 2001, Vol. 37
2001, Vol. 37文章信息
- 白尚斌, 张彦东, 王政权.
- Bai Shangbing, Zhang Yandong, Wang Zhengquan.
- 落叶松根际pH值与供磷水平及土壤磷有效性的关系
- THE RELATIONSHIP BETWEEN pH CHANGES AND P-AVAILABILITY IN RHIZOSPHERE OF LARIX GMELINII
- 林业科学, 2001, 37(4): 129-133.
- Scientia Silvae Sinicae, 2001, 37(4): 129-133.
-
文章历史
- 收稿日期:2001-01-18
-
作者相关文章


2. 中国科学院西双版纳热带植物园 昆明 650223;
3. 东北林业大学森林资源与环境学院 哈尔滨 150040
2. Xishuangbanna Tropical Botanic Garden, Chinese Academy of Sciences Kunming 650223;
3. Forest Resource and Environment College, Northeast Forestry University Harbin 150040
植物在生长过程中由于根分泌物的作用, 根际pH值将发生不同于土体的变化。这种变化在植物利用难溶性营养元素(如磷素)上是极其重要的。迄今为止关于草本植物和作物根际pH值变化与土壤中难溶性磷利用的研究已有很多报道。Grinsted和Hedley (1986)研究发现生长35天的油菜幼苗根际pH值可降低2.4个单位(由6.5到4.1), 相应其根际有效磷的浓度增加10倍(王庆仁等, 1998)。Marchner (1986)发现白羽扇豆在缺磷土壤中形成排根, 排根分泌的有机酸能降低根际土壤的pH值以促进其对磷的吸收。目前人们普遍认为, pH值降低主要是有利于土壤中难溶性Ca-P的溶解, 从而提高了磷的有效性(陆景陵, 1994; Hinsinger et al., 1995)。但关于树木根际pH值变化的研究报道目前相对较少(Gijsman A J, 1990;Gillespie et al., 1989)。在国内近年经研究发现, 杉木(Cunninghamia lanceolata)在不同发育阶段, 根际土壤速效磷均高于非根际土壤(杨承栋等, 1999), 杉木根际土壤pH普遍低于非根际土壤(陈竑竣等, 1994); 在落叶松(Larix gmelinii Rupr.)根际也有同样的变化趋势(张彦东等, 1997; 崔晓阳等, 1994), 因此, 有人认为落叶松根际土壤磷有效性的增加可能与根际pH值的变化有关(崔晓阳等, 1994)。但由于这些研究都是在野外林地采样分析进行的, 其他植物的根系干扰难以避免, 精确测定某种树木的根际pH十分困难。假设落叶松根际土壤有效磷的增加是由根际pH值的变化引起的, 那么必将存在根际pH值与根际有效磷的相关关系, 另一方面, 在缺磷条件下, 如果落叶松本身具有通过分泌质子或有机酸来降低根际土壤pH值, 那么在不同供水平下, 落叶松根际pH值将存在较大的差异。这样, 我们就可以说明在缺磷条件下, 落叶松具有主动的适应机制, 可以通过降低根际pH值来活化根际磷。反之, 落叶松根际有效磷的提高可能是由于其他原因引起的。为此本研究的主要目的是采用室内显色测定与野外林地取样相结合的方法测定落叶松根际pH值, 以探讨落叶松根际pH值变化与供磷水平及土壤磷有效性的关系。
1 材料与方法本研究以温室砂培和野外现实林分取样两种方法进行。
1.1 温室砂培试验供试苗木及处理 试验所用苗木为兴安落叶松(Larix gmelinii Rupr.), 分当年生与2年生两批。当年生落叶松苗木于6月下旬从苗床取出, 2年生落叶松苗木于5月初取出。苗木取出后根部用清水漂洗, 用0.5%次氯酸钠(NaClO)溶液消毒, 再用蒸馏水漂洗3次, 然后用于砂培。
苗木培养 苗木培养基质为石英砂, 先将石英砂洗去泥土, 经0.5%盐酸浸泡24h, 用自来水冲洗至中性。然后将石英砂装入容积为0.13m3的塑料盆中, 每盆装砂13kg。将上述经过处理的苗木分别栽植在盆中, 2年生苗每盆植苗5株, 当年生苗每盆植苗50株。然后以不同磷浓度的营养液进行浇灌。营养液的磷浓度共设4种:正常营养液(P的浓度为0.071g·L-1); 缺磷营养液(P的浓度为0);低磷营养液(P的浓度为0.035g/L)和高磷营养液(P的浓度为0.142g·L-1)。2年生苗木的每个处理5盆(相当于5个重复), 共20盆。当年生苗木每个处理2盆(相当于2个重复), 共8盆。上述每个处理每天浇一次营养液, 每盆每次浇100ml。当年生苗木在室温内培养40d。2年生苗木培养90d。然后取样测定其根际pH值。
