 研究区自然概况研究区位于福建省华安县中部的刀石山,地处华东与华南区的分界线上,其地理坐标为北纬24°38′~25°11′、东经117°17′~117°40′,属戴云山南端与博平岭东端的延支脉,海拔90~390 m。该区属东南亚热带季风气候,温暖少严寒,年平均气温为22℃,极端最高温为39℃,极端最低温为-5℃,年平均降雨量为1770 mm,无霜期330 d,年平均相对湿度在77%~81%之间,土壤主要类型为红壤,亚热带植被发育旺盛,原生植被为亚热带常绿阔叶林。红锥种群分布广泛,密度约600株·hm-2,主要伴生树种有木荷(Schima superba)、闽粤栲(Castanopsis fissa)、山杜英(Elaeocarpus sylvestris)、米槠(Castanopsis carlesii)、冬青(Ilex purpurea)等,灌草及藤本有狗脊(Woodwardia japonica)、芒萁(Dicranopteris dicotoma)、毛鳞省藤(Calamus thysandepsis)、扁担藤(Tetrastigma planicaule)等。
2 研究方法 2.1 材料收集在刀石山红锥种群典型分布地段,设置6块20 m×30 m的样地,测定每块样地的海拔、坡向、坡度、土壤厚度和群落类型等立地因子,如表 1。在各样地内分别采用相邻格子法布置10 m×10 m的小样方,共计36个,对每个小样方进行每木检尺,记录样方内红锥的胸径、树高、冠幅、枝下高(起测每径阶≥4 cm)、盖度,以及胸径4 cm以下的幼苗、幼树的株数。
|
|
每一个体大小的确定,根据研究经验(Rundel,1971),采用胸径或树高作为个体大小的指标,具有良好的一致性。本文选取胸径代表个体大小来分析红锥种群大小结构,根据种群生活史和研究目的,红锥种群大小结构划分按以下方法处理:胸径小于2.5 cm为1级;胸径2.5~10 cm为2级;胸径大于10 cm的按每增加10 cm,大小级增加1级。红锥种群等级度结构按曲仲湘的树木等级度划分方法(曲仲湘等,1985),对6块样地的红锥个体逐一确定其等级度。
2.3 红锥种群分布格局类型和动态的研究方法测定种群空间分布格局的方法很多(丁岩钦,1980;洪伟等,1987;1992;2000),本文主要采用聚集度指标进行测定。聚集度指标是度量一个种群空间分布的聚集程度(随机、均匀或聚集),它克服了频次比较法出现种群同时属于多种分布的混乱矛盾的解释状态(洪伟,2000)。按如下指标进行计算:
2.3.1 扩散系数C (丁岩钦,1980)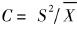
Iδ=n (∑xi2fi-N)/[N (N-1)],当Iδ=1时为随机分布;Iδ < 1时为均匀分布;Iδ > 1时为聚集分布。指数的随机分布偏离度采用F检验法进行显著性检验。
2.3.3 平均拥挤度M* (丁岩钦,1980)M*=∑xi2/∑xi-1,当M*>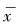
当
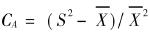
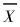
回归方程

|
种群动态依据空间差异代替时间变化的理论,以一次性调查结果来分析空间分布格局的变化,即用不同大小级种群的分布格局变化来揭示种群发育过程中分布格局的动态变化。
3 结果和分析 3.1 种群结构将6块样地中红锥种群的大小级加以整理,以1个径级龄为1个龄级,将各龄级现存个体数标准化后用径级龄比表示,结果如图 1。在种群结构图中,横坐标表示各个龄级的个体数所占的比例,纵坐标表示各个大小级。从红锥种群大小级结构分析,红锥种群可以分为2种类型:(1)增长型种群 种群存在较多的幼苗和幼树,大小结构大致呈金字塔型,如样地1、3、5。(2)下降型种群 种群大小级结构虽然还存在连续性,但分布已不规则,中树和大树较多,幼苗数量很少,种群更新能力减弱,呈下降趋势,如样地2、4、6。同一发育阶段的红锥种群存在两种不同类型的种群结构,分析其产生的可能原因,主要是由于种群的生物学特性和生境条件及其相互作用决定的。