 2001, Vol. 37
2001, Vol. 37文章信息
- 李丽霞, 梁宗锁, 韩蕊莲.
- Li Lixia, Liang Zongsuo, Han Ruilian.
- 干旱对沙棘休眠、萌芽期内源激素及萌芽特性的影响
- EFFECT OF SOIL DROUGHT ON ENDOGENOUS HORMONE AND SPROUT CHARACTER IN DORMANCY AND SPROUT PERIOD OF SEABUCKTHORN SEEDLINGS
- 林业科学, 2001, 37(5): 35-40.
- Scientia Silvae Sinicae, 2001, 37(5): 35-40.
-
文章历史
- 收稿日期:2000-08-08
-
作者相关文章


在黄土高原地区, 冬春连旱是限制造林成活的主要因素, 土壤水分不足也是树木萌芽后生长发育的重要限制因子, 对生长季节水分亏缺下树木水分生理与生长已有较多报道(夏新莉, 2000; 王邦锡, 1997; 董学军, 1999; 王俊峰, 1999), 抗旱特性及机制也已在有关综述中得到反映(胡新生, 1998), 而冬季休眠和春季萌芽期土壤缺水对其影响的关系研究较少。栽植后第1a沙棘幼苗休眠与第2 a春季萌发是一个复杂的生理过程, 若土壤中缺水, 则其休眠与萌发情况更为复杂, 这是一个水分与激素不断平衡与失衡的动态过程。苗体水分变化控制五大类内源激素水平的变化, 已有文献表明在冬季促进植株休眠的内源激素主要是ABA, 而春季打破休眠促进萌发的激素为GA1/3, 2类激素协调作用, 调控植株的萌芽情况。春季, 温度回升, 日照延长, 二者共同的合成途径一甲瓦龙酸途径倾向于合成GA1/3, 故GA1/3/ABA呈上升趋势, 促发幼苗的萌芽过程; 但如果此时水分不足, 导致植株含水量下降, 则ABA合成加强, GA1/3 ABA趋于下降, 萌芽受抑, 随时间推延, 温度升高, 则死亡机率增大。在黄土高原地区造林后若在雨水充足的年份幼苗易成活, 在完成第1个生长季后的冬季休眠期和第2 a春季萌芽期遇到干旱仍然会大量死亡或生长不良。为揭示这一生理机制, 了解沙棘休眠与萌发生理过程而进行本研究, 探讨了土壤干旱条件下沙棘幼苗栽植成活后冬季休眠期与第2 a春季萌芽期苗体水分关系, GA1/3/ABA比值与萌芽率的相关关系, 为沙棘抗旱造林与土壤缺水下幼林抚育管理和促进萌芽生长提供理论依据。
1 材料与方法 1.1 试验材料与处理方法选用中国沙棘2 a实生苗。1999-04初选大小基本一致, 生长健壮, 根系良好的苗木栽入高32 cm, 内径30 cm的白瓷盆内, 每盆3株, 土壤最大持水量30.1 %; 每盆装土12500 cm3, 放置于西北农林科技大学(西农校区)农业水土工程研究所玻璃遮雨棚下, 使其正常生长。于同年12月筛选出地径1 cm左右, 株高30 ~ 50 cm, 生长一致的18盆植株, 分为3组; 设置3种土壤含水量处理, 分别为21 %、15 %及9 %, 相当于田间持水量70 %、50 %及30 %, 待土壤水分自然干燥至设计要求后开始取样测定, 用TDR(时域反射仪)监测土壤含水量, 适时补水, 使其土壤含水量稳定在设计要求。随机取样, 3次重复, 休眠期约10 d测定1次, 萌芽期3 ~ 4 d取样测定1次, 整个试验从1999-12-10至2000-03-29共持续110 d。
1.2 测定项目及方法 1.2.1 沙棘苗体水分状况研究沙棘苗体含水量:烘干称重法; 沙棘苗体自由水束缚水:取用沙棘韧皮部, 马林契诃夫法测定; 水势测定用美国产3005型压力室测定(高俊凤, 1999)。
1.2.2 内源激素含量测定随机取样、休眠期取韧皮部, 约半月1次, 萌芽关键期取芽并加强取样, 平均每周1次, 每次取样时用剪刀截取侧枝上梢10 cm左右, 刀片刮取韧皮部或剥取芽, 迅速称重, -40 ℃冰冻保存用于内源ABA和GA1/3测定, 测定方法为酶联免疫法, 试剂盒由南京农业大学植物激素室提供。
1.2.3 沙棘苗体幼芽长、单枝叶片数刻度尺准确测量芽长; 生长幼枝叶片数统计选定植株、枝条挂牌标记, 连续观测。
2 结果分析 2.1 缺水条件下沙棘苗木水势的变化表 1表明, 随土壤干旱加剧和时间延长, 沙棘苗体水势呈逐步下降趋势, 与适度水分的对照相比, 重度干旱使苗木水势最低达-2.89 MPa, 降低23.38 %; 中度干旱使苗木水势低至-2.67 MPa, 降低14.10 %; 干旱处理的苗木水势均在胁迫40 d水势降至最低值, 此后随气温回升而缓慢上升。
|
|
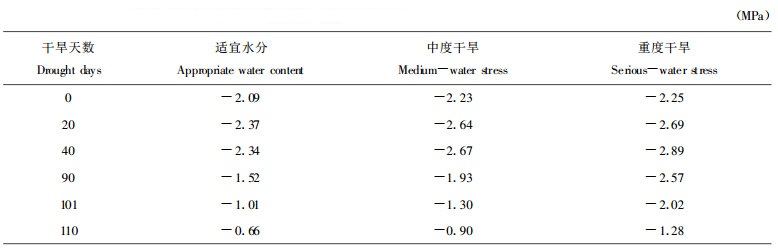
图 1表明土壤干旱使沙棘苗体含水量在休眠期一直呈现较快下降的趋势, 重度干旱下苗木含水量下降更多。气温回升后, 苗木根系吸水加强, 对照苗木的含水量呈现稳定的上升趋势, 中度干旱下苗木含水量上升较缓, 重度干旱处理的苗体含水量不能恢复到初期水平, 一直维持在较低的范围内, 且在胁迫104 d又出现第2次低值。

