 2001, Vol. 37
2001, Vol. 37文章信息
- 张彦东, 白尚斌, 王政权, 沈有信.
- Zhang Yandong, Bai Shangbin, Wang Zhengquan, Shen Youxin.
- 混交条件下水曲柳落叶松根系的生长与分布
- EFFECTS OF THE MIXED ON ROOT GROWTH AND DISTRIBUTION OF FRAXINUS MANDSHURICA AND LARIX GMELINII
- 林业科学, 2001, 37(5): 16-23.
- Scientia Silvae Sinicae, 2001, 37(5): 16-23.
-
文章历史
- 收稿日期:2000-08-08
-
作者相关文章


2. 云南省林科院 昆明 650204;
3. 东北林业大学 哈尔滨 150040
2. Yunnan Academy of Forest Sciences Kunming 650204;
3. Northeast Forest University Harbin 150040
相邻植物个体间的竞争是普遍存在的现象, 一些研究表明, 在大多数生态系统中, 竞争主要发生在地下, 彼此相邻的植物主要是为土壤资源而竞争(田蒴宁, 1992; 格雷戈里, 1992)。在竞争过程中植物的根系相互作用, 在同种和异种组成的群落中, 由于竞争对象的差异, 植物的根系会产生不同的反应。有能力的竞争者可能将大量的碳化物投入到根系中, 以获得更多的养分和水分。被动适应者可能改变根系的分布格局, 以避开直接的养分争夺(格雷戈里, 1992)。异种相邻树木间根系的相互作用, 使双方的根系在形态、分布以及生长上发生一系列不同于纯林的变化(Mckay, 1988)。了解这些变化对于深入分析植物种间关系具有重要意义。然而, 到目前为止不同植物种间根系相互作用并未得到很好的研究(格雷戈里, 1992; Mckay, 1988)。
水曲柳(Fraxinus mandshurica)和落叶松(Larix gmelinii)是我国东北林区的重要用材树种, 水曲柳和落叶松组成的混交林是非常成功的混交类型, 在混交林中水曲柳始终表现出增产效应。对于水曲柳和落叶松混交增产的机制, 在种间营养关系等方面已进行了较多的研究(张彦东, 1997)。但关于地下根系相互作用对种间关系影响问题的探讨还较少。国庆喜(1991)等在野外对26 a生的水曲柳落叶松带状混交林的根密度进行过比较, 王政权等(2000)在室内用盆栽的方法研究了水曲柳落叶松幼苗根系间的相互作用。本研究采用野外取样和室内栽培相结合的方法, 对混交条件下的水曲柳落叶松根系进行比较研究, 为进一步探讨水曲柳落叶松混交增产的机制提供依据。
1 研究方法 1.1 野外根系调查野外根系调查于1997-07上旬在东北林业大学帽儿山实验林场尖砬沟森林培育实验站进行。该站位于东经127°43′, 北纬45°16′。属中温带湿润气候, 地貌类型为低山丘陵, 土壤为暗棕壤。调查的水曲柳和落叶松混交林及相应的纯林均为1987年营造, 调查时林龄为10 a。3种林分初植时的株行距均为1.5 m×1.5 m, 调查时尚未进行间伐。混交林采用带状混交, 水曲柳3行为1条带, 落叶松5行1条带。纯林和混交林同处于东坡的中下部, 土壤是典型暗棕壤, 林分状况如表 1。
|
|
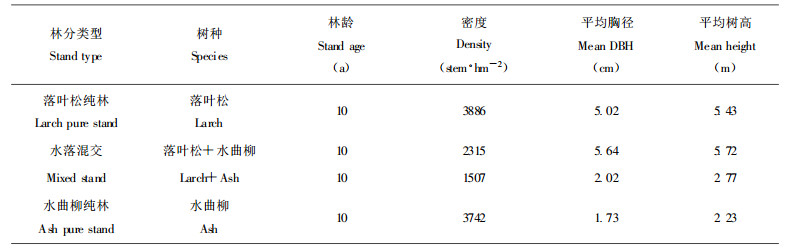