营养液配方 大量元素用Hoagland配方(上海植物生理学会, 1985) :KNO3 0.51g/L, Ca (NO3) 20.82g·L-1, MgSO4·7H2O 0.49g·L-1, KH2PO4 0.136g·L-1。微量元素用Arnon配方(上海植物生理学会, 1985) :H3BO3 2.86mg·L-1, CuSO4·5H2O 0.08mg·L-1, ZnSO4·7H2O 0.22mg·L-1, MnCl2·4H2O 1.81mg·L-1, H2MoO·4H2O 0.09mg·L-1, Fe-EDTA 20mg·L-1。pH调至6.0。缺磷营养液中不含KH2PO4, K+以等量KCl补齐。
显色测定 将培养了40d的当年生落叶松苗木从盆中取出, 立即用蒸馏水中去根表面粘附的砂粒, 平铺在滤纸上吸去多余的水分, 并用平头镊子将根展开放入盛有调试好的琼脂-脂示剂混合液的培养皿中培养。琼脂-指示剂混合液是由称取10mg间苯二酚蓝(变色范围pH4.0~6.4)溶于10ml无水乙醇后加蒸馏水50ml再加入1g琼脂, 加热溶解, 充分摇匀制成。试验前pH值调至6.4, 颜色为深蓝色。将培养了90d的2年生落叶松苗木进行同样的处理。每个处理重复10次。同时与标准指示剂-琼脂溶胶作对照, 判读各根际的pH值(根尖后1.5cm范围内)。
1.2 野外现实林分取样采样方法 在东北林业大学帽儿山实验林场, 选择12年生, 21年生和40年生的落叶松人工纯林, 设置0.1hm2的标准地作为采集土样的样地, 按“S”形在标准地内分别设置30个采样点, 挖掘林下表层土壤(0~20cm), 取出落叶松根系, 轻轻拍打抖动, 抖落的土壤为非根际土, 待沾根的大块土除去后, 再将附着在根系上的土壤用刷子刷下即为根际土样品。
土壤pH值和有效磷的测定 水浸pH值测定采用电位法(劳家柽, 1988)。全磷测定采用H2SO4-HClO4酸溶-钼锑抗比色法(劳家柽, 1988), 有效磷测定以0.03mol·L-1的NH4F-0.025mol·L-1的HCl作为浸提剂, 钼锑抗比色法测定(劳家柽, 1988)。
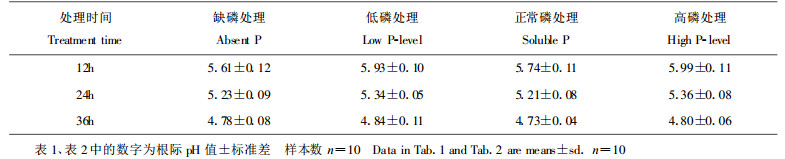
2 结果 2.1 落叶松幼苗根际pH值的变化对培养皿中显色测定的落叶松幼苗经观察发现, 当年生幼苗显色12h后, 4种处理苗木根尖及稍后部位根区的颜色均开始由蓝变红, 说明根系出现分泌物, 根际pH值开始下降。24h时颜色加深且变化区域加宽, 36h时上述现象趋于稳定。2年生苗木根区同样可以看出上述变化过程, 只是变化的速率更快些, 在24h即趋于稳定。还可以看到这种颜色的变化在新生根周围尤为明显, 红色最深的区域集中于根尖稍后的部位, 根基部周围的颜色变化较弱。
为了便于比较, 我们对根尖后1.5cm范围内的根际pH值进行了判读(表 1, 表 2)。结果表明, 当年生苗木的4种处理在36h显色稳定时, pH值均降至5.0以下, 最低的为4.73, 最高的为4.84, 降低1.56~1.67个单位。根据土壤酸性的定义, 此范围pH值为微酸性。对各处理间pH值进行方差分析表明, 各处理间pH值相差不明显(F=1.06 < F0.05 (3, 36) =2.87)。2年生苗木的变化过程与当年生苗相似, 只不过是达到稳定的时间更快些。2年生苗木在达到稳定时, 根际pH均在5.0以上, 最低的为5.30, 最高的为5.50, 降低0.90~1.10个单位。要高于当年生苗木稳定时的pH值, 平均高约0.60个单位。比较显色稳定时各处理间pH值表明, 其差异亦不明显(F=1.12 < F0.05 (3, 36) =2.87)。
|
|
|
|
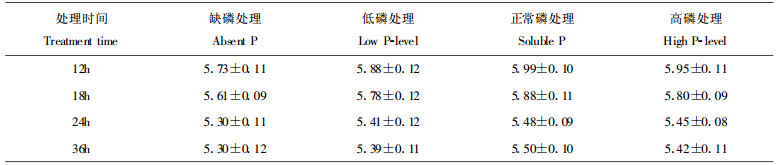
本实验落叶松根际pH值变化显色较慢, 在显色2h后观察, 根周围颜色几乎无变化。在6h后观察, 根周围颜色才略有变化。12h后观察, 根周围颜色已有明显变化, 但还不稳定, 在36h后才达到稳定。有些草本及农作物显色测定判读时间一般为显色后2h, 树木和草本及农作物在根分泌物分泌速率上可能存在一些差异。如果不考虑根际周围颜色是否稳定, 而以12h出现明显变色时pH作为落叶松根际pH的判读值的话, 则当年生幼苗根际pH值的变化范围为5.61~5.99, 2年生幼苗的变化范围为5.73~5.99, 比36h的判读值高。
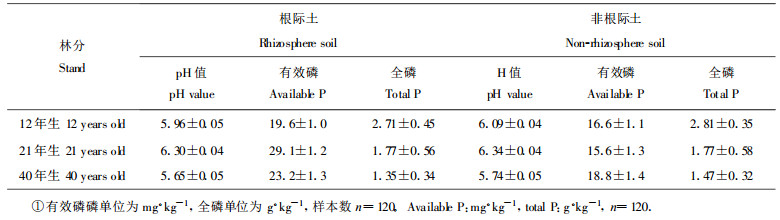