|
图 1 不同土壤水分下沙棘苗体含水量变化 Fig. 1 Changes of water content in seabuckthorn seedlings under different soil water content
 适宜水分Control; 适宜水分Control;  中度干旱Medium water stress; 中度干旱Medium water stress;
 重度干旱Serious water stress. 重度干旱Serious water stress.
|
由图 2可以看出, 中度干旱使沙棘苗体自由水含量下降, 始终低于对照水平, 重度干旱下苗木自由水下降更多, 其最低值与对照相比下降9.32 %。总趋势与苗体含水量类似, 具备休眠期下降, 直至最低值, 萌芽期逐步回升这一共同变化特征。

|
图 2 不同土壤水分下沙棘苗体自由水变化 Fig. 2 Changes of free water in seabuckthorn under different soil water content
适宜水分Control; 中度干旱Medium water stress;
重度干旱Serious water stress.
|
图 3显示3种处理间苗木束缚水自由水差异不明显, 总体趋势为从休眠期的较高值向萌芽期较低值的变化。

|
图 3 同土壤水分下沙棘苗体束自变化 Fig. 3 Binging water free water of seakbuckthorn under different soil water content
适宜水分Control; 中度干旱Medium water stress;
重度干旱Serious water stress.
|
图 4(A)表明沙棘在休眠期其韧皮部内ABA含量较低, 约在3000 pmol/g·FW范围之内, 总趋势为重度干旱下苗木ABA含量>中度干旱处理>适宜水分处理。与韧皮部含量相比, 沙棘芽中ABA含量差异较大, 分布范围为527.15 ~ 13707.83 pmol/g·FW, 中度干旱下苗木内源ABA含量稍高于适宜水分处理, 而重度干旱使苗木内源ABA水平显著高于适宜水分处理。胁迫104 d时重度干旱苗木内源ABA含量出现一峰值(见图 4(B)), 分析认为胁迫101 ~ 107 d外界持续高温, 使苗木加速失水, 苗木含水量持续下降, 伴随含水量降低, ABA骤增, 后随气温逐步下降, 苗木失水减少, 含水量上升, 则ABA快速下降, 内源ABA的这一变化十分快速灵敏, 这也是ABA作为对干旱逆境响应的根源信号所具备的反应特性之一。适宜水分与中度干旱下苗木无含水量下降及ABA骤增这一过程的发生。

|
图 4(A) 不同土壤水分下沙棘苗体韧皮部ABA含量变化 Fig. 4(A) Changes of ABA content in seabuckthorn seedlings under different soil water content |

|
图 4(B) 不同土壤水分下沙棘苗体芽中ABA含量变化 Fig. 4(B) Changes of ABA content of shoots in seabuckthorn seedlings under different soil water content
适宜水分Control; 中度干旱Medium water stress;
重度干旱Serious water stress.
|
从图 5(A)中可以看出, 休眠期沙棘韧皮部中GA1/3水平处于553.13 ~ 1755.07 pmol g·FW之间, 总体表现为适宜水分处理下苗木内源GA1/3含量>中度干旱处理>重度干旱处理, 但对照与土壤干旱的苗木内源GA1/3含量差异不明显。