采用土钻法, 土钻的内径为6.5 cm。在混交林中设置4块样地, 每块样地宽6 m(左侧以水曲柳带的中间行为边界右侧以落叶松带的中间行为边界), 长30 m。在每块样地内的每两行树木中间每隔4 m设置1个采样点(每行8个样点, 每块样地共32个样点), 分0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm 3个层次钻取土壤中的根系(由于幼林根系集中分布于表层, 而且土钻法较难取深层土样, 故采样深度只达到30 cm)。将样品分水曲柳落叶松相邻行间; 水曲柳带内1、2行间; 落叶松带内1、2行间(与水曲柳相邻行为第1行)和落叶松带内2、3行间归类处理。在水曲柳纯林和落叶松纯林内按同样方法各设置1块样地, 并按同样方法进行取样。将采集到的根系样品放入尼龙纱网中, 用水冲洗干净, 阴干。分别不同树种按直径分为3级:≤2 mm, 2 ~ 5 mm, ≥5 mm, 并分别称鲜重, 取样在80 ℃下烘干8 h, 计算含水率, 求算生物量并计算根密度。
1.2 温室床栽和盆栽实验 1.2.1 床栽实验栽植实验在东北林业大学帽儿山实验林场尖砬沟森林培育实验站温室内进行。栽植土壤为草甸土, 实验持续2个生长季。实验苗木为2年生, 栽前水曲柳平均地径为0.57 cm, 平均高为40.3 cm; 落叶松平均地径为0.75 cm, 平均高为35.2 cm。用于栽培的床面长80 cm, 宽40 cm。为防止床内所栽苗木根系伸出床外, 在床的四周用聚乙烯塑料膜相隔。栽植分落叶松纯栽、水曲柳纯栽、落叶松与水曲柳混栽3种方式。每床栽两行共12株苗木, 株行距为12 cm ×20 cm。纯栽各重复16次, 混栽采用株间混交重复32次。生长季结束时, 纯栽落叶松平均地径为1.28 cm, 平均高为142.4 cm; 纯栽水曲柳平均地径为1.12 cm, 平均高为115.0 cm; 混栽的落叶松平均地径为1.13 cm, 平均高为119.8 cm; 混栽的水曲柳平均地径为1.25 cm, 平均高为127.7 cm。在实验结束时, 对每株苗木分叶、茎(包括枝)、直径>2mm的根、直径≤2 mm的根测定生物量。
1.2.2 盆栽实验所用容器是底径22.5 cm, 上径29.0 cm, 高30.0 cm的聚乙烯塑料桶。所用土壤为典型暗棕壤, 每桶10.0 kg。实验苗木与床栽相同, 每盆栽植4株, 分落叶松纯栽、水曲柳纯栽和混栽。纯栽各重复24次, 混栽每盆2株水曲柳2株落叶松, 重复48次。实验持续1个生长季。生长季结束时, 纯栽落叶松平均地径为0.83 cm, 平均高为50.9 cm; 纯栽水曲柳平均地径为1.12 cm, 平均高为69.3 cm; 混栽的落叶松平均地径为0.80 cm, 平均高为47.4 cm; 混栽的水曲柳平均地径为1.27 cm, 平均高为70.6 cm。实验后的生物量测定与床栽相同。
2 结果 2.1 混交林中水曲柳落叶松根系分布的变化 2.1.1 根系的水平分布变化在混交林中, 从水曲柳与落叶松相邻行开始, 随着与落叶松的接近, 水曲柳在各行间的根密度依次降低(表 2)。然而在水曲柳与落叶松相邻行间和水曲柳带内1、2行间水曲柳根密度则相近, 分别为3237.2 g·m-3和3130.2 g·m-3, 仅略低于纯林中的3718.1 g·m-3, 3者差异不明显(dF =95, F =2.96 < F0.05 =3.09)。