在增长型种群的样地中,大树数量不多,中、幼树有良好发育,且天然更新旺盛;在下降型种群的样地中,大、中树数量较多,并且存在大小不一的林窗,而且其他阔叶树种,如木荷、闽粤栲等有较大程度的入侵,因此,种间竞争更为激烈,阻碍了红锥幼苗和幼树的生长,天然更新受到限制。但更为主要的原因可能还是由于生境条件的差异,在整个调查区明显存在斑块性,斑块间差异明显,斑块内条件相对一致,下降型种群的样地土质明显劣于增长型种群的样地,土粒粗大,沙石含量高,土层厚度也相对较薄,因此,不利于红锥种群的生长发育,随着发育进程的延续,一些对生境适应能力更强的树种入侵并定居下来,加剧了种间竞争,使得红锥种群结构的变化速率加快,表现出衰退趋势。另外,由于红锥自身的生物学特性,在5a以前生长缓慢,以后生长迅速,种内竞争比较激烈,强烈的自疏作用也为其他树种侵入提供了机会,并阻碍了红锥幼苗的发育,种群更新受到限制。

|
图 1 各样地红锥种群大小结构 Fig. 1 The size structure of Castanopsis hystrix in six plots |
进一步利用曲仲湘的树木等级度划分方法对6块样地的红锥种群个体逐一确定其等级度,以等级度为横坐标,各等级度所包含的个体数作总和标准化后的相对数量为纵坐标绘制等级度结构图(蔡飞,1997; 于明坚,1999)。参考钟章成(1988)将常绿阔叶林植物种群等级度结构划分为初生正常型、旺盛型、成熟型、始衰型、中衰型和老衰型6种类型的标准,6个样地中样地1、5中的红锥种群属于旺盛型,样地3中的种群属于成熟型,样地2、6中的红锥种群林属于始衰型,而样地4中的红锥种群属于中衰型,与采用径级龄比表示种群结构图结果相一致。
3.2 种群分布格局根据6块样地资料,分别测定上述聚集度指标(表 2)。由表中结果可以看出,C值均近似为1,经χ2检验,均小于查表值χ0.052;Iδ值均近似为1,经随机偏离度显著性检验F0值均小于F0.05;M*均近似等于
|
|
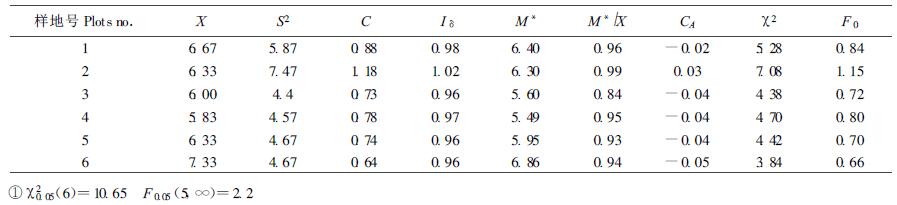
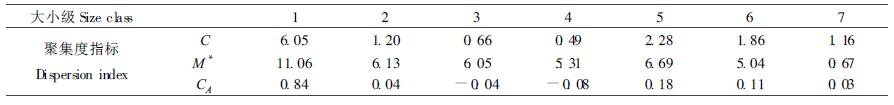
采用C、M*、CA 3种指标,进一步对种群各大小级的空间分布格局进行分析(表 3)。C值经χ2及蒙特卡洛法检验,1、5、6级属聚集型,其他各级属随机型;M*值测定表明1、5级属聚集型,其他各级属随机型,并且反映出种群从幼苗幼树(11.06)到成熟个体(0.67)具有一定的扩散性;CA值与C值具有相同的判断结果。可以看出,红锥种群从幼苗幼树到成熟个体,其空间分布格局的变化的总体趋势是由聚集型转化为随机型。因为随着红锥种群的发育、个体的年龄与体积的增大,它们对环境条件的要求提高,受环境条件的限制,种群会发生自疏与它疏,从而导致种群由聚集分布向随机分布过渡。所以红锥种群分布格局分布动态总体趋势表现为从幼苗—小树—中树—大树,种群的分布格局由聚集逐渐向随机分布过渡。
|
|
分析这种空间分布格局的形成原因,主要是由红锥的生物学特性及与其他树种相互作用的结果所决定的。红锥林分起源于“根出条”的无性繁殖,5a以前生长缓慢,因此,在幼年阶段生长地域较为集中,并且由于母树生境差异导致分布不均,表现为明显的聚集分布;5a以后生长迅速,种内竞争开始加剧,发生自疏现象,分化作用强烈,部分植株死亡,个体在争夺营养空间和生态位重新分配的过程中使得种群的聚集程度逐渐降低,到个体成熟时,聚块已经完全解体,种群的分布格局也由聚集型转化为随机型,成熟个体是原来同一聚块中竞争的优胜者,占有上层的空间生态位,并在资源利用上具有优势;在中树向大树的转化过程中,又出现了轻微的聚集的现象,但存在的时间并不太长,这可能是种群从中树向大树转化的过程中的过渡现象。在种群从幼树向中树转化的过程中,扩散作用强烈,激烈的种内竞争也为其他树种的入侵提供了条件,从而又使种间竞争开始逐渐加强,种群的分布格局有所改变,但竞争的结果并未改变扩散的趋势,成熟的红锥个体最终占据着主林层,并呈现出随机分布状态。
从上述的分析结果可见,各指数的计算结果存在一定的波动,尤其在计算红锥种群各大小级6级林木的空间分布格局聚集指数时,扩散系数C与其它指数的值出现矛盾的结果,但经聚集度各指数相互验证,6级林木的分布格局为随机分布,这也与实际情况相符。而其它指标测定结果一致,因此,扩散系数Iδ、平均拥挤度M*、CA、Iwao回归等指标能较好地反映红锥种群的分布格局。由于各聚集度指标构造方法不一样,所以有时会出现各测定指标测定结果不一致的现象。故建议最好应用两种以上的检验方法,当两种以上方法给出一致性的结果时,则可以认为已反映一定的真实性。