|
图 5(A) 不同土壤水分下沙棘苗体韧皮部GA1/3含量变化 Fig. 5(A) Changes of GA1/3 content of phleom in seabuckthorn seedlings under different soil water content |
如图 5(B)所示到萌芽关键期时, 处理之间GA1/3含量差异趋于显著, 适宜水分条件下苗木萌发的芽中GA1/3含量有大幅提高, 而中度干旱下苗木随休眠的逐步解除, GA1/3呈较快上升的趋势, 重度干旱处理的苗木芽中内源GA1/3上升非常缓慢。

|
图 5(B) 不同土壤水分下沙棘苗体芽中GA1/3含量变化 Fig. 5(B) Changes of GA1/3 content of shoots in seabuckthorn seedlings under different soil water content
适宜水分Control; 中度干旱Medium water stress;
重度干旱Serious water stress.
|
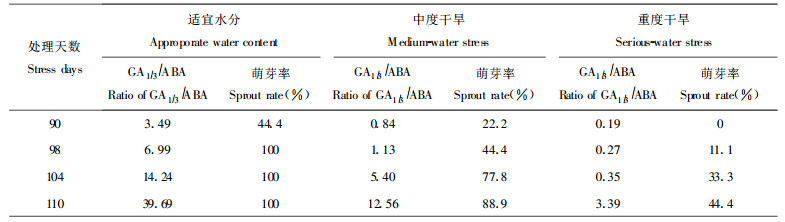
由ABA与GA1/3的测定结果计算出GA1/3/ABA比值。图 6(A)表明, 沙棘休眠期韧皮部GA1/3与ABA含量相近, 比值范围为0.228 ~ 1.545之间。中度干旱使沙棘苗木GA1/3/ABA比值大致低于对照, 重度干旱下苗木该比值下降非常明显。图 6(B)所示, 到萌芽关键期时, 随苗体含水量的增加, 内源GA1/3表现增加, ABA下降, 故适宜水分处理的苗木GA1/3/ABA平稳上升, 重度干旱使沙棘苗体内较长时间维持高水平ABA, 故GA1/3/ABA上升较缓。若将GA1/3/ABA比值与萌芽率统计结果联系起来分析, 如表 2所示, 可以看出, 适宜水分处理的苗木GA1/3 ABA提高早、快, 表现萌芽早, 在较短时间内即全部萌芽成活; 中度干旱延迟了苗木的萌芽期, 若以萌芽率>50 %为时间界限, 则达到该日期较适宜水分处理苗木推迟10 d左右。重度胁迫使苗木达到有效萌芽率的日期更迟, GA1/3/ABA上升的幅度也较小。

|
图 6(A) 不同土壤水分下沙棘苗体韧皮部GA1/3/ABA含量变化 Fig. 6(A) Changes of GA1/3/ABA of phleom in seabuckthorn seedlings under different soil water content |