在落叶松带内1、2行间和2、3行间水曲柳的根密度较低, 分别为1154.5 g·m-3和296.6 g·m-3, 二者与水曲柳落叶松相邻行间的根系密度差异达极显著(落叶松带内1、2行间:dF =63, F =8.64 >F0.01 =7.08;落叶松带内2、3行间:dF =63, F =14.20 >F0.01 =7.08)。在水曲柳与落叶松相邻行间, 水曲柳的根密度并未因与落叶松相邻而出现下降, 看来与落叶松混交有利于水曲柳根系的生长。在各行间水曲柳的根量均以≤2 mm的细根为主, 在水曲柳与落叶松相邻行间≤2 mm的根密度为2288.9 g·m-3高于水曲柳带1、2行间的2074.5 g·m-3, 出现增加趋势。
|
|

在混交林中, 从落叶松带内2、3行间开始, 随着与水曲柳的接近, 落叶松在各行间的根密度逐渐下降(表 2)。在落叶松带2、3行间, 落叶松的根密度出现最大值为3717.2 g·m-3, 比纯林增加22.2 %。在落叶松带1、2行间, 根密度为2745.3 g·m-3, 开始下降, 比落叶松带2、3行间低35.4 %, 比纯林低10.8 %, 落叶松带2、3行间的根密度与落叶松带1、2行间的差异显著(dF =63, F =4.23 >F0.05 =4.00), 与纯林的差异不显著(dF =63, F =3.80 < F0.05 =4.00)。在水曲柳落叶松相邻行间和水曲柳带1、2行间, 落叶松的根密度相近, 分别为939.4 g·m-3和702.1 g·m-3, 明显低于其它行间(水曲柳落叶松相邻行间和落叶松带1、2行间比较:dF =63, F =7.82 >F0.01 =7.08)。由于在水曲柳落叶松相邻行间落叶松的根密度明显降低, 因此, 落叶松的根系可能受到水曲柳的竞争抑制。在水曲柳落叶松相邻带间和水曲柳带1、2行间, 落叶松无≥5 mm的根系分布, 而≤2 mm的根系占绝对优势。
混交林中各行间总的根系密度(水曲柳与落叶松根密度相加)均高于水曲柳和落叶松纯林(表 2)。尤其在水曲柳落叶松相邻行间根系总密度最大, 达4176.6 g·m-3, 比水曲柳纯林高12.3 %, 比落叶松纯林高37.4 %(经检验达极显著差异:dF =63, F =7.64 >F0.01 =7.08)。两树种的纯林相比, 落叶松的根密度较低, 水曲柳的根密度较高, 这可能是由树种的生物学特性决定的。
2.1.2 根系的垂直分布变化与纯林相比混交林中水曲柳的根系垂直分布更加均匀(表 3), 在水曲柳与落叶松相邻行间, 水曲柳在0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别占总根量的33.3 %、32.7 %和34.0 %。而在水曲柳纯林中在0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别为总根量的61.0 %、31.6 %和7.4 %。纯林中的水曲柳根系主要分布于0 ~ 10 cm和10 ~ 20 cm土层内, 20 ~ 30 cm土层内仅有极少量的根系分布。在落叶松带内1、2行间水曲柳的根系垂直分布也较均匀。而在水曲柳带内1、2行间水曲柳根系的垂直分布与纯林相似, 在0 ~ 10 cm和10 ~ 20 cm土层内的根量占总根量的84.1 %, 根系主要集中分布于表层。混交使水曲柳的根系分布加深, 有利于水曲柳吸收土壤深层的水分和养分。
|
|
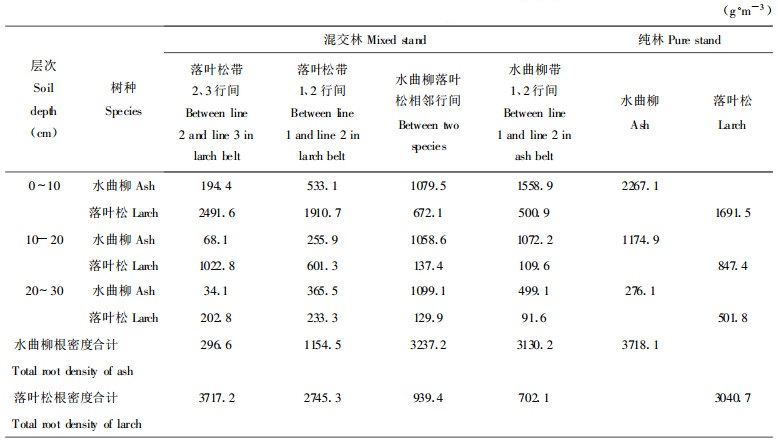