2.2 现实林分根际pH值变化与磷有效性的关系通过测定现实林分根际与非根际土壤的pH值表明(表 3), 无论是12年生、21年生、还是40年生落叶松根际的pH值都低于非根际, 分别低0.11、0.04和0.09个单位。虽然落叶松根际土的pH值低于非根际土, 但经差异显著性检验, 两者相差不明显(F=2.06 < F0.05 (1, 58) =4.01), 说明在现实林分中落叶松根际酸化现象不明显。比较各林分根际土与非根际土有效磷含量表明, 根际土有效磷均高于非根际土。12年生林分高出18.1%, 21年生林分高出86.5%, 40年生林分高出23.4%。经差异显著性检验, 有效磷在根际与非根际差异显著(F=4.62 < F0.05 (1, 58) =4.01)。对根际土与非根际土的全磷含量进行比较表明, 根际土的全磷含量与非根际土的含量相等或略低于非根际土。这说明根际土有效磷的增加并不是由于根际土全磷含量增高引起的。许多研究认为根际pH值降低有利于土壤难溶性Ca-P的活化(Hinsinger et al., 1995; Hoffland et al., 1998a), 从而增加磷的有效性。我们对pH值与有效磷作相关分析得出相关系数r=-0.026 (n=120), 经t检验(|t|=0.26809 < t0.05 (120) =1.980)得知二者的相关不显著。这说明根际有效磷的增高与pH值的降低不存在显著的线性相关, 有效磷的提高可能是由于其它原因引起的。
|
|
目前的研究认为, 引起植物根际pH值变化的原因主要有(1)阴阳离子吸收不平衡; (2)根际呼吸作用产生的CO2; (3)根系分泌的有机酸; (4)根分泌H+。当受到磷营养胁迫时, 有些植物具有主动适应的机制, 可通过降低根际pH值的方式增加磷的吸收(Moorby et al., 1988; Gahoonia et al., 1992)。而此时根际pH值的调节, 主要是通过根分泌有机酸或H+数量的增加使根际酸化(郭朝晖等, 1999)。具有这种主动调节机制的植物其主要特点是根际pH值变化与供磷水平密切相关。在本实验条件下, 无论缺磷与否, 落叶松根际pH值都表现下降, 而且在各种不同磷浓度处理间, 根际pH值差异不明显。即使在磷胁迫条件下, 也没有出现根际pH值明显降低的现象, 说明落叶松根际pH值的变化不受磷胁迫的影响, 落叶松不存在通过酸化根际而使土壤磷活化的主动适应机制。虽然落叶松根际pH值出现降低, 但这种下降与供磷水平无关。我们认为这种酸化现象是根系进行正常呼吸产生的CO2, 根分泌的有机酸和根分泌H+引起的。由于本实验N素是以NO3-的形式供给的, 所以阴阳离子吸收不平衡对根际pH的下降影响不大。
苗木在代谢过程中, 分泌的H+不断在根际累积, 使培养基质酸化, 颜色由原来的蓝色变为红色。变色时间直接受H+分泌速率影响, 变色深度与该区域H+的浓度有关, 而区域的大小由根分泌H+的数量决定。2年生苗木根际的颜色变化较快, 说明2年生苗木代谢更快些, 即分泌质子的速度较快。当年生苗木36h稳定时颜色较深, 说明当年生苗木根分泌的浓度高于2年生苗木。我们发现在同一批幼苗4种处理的变色时间、区域大小以及变色深度都不存在明显差异, 这进一步说明落叶松根际pH值的变化不是由磷胁迫引起的。
在野外树木根际经土壤的缓冲作用, 结果常与室内培养法测定有差异。虽然落叶松根际土的pH值低于非根际土, 但经方差分析检验得出两者差异不显著, 说明落叶松根际酸化现象不明显。由于室内栽培试验表明, 落叶松根际pH值下降不受供磷水平的影响, 野外现实林分中, 其根际pH值有下降的趋势可能是落叶松在生长过程中自身根系分泌一些酸性物质引起的。而有效磷在根际土与非根际土之间差异显著, 说明磷在落叶松根际发生了活化作用。许多研究认为根际pH值降低有利于土壤难溶性Ca-P的活化(Hinsinger et al., 1995; Hoffland et al., 1989a), 从而增加磷的有效性。我们对pH值与有效磷作相关分析知二者的线性相关不显著。这说明以Ca-P形态存在的无机磷可能不是落叶松根际土壤中有效磷的主要来源。有效磷的提高可能是由于其它原因引起的, 深入机理有待今后继续探讨。在林地出现根际土的pH值稍低于非根际的趋势, 但不能导致明显酸化效果是受到了林地土壤的缓冲能力以及土壤湿度的影响。Gillespie等(1990)对土壤的pH缓冲能力的研究证实了这一点。低的土壤缓冲能力和高的土壤湿度使根分泌的H+迅速向林地土壤中扩散, 不能在根际聚集, 导致现实林分根际pH梯度不明显。
陈竑竣, 李传涵. 1994. 杉木根际与非根际土壤酶活性比较. 林业科学, 30(2): 170-175. |