|
图 6(B) 不同土壤水分下沙棘苗体芽中GA1/3/ABA含量变化 Fig. 6(B) Changes of GA1/3/ABA of shoots in seabuckthorn seedlings under different soil water content |
|
|
经长时间不同程度干旱胁迫后使幼苗生长受到不同程度的抑制, 与对照相比, 萌芽日期推迟, 生长减慢。中度干旱下苗木单枝平均叶片数为5.67个, 幼芽平均伸长量为1.08 cm, 重度干旱下苗木单枝平均叶片度仅为2.10个, 幼芽平均伸长量仅为0.47 cm, 分别为对照处理的23.84 %及25.27 %(图 7)。
|
|
图 7 不同土壤水分下沙棘苗体短枝平均芽长及芽数变化 Fig. 7 Changes of average shoots length and shoots number of short branches in seakbuckthorn seedlings under different soil water content
 芽数Shoots number; 芽数Shoots number;  芽长Shoots length. 芽长Shoots length.
|
外界环境因子例如光照、温度、水分等对植物生长发育的调控主要是通过改变体内各种激素的平衡来实现的, 并且这种调控具备明显的时间效应。随季节和生育期变化, 生长促进类激素与生长抑制类激素发生有规律消长, 随着新的平衡的建立, 植物进入新的特定的发育阶段。因此, 季节变换及生育期转化阶段的内源激素变化更值得重视。在草本植物特别是农作物上内源激素研究较集中(Gropt, 1992; 王宏, 1997;), 以木本植物为试材的研究中, 又以激素与生根关系研究较多(郑均宝, 1991; 张志翔, 1994)而苗木在土壤缺水条件下冬季休眠与春季萌芽与内源激素的互作研究报道较少。本文研究了中国沙棘休眠期及萌芽期内源GA1/3及ABA连续变化动态, 并确立了枝条达有效萌芽率(>50 %)时GA1/3 ABA比值应不小于5, 为探讨栽植后经1年期生长后在冬季休眠和第2 a春季萌芽期的生理变化机制积累了有价值资料。
在本研究中, 适宜水分处理的沙棘苗木已100 %萌芽成活后, 重度水分亏缺的苗木仍有2/3以上植株处于休眠状态, 说明重度干旱使其休眠程度较深, 在春季来临后, 不能及时降低ABA, 提高GA1/3, 故萌发延迟。如果外界维持较低温度, 无风天气则不致于引起苗体过度失水, 苗木可继续维持一段时间休眠状态, 在较长时间内调整内源激素水平。若遭遇不适的外界环境(在我国北方, 特别是在黄土高原地区这种情况经常发生), 随温度持续回升并伴有多风的春季则必然引起苗木加速失水, 而苗木吸水功能尚未完全恢复, 土壤中可利用水又十分有限, 最终必然造成苗木失水枯死。因此持续不断的土壤干旱对成活幼龄苗木危害很大。这与作者以前的研究结论相符(李丽霞等, 1999)。这一研究同时也证明在成活关键期, 缺水的外界环境影响较大, 能否萌芽及萌芽后能否成活与外界环境密切相关。沙棘苗萌芽后, 芽顶破原有茎表皮形成伤口, 温和的外部条件有利于苗木水分平衡的维持, 有利于成活; 不适水分条件必然使苗体过度失水, 而处于干旱土壤中的根系无法吸收足够的水补给地上部分, 则萌芽后不一定能够成活。
经本研究证明, 休眠与萌芽期土壤干旱使苗木含水量下降, 失去水分平衡, 内源ABA增加, GA1/3减少, GA1/3/ABA比值下降, 解除休眠时间延长故萌发延迟, 中度胁迫苗木延迟约10 d, 重度胁迫下苗木延迟约25 d; 沙棘作为一种抗旱性强的造林树种, 经历长期土壤中度干旱后, 并未表现明显受害及萌芽后生长受抑的症状, 因此, 在实际生产中, 保证土壤含水量在田间持水量50 %以上, 即可保证幼苗的萌芽与生长。我们试图采取外源GA3处理提高苗木萌芽率的措施业已取得初步成效。
董学军, 陈仲新, 阿拉腾宝, 等. 1999. 毛乌素沙地沙地柏(Sabina vulgaris)的水分生态初步研究. 植物生态学报, 23(4): 311-319. DOI:10.3321/j.issn:1005-264X.1999.04.003 |
高俊凤主编.植物生理学实验指导.西安: 世界图书出版公司, 1999
|
胡新生, 王世绩. 1998. 树木水分胁迫生理与耐旱性研究进展及展望. 林业科学, 34(2): 77-89. DOI:10.3321/j.issn:1001-7488.1998.02.011 |
李丽霞, 梁宗锁, 王俊峰. 1999. 土壤水分和风速对沙棘苗木水分状况和成活率影响的实验研究. 沙棘, 12(4): 18-21. |
王邦锡, 王辉, 黄久常. 1997. 沙拐枣同化枝的光合作用和呼吸作用对生长季节、光照强度、高温和干旱的响应. 林业科学, 33(1): 18-23. DOI:10.3321/j.issn:1001-7488.1997.01.003 |
王宏, 项时康, 陈建华. 1997. 棉花种子发芽过程中生理生化变化及赤霉素(GA 3)的调节. 棉花学报, 9(3): 126-131. |
王俊峰, 梁宗锁编著.沙棘生物学特性与利用.西安: 陕西科学技术出版社, 1999, 84~ 93
|
夏新莉, 郑彩霞, 尹伟伦. 2000. 土壤干旱胁迫对樟子松针叶膜脂过氧化、膜脂成分和乙烯释放的影响. 林业科学, 36(3): 8-12. DOI:10.3321/j.issn:1001-7488.2000.03.002 |
张志翔, 火树华. 1994. 中国沙棘休眠枝条中ABA和IAA含量消长对生根的影响. 林业科技通讯, 12: 36. |
郑均宝, 刘玉军, 裴宝华. 1991. 几种木本植物插穗生根与内源IAA, ABA关系. 植物生理学报, 17(3): 313-316. DOI:10.3321/j.issn:1671-3877.1991.03.005 |
Groot spc, Karssen CM. 1992. Dormancy and germination of abscisic acid-deficient tomato seeds.Studies with the sitiens mutant. Plant physiol, 3: 952-958. |