在混交林中落叶松的根系分布更趋向于表层(表 3), 在落叶松纯林中0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别占总根量的55.6 %、27.9 %和16.5 %, 而在在水曲柳与落叶松相邻行间, 落叶松在0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别为总根量的71.3 %、15.6 %和13.1 %。随着与水曲柳的接近落叶松的根系垂直分布表现出向表层增加的趋势。
混交使总的根系(水曲柳根量与落叶松根量之和)垂直分布更加均匀(表 3), 在水曲柳与落叶松相邻行间, 根系的垂直分布最均匀, 0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别为总根量的41.9 %、28.7 %和29.4 %。而在水曲柳带内1、2行间和落叶松带内1、2行间, 表层0 ~ 10 cm的根系比例开始增加分别为53.7 %和62.7 %, 底层20 ~ 30 cm的根系比例下降分别为15.4 %和15.4 %。与混交林中的水曲柳落叶松相邻行间相比, 两树种纯林的根系垂直分布主要集中于表层。
2.2 温室混栽幼苗根系的生长混栽后两树种的根系生长均发生变化(图 1), 在盆栽条件下, 与落叶松混栽的水曲柳根系生长加快, 根系总生物量、≤2 mm根生物量和>2 mm根生物量, 分别比纯林增加21.2 %、28.8 %和16.8 %。经显著性检验≤2 mm根生物量差异明显(dF =47, F =4.28 >F0.05 =4.05), 总生物量和>2 mm根生物量差异不明显(总生物量:dF =47, F =2.91 < F0.05 =4.05;>2 mm根生物量:dF =47, F =2.54 < F0.05 =4.05)。与水曲柳混栽的落叶松根系生长下降, 根系总生物量、≤2 mm根生物量和>2 mm根生物量, 分别比纯栽下降20.8 %、25.0 %和13.5 %, 2者差异不明显(总生物量:dF =47, F =2.87 < F0.05 =4.05;≤2 mm根生物量:dF =47, F =3.67 < F0.05 =4.05; >2 mm根生物量:dF =47, F =2.33 < F0.05 =4.05)。

|
图 1 混栽对水曲柳落叶松根生长的影响 Fig. 1 Effects of ash and larch in mixed planted on root growth
 ≤2 mm 根生物量≤2 mm Root mass; ≤2 mm 根生物量≤2 mm Root mass; >2 mm 根生物量>2 mm Root mass.L:纯栽落叶松 Larch in pure planted;ML:混栽落叶松 Larch in mixed planted with ash;MF:混栽水曲柳 Ash in mixed planted with larch;F:纯栽水曲柳 Ash in pure planted.a:盆栽试验 Pot experiments;b:床栽试验 Seedbed experiments. >2 mm 根生物量>2 mm Root mass.L:纯栽落叶松 Larch in pure planted;ML:混栽落叶松 Larch in mixed planted with ash;MF:混栽水曲柳 Ash in mixed planted with larch;F:纯栽水曲柳 Ash in pure planted.a:盆栽试验 Pot experiments;b:床栽试验 Seedbed experiments.