崔晓阳, 等. 1994. 落叶松水曲柳幼林土壤有效磷的空间分布特点.现代森林经营与资源利用研究. 哈尔滨: 黑龙江省科学技术出版社, 231-240.
|
郭朝晖, 张扬珠, 黄子蔚. 1999. 根际微域营养研究进展(一). 土壤通报, 30(1): 46-封3. DOI:10.3321/j.issn:0564-3945.1999.01.017 |
劳家柽主编. 土壤农化分析手册. 北京: 农业出版社, 1988
|
陆景陵. 1994. 植物营养学. 北京: 北京农业大学出版社, 108-111.
|
上海植物生理学会编. 植物生理学实验手册. 上海: 上海科技出版社, 1985
|
王庆仁, 李继云, 等. 1998. 植物高效利用土壤难溶态磷研究及展望. 植物营养与肥料学报, 4(2): 107-116. DOI:10.3321/j.issn:1008-505X.1998.02.003 |
杨承栋, 焦如珍. 1999. 杉木人工林根际土壤性质变化的研究. 林业科学, 35(6): 2-9. DOI:10.3321/j.issn:1001-7488.1999.06.001 |
张彦东, 于兴君, 杨文化. 水曲柳落叶松混交林林地与根际土壤养分特性. 见沈国舫, 翟明普. 混交林研究. 北京: 中国林业出版社, 1997: 145~ 149
|
Gahoonia T S, Nielsen N E. 1992. The root induced pH change on the deletion of inorganic and organic phosphorous in the rhizosphere. Plant and soil, 143: 183-189. |
Gijsman A J. 1990. Rhizosphere pH along different root zone of Douglas-fir (Pseudtsuga menziesii), as affected by source of nitrogen. Plant and Soil, 124: 161-167. DOI:10.1007/BF00009254 |
Gillespie A R, Pope P E. 1989. Alfalfa N2-fixation enhances the phosphorous uptake of walnut in interplantings. Plant and soil, 113: 291-293. DOI:10.1007/BF02280196 |
Gillespie A R, Pope P E. 1990. Rhizosphere Acidification Increase Rhosphorus Recovery of Black Locust: Ⅰ. Induced Acidification and Soil Response. Soil Sci. Soc. Am. J., 54: 533-537. DOI:10.2136/sssaj1990.03615995005400020041x |
Hinsinger P, Gilkes R J. 1995. Root-induced Dissolution of Phosphate Rock in the Rhizosphere of Lupins Grown in Alkakine Soil. J. Soil Res., 33: 477-489. DOI:10.1071/SR9950477 |
Hoffland E, Findenegg G R, Nelemans J A. 1989a. Solubilization of rock phosphate by rape: Ⅰ. Evaluation of the role of the nutrient uptake pattern. Plant and Soil, 113: 155-160. DOI:10.1007/BF02280175 |
Marschner H. 1986. Mineral nutrition of high plant. London UK: Academic Press.
|
Moorby H, White R E, Nye P H. 1988. The influence of phosphate nutrition on H ion efflux from the roots of young rape plants. Plant and soil, 105: 247-256. DOI:10.1007/BF02376789 |