4 讨论用胸径代表个体大小来研究福建华安刀石山红锥种群的结构特征,大小级和等级度的分析结果一致,表明红锥种群存在增长和下降两种类型。产生这种现象的主要原因可能在于生境的差异,反映出生境差异对种群结构的变化速率有显著影响,导致不同的发育类型。
种群分布格局是物种与环境长期相互适应、相互作用的结果,不仅与物种的生物学特性和种群间的竞争排斥有关,而且与物种的生境有密切的联系。聚集度指标对红锥种群空间分布格局的测定结果具有良好的一致性,红锥种群整体呈随机分布,这是由红锥的生物生态学特性所决定的。从种群的生物学特性和分布格局的动态来看,种群具有一定的扩散性,幼苗幼树呈聚集分布,中、大树呈随机分布。这种变化趋势是种内和种间竞争以及种群与生境相互作用的共同结果,反映了种群的一种适应机制(洪伟等,1990; 吴承祯等,2000b)。红锥种群为优势种,处于乔木主林层,深根性,侧根发达。萌芽力极强,每个伐根长出1~8株萌条。即使没有采伐,也可由树干基部的“根出条”长成大、中径级林木。这种“根出条”的繁衍演替方式,使红锥始终维持着多维空间的生态位。在天然林中,其既可实生繁殖,也可无性繁殖更新,因此,在幼年阶段表现为明显的聚集分布;随着种群个体的增大,种内与种间竞争加剧,自疏与它疏共存,红锥种群个体数量减少,从而形成随机分布。
红锥种群结构和空间分布格局的研究结果对种群的保护、合理经营都具有重要的理论和现实意义,同时也为阔叶林生态系统结构和功能的进一步研究提供理论依据。因此,在红锥林经营过程中,应遵循其种群结构与空间格局分布规律,通过人为的干扰如抚育间伐等,促使群落物种在水平结构和垂直结构上达到合理配置,以充分利用红锥林物种种内种间竞争规律,提高林分生产力。由于在天然林中红锥竞争性极强,在次生林中红锥占绝对优势,伴生树种一般只居于第2层,因此,在营造人工林时可营造纯林,也可与杉木、马尾松等针叶树种混交。
蔡飞, 宋永昌. 1997. 武夷山木荷种群结构和动态研究. 植物生态学报, 21(2): 138-148. DOI:10.3321/j.issn:1005-264X.1997.02.005 |
丁岩钦. 1980. 昆虫种群数学生态学原理与方法. 北京: 科学出版社, 84-124.
|
洪伟, 吴承祯. 1999. 马尾松人工林经营模式及其应用. 北京: 中国林业出版社, 110-129.
|
洪伟主编. 2000. 闽江流域森林生态研究. 厦门: 厦门大学出版社, 176-183.
|
洪伟, 罗顺跃, 陈顺立, 等. 1992. 油茶主要病虫害生态空间分布规律的研究. 应用生态学报, 3(4): 308-312. DOI:10.3321/j.issn:1001-9332.1992.04.009 |
洪伟. 1989. Iwao M*-M模型随机分析偏离度检验方法的研究. 植物保护学报, 16(2): 107-112. DOI:10.3321/j.issn:0577-7518.1989.02.009 |
洪伟, 吴承祯. 1997. Taylor幂法则模型的遗传算法最优拟合. 福建林学院学报, 17(1): 1-5. |
洪伟, 郑康宏, 龚其锦. 1990. 毛竹在杉木与毛竹混交林中的空间分布型的研究. 竹子研究汇刊, 9(2). |
曲仲湘, 吴玉树, 王焕校, 等. 1985. 植物生态学. 北京: 高等教育出版社, 152-170.
|
吴承祯, 洪伟, 吴继林, 等. 2000a. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 11(3): 333-336. |
吴承祯, 洪伟, 吴继林, 等. 2000b. 珍稀濒危植物长苞铁杉的分布格局. 植物资源与环境学报, 9(1): 31-34. |
于明坚. 1999. 青冈常绿阔叶林群落动态研究. 林业科学, 15(6): 42-57. DOI:10.3321/j.issn:1001-7488.1999.06.006 |
钟章成. 1988. 常绿阔叶林生态学研究. 重庆: 西南师范大学出版社, 281-296.
|