|
在床栽条件下, 与落叶松混栽的水曲柳根系生长也表现增加, 根系总生物量、≤2 mm根生物量和>2 mm根生物量, 分别比纯栽增加17.9 %、36.5 %和14.6 %。经显著性检验≤2 mm根生物量明显增加(dF =31, F =4.42 >F0.05 =4.17), 总生物量和>2 mm根生物量差异不明显(总生物量:dF =31, F = 2.75 >F0.05 =4.17;>2 mm根生物量:dF =31, F =2.27 >F0.05 =4.17)混栽的落叶松根系与纯栽相比生长也表现下降, 根系总生物量、≤2 mm根生物量和>2 mm根生物量, 分别比纯林下降18.7 %、18.2 %和18.8 %, 但二者差异不明显(总生物量:dF =31, F =2.97 >F0.05 =4.05;≤2 mm根生物量:dF =31, F =2.82 >F0.05 =4.17;>2 mm根生物量:dF =31, F =2.99 > F0.05 =4.17)。
与纯栽相比混栽落叶松的地下地上比表现出下降的趋势, 盆栽时混栽和纯栽落叶松的地下地上比分别为0.40和0.45, 床栽时分别为0.24和0.30(图 2)。与纯栽相比混栽水曲柳的地下地上比无明显变化趋势。水曲柳与落叶松相比, 水曲柳的地下地上比较高, 盆栽时为0.78 ~ 0.81, 床栽时为0.52 ~ 0.55, 约是落叶松的2倍。而且在混栽时水曲柳根生物量约是落叶松的4 ~ 6倍, 这可能是水曲柳具有较强竞争力的主要原因。

 盆栽地下 地上 Root shoot ratio of potted seedlings;
盆栽地下 地上 Root shoot ratio of potted seedlings;
 床栽地下 地下 Root shoot ratio of seedbed seedlings.
床栽地下 地下 Root shoot ratio of seedbed seedlings.
混栽对两树种地上部生物量均产生影响(图 3), 混栽落叶松与纯栽相比地上生物量下降, 盆栽时下降20.4 %, 床栽时下降34.0 %。而混栽水曲柳的地上部生物量与纯栽相比则表现增加, 盆栽时增加17.0 %, 床栽时增加23.3 %。

 茎生物量 Shoot mass;
茎生物量 Shoot mass; 叶生物量 Leaf mass.
叶生物量 Leaf mass.
混交林中各树种根系的生长和分布受树种特性、种间竞争以及林地营养条件改善等多方面因素影响, 到目前为止虽然有一些研究结果(王政权等, 2000; 刘春江, 1985; 李振问等, 1993; 国庆喜等, 1991; 廖利平等, 1999; 翟明普等, 1982; Mckay et al., 1988), 但我们很难从这些结果中找到普遍的规律。
在温室栽培实验中, 无论盆栽和床栽, 混栽水曲柳的根生物量和地上部生物量均高于纯栽的, 尤其是≤2 mm的细根生物量表现明显增加。而混栽时落叶松的根生物量和地上部生物量则低于纯栽的。水曲柳的地下地上比约是落叶松的2倍, 从绝对量上看, 混栽时水曲柳的根生物量约是落叶松的4 ~ 6倍。水曲柳将大部分碳化物分配给地下部, 使其在地下竞争中占有优势。Ennik等(格雷戈里, 1992)对Lolium纯系黑麦草进行的竞争实验表明, 最有竞争性的黑麦草纯系, 它们始终具有较大的根系和地下地上比率。竞争能力强的纯系在单种栽培时产量常常较低, 然而在与其它竞争能力弱的纯系一起栽培时产量出现增加。纯栽和混栽时, 水曲柳落叶松在生长上的表现与Ennik等的纯系实验非常相似。由于树木幼苗根系较小, 对土壤条件的改善作用较弱。因此, 此时竞争是影响混栽幼苗根系及地上部生长的主要因素。
在单种栽培下, 当土壤养分和水分缺乏时, 树木将更多的生物量分配给根系, 出现较高的地下地上比(Fabião et al., 1995)。目前还很少有证据证明混栽时处于竞争中的植物也具有类似的适应性。但在我们的实验中, 混栽条件下的落叶松地下地上比均高于纯栽的。有可能是落叶松在混栽时受到较强养分水分竞争的一种反应。
在水曲柳落叶松混交林中水曲柳和落叶松根系的水平公布表现不同。在水曲柳落叶松相邻行间和水曲柳带1、2行间, 水曲柳根密度无明显差异, 而且在落叶松带内也有一定数量的水曲柳根系分布。然而, 在水曲柳落叶松相邻行间落叶松的根密度却明显低于落叶松带1、2行间和2、3行间的, 而且在水曲柳带内也只有少量的落叶松根系分布。国庆喜等(1991)对26 a生的水曲柳落叶松混交林的研究也得出类似的结论。结果不考虑种间的相互作用, 在水曲柳落叶松相邻行间, 由于水曲柳的相邻行为落叶松, 水曲柳的根密度应出现明显低于水曲柳带1、2行间的趋势。水曲柳在水曲柳落叶松相邻行间未出现根密度下降可能与两方面因素有关。一是落叶松对林地营养条件的改善, 促使水曲柳根密度增加。目前已有研究证明, 落叶松可以明显提高林地土壤磷的有效性(张彦东, 1997), 而养分条件的改善可以促进树木根系的生长。Mou等(1995)和Geroge等(1997)的研究表明, 土壤养分的空间异质性能够改变细根的密度, 在养分较肥沃的土壤斑块内, 植株的细根密度增加。Fabião等(1995)的施肥实验也证明, 施肥改变了表层土壤的养分状况, 导致树木的细根主要分布于土壤表层。二是树种竞争能力的差异, 促使水曲柳根密度增加, 落叶松根密度降低。如果仅是由于落叶松改善了混交林中的土壤养分状况, 而导致根系分布发生变化, 还不能很好地解释在相邻行间落叶松根系明显降低的现象。竞争对混交林根系分布具有重要影响, 翟明普(1982)对油松栓皮栎混交林的研究表明, 由于种间竞争能力的差异, 使混交林中的油松细根量和粗根量降低。在我们的盆栽和床栽实验中, 水曲柳的地下地上比和根生物量均明显高于落叶松, 说明水曲柳具有较大的地下竞争能力。在水曲柳落叶松相邻行间出现落叶松根密度明显下降的现象, 可能是水曲柳通过较大的根系, 占据较大的土壤空间, 并部分排斥落叶松的根系所致。
水曲柳落叶松混交林中的根系垂直分布更加均匀, 在水曲柳与落叶松相邻行间, 0 ~ 10 cm、10 ~ 20 cm和20 ~ 30 cm土层内的根量分别为总根量的41.9 %、28.7 %和29.4 %。而在水曲柳纯林中分别为61.0 %、31.6 %和7.4 %; 在落叶松纯林中分别为55.6 %、27.9 %和16.5 %。刘春江(1985), 李振问等(1993), 国庆喜等(1991), 廖利平等(1999)对不同混交林的研究均得出类似的结论。看来这是一种较普遍的现象, 然而值得注意的是这种现象几乎都出现在成功的混交林中, 此时混交林的产量均高于纯林, 混交林中的树木在生长上也优于纯林, 混交林中树木根系的数量也要高于纯林中的。当表层土壤无充足的养分和水分供应时, 根系自然要向土壤深层伸展。Bernat等(1996)的疏伐实验表明, 未疏伐的林分密度较高, 根系数量较大, 根系向深层分布。在我们研究的水曲柳落叶松混交林中, 各行间的根系总密度均高于纯林的, 而且混交林地上部的生长也优于纯林(表 1)。所以可能是因混交林中树木生长加快, 根系量增加, 导致根系向深层发展。当然种间竞争也可能影响混交林中根系的垂直分布, 但我们还无法弄清竞争与根系垂直分布的关系, 有待今后继续研究。
总之, 竞争对混交林中树种的根系分布会产生一定影响, 竞争能力较强的树种在混交林中根系分布范围较广, 竞争能力弱的树种根系分布受到一定排斥。水曲柳具有较大的根系, 竞争能力强, 落叶松的竞争能力相对较弱。当水曲柳与落叶松混栽时, 水曲柳的根系生长加快, 而落叶松根系生长降低。竞争能力的差异是导致混交林中水曲柳增产的主要原因之一, 这一点常常被人们忽视。
格雷戈里P J等编著.作物根的发育与功能.陈放, 周桂梅, 陈浩编译.第1版.成都: 四川大学出版社, 1992, 225~ 243
|
国庆喜, 陈乃全, 陈志国. 1991. 水曲柳落叶松混交林根系的研究. 东北林业大学学报(水胡黄椴专刊): 74-80. |
黄建辉, 韩兴国, 陈灵芝. 1999. 森林生态系统根系生物量研究进展. 生态学报, 19(2): 270-277. DOI:10.3321/j.issn:1000-0933.1999.02.021 |
廖利平, 杨跃军, 汪思龙, 等. 1999. 杉木火力楠纯林及其混交林细根分布、分解与养分归还. 生态学报, 19(3): 342-346. DOI:10.3321/j.issn:1000-0933.1999.03.009 |
廖利平, 陈楚莹, 张家武, 等. 1995. 杉木、火力楠纯林及混交林细根周转的研究. 应用生态学报, 19(3): 342-346. |
李振问, 杨玉盛, 吴擢溪, 等. 1993. 杉木、火力楠混交林根系的研究. 生态学杂志, 12(1): 20-24. DOI:10.3321/j.issn:1000-4890.1993.01.008 |
刘春江. 1985. 北京西山地区人工油松栓皮栎混交林根系研究初报. 北京林学院学报, 7(1): 77-84. |
田蒴宁.植物竞争研究进展.见刘建国主编.当代生态学博论.第1版.北京: 中国科学技术出版社, 1992, 40~ 48
|
王政权, 张彦东. 2000. 水曲柳落叶松根系之间的相互作用研究. 植物生态学报, 24(3): 346-350. DOI:10.3321/j.issn:1005-264X.2000.03.017 |
张彦东, 于兴君, 杨文化.水曲柳落叶松混交林土壤磷活化机理.见沈国肪等主编.混交林研究.第1版.北京: 中国林业出版社, 1997, 136~ 140
|
翟明普. 1982. 北京西山地区人工油松元宝枫混交林根系的研究. 北京林学院学报, 4(1): 1-11. |
Bernat L, Santiago S, Carlos G. 1998. Fine roots dynamics in a Mediterranean forest:effects of drought and stem density. Tree Physiology, 18: 601-606. DOI:10.1093/treephys/18.8-9.601 |
Fabião A, Madeira M, Steen E, et al. 1995. Development of root biomass in an Eucalyplus globulus plantation under different water and nutrient regimes. Plant and Soil, 168-169: 215-223. DOI:10.1007/BF00029331 |
Geroge E, Seith E, Schaeffer C, et al. 1997. Responses of Picea, Pinus and Pseudotsuga root to heterogeneous nutrient distribution in soil. Tree Physiology, 17(1): 39-45. DOI:10.1093/treephys/17.1.39 |
Mckay H M, Malcolm D C. 1988. Acomparison of the fine root component of a pure and mixed coniferous stand. Canadian Journal of Forestry Research, 18: 1416-1426. DOI:10.1139/x88-220 |
Mou P, Jonse RH, Mitchell R J, et al. 1995. Spatial distribution of roots in sweetgum and Loblolly pine monocultures and relations with above-ground biomass and soil nutrients. Functional Ecology, 9(3): 689-699. |
Sword M A, Gravatt D A, Faulkner P L, et al. 1996. Seasonal branch and fine root growth of juvenile loblolly pine five growing seasons after fertilization. Tree Physiology, 16: 889-904. |