 2001, Vol. 37
2001, Vol. 37文章信息
- 杨万勤, 钟章成, 陶建平, 何维明.
- Yang Wanqin, Zhong Zhangcheng, Tao Jianping, He Weiming.
- 缙云山森林土壤酶活性与植物多样性的关系
- STUDY ON RELATIONSHIP BETWEEN SOIL ENZYMIC ACTIVITIES AND PLANT SPECIES DIVERSITY IN FOREST ECOSYSTEM OF MT.JINYUN
- 林业科学, 2001, 37(4): 124-128.
- Scientia Silvae Sinicae, 2001, 37(4): 124-128.
-
文章历史
- 收稿日期:1999-10-11
-
作者相关文章


森林土壤酶系主要来源于动植物的分泌及其残体的腐解和微生物的分泌等(关松荫, 1986), 可见, 植物种类和组成不同, 其根系分泌物和枯落物的质和量就不同, 从而可能引起土壤酶活性的差异, 但已有的报道主要涉及到土壤酶活性的测定(Sarathchander et al., 1984), 土壤酶性质的研究(Tate, 1984; Businan et al., 1985; Lajtha et al., 1985)以及不同森林类型的土壤酶活性的分布特征和动态及其与其它土壤理化性质和生态因子的关系研究等(杨万勤等, 1999a, 1999b; 张其水, 1992; 郑文教等, 1995; 张萍, 1995)。但有关土壤酶活性与植被的具体指标之间的关系研究尚未见报道。因此, 我们选择了植物多样性指数(Hill的多样性指数族NA和Shawnon-Wiener H′ index)作为植被指标, 初步研究了缙云山森林生态系统内三个次生演替群落内的土壤酶活性与植物多样性之间的关系, 试图揭示土壤酶与植被系统之间的关系, 以期为森林经营和管理提供科学依据。
1 研究地区的自然概况和样地资料缙云山位于重庆市北部约40km的北碚区境内, 位于29°49′N, 106°20′E, 属典型的亚热带季风性气候, 海拔900m左右, 年均降雨1143mm, 土壤为酸性黄壤。其环境特点及植被概况已有报道(钟章成, 1988)。样地资料见于表 1。
|
|
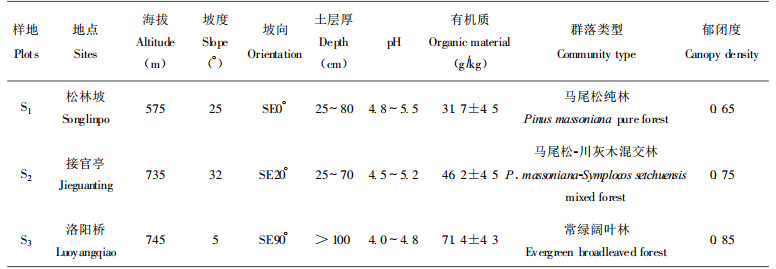
于1996年3月下旬、6月下旬、10月上旬和12月下旬, 分别在三个样地中以对角线方式设置5个样点, 分别按自然土壤剖面取样法采集腐殖质层(A)、淀积层(B)和母质层(C)中央的土样, 分别装于采集袋中, 带回实验室, 分出杂物, 风干, 磨碎, 过2mm筛, 分装于广口瓶中, 待测。在夏季采样时, 采用样方法进行群落调查(Cox, 1979)。
2.2 土样分析土壤pH采用pH计法; 有机质的测定采用重铬酸钾法(劳家柽, 1988)。土壤过氧化氢酶活性的测定:容量法; 蛋白酶活性的测定:茚三酮比色法; 转化酶活性的测定:3, 5-二硝基水杨酸比色法; 酸性磷酸酶活性的测定:磷酸苯二钠比色法(关松荫, 1986)。
2.3 数据处理相关分析应用SPCOVAR.BAS软件包, 植物多样性指数应用SPDIVERS.BAS软件包于IBM-PC586计算机上处理(Ludwig et al., 1990)。计算公式如下:

|
Pi—第i种个体(或胸面积盖度)所占比值; N0—物种数; N1—测定样本中丰富的物种数; N2—测定样本中“非常丰富”的物种数。

|
使用H′、N1和N2便于排除偶见种和稀有种, 即以物种数和每种个体数计算总物种多样性指数。
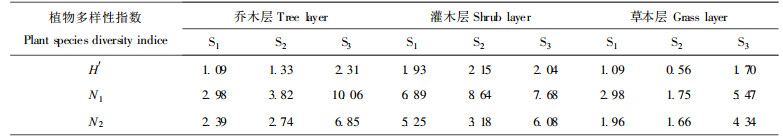
3 结果与分析 3.1 三个演替群落的植物多样指数特征研究表明(表 2), 乔木层的物种多样性指数H′、N1和N2都随演替方向升高, 即马尾松纯林(S1) < 马尾松-川灰木混交林(S2) < 常绿阔叶林(S3)。灌木层的多样性指数H′和N1不随演替方向升高, 以针阔混交林最高, 但如以“非常丰富”的物种数(N2)来计算, 则表现为S2 < S1 < S3, 以常绿阔叶林阶段最高。草本层总物种多样性指数H′、N1和N2以常绿阔叶林最高。以上结果表明, 用物种多样性指数, 尤其是用N2 (便于排除稀有种和偶见种)来反映各演替阶段的物种多样性变化, 以常绿阔叶林最高。
|
|
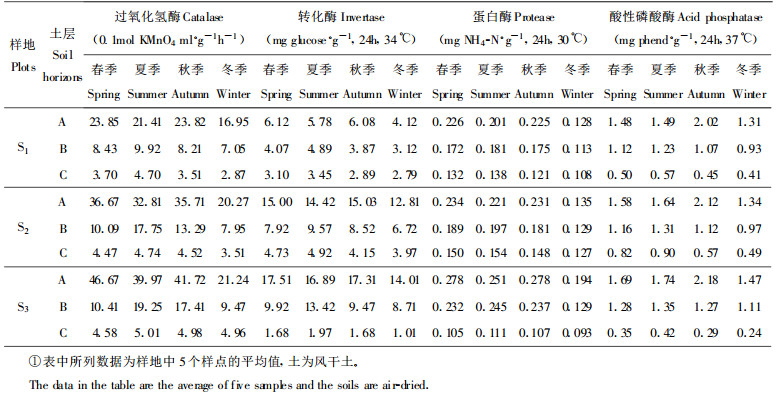
关于缙云山森林生态系统内土壤酶活性的分布特征和季节动态的研究, 作者已有报道(杨万勤等, 1999a, 1999b), 本项研究就不再作阐述, 现将研究结果列于表 3中, 以供参考。
|
|
相关分析表明, 土壤过氧化氢酶活性与植物多样性指数之间存在不同程度的正相关关系, 并且, 土壤过氧化氢酶活性与植物多样性指数H′、N1和N2之间的相关程度依A→B→C的顺序减弱, 植物多样性指数H′、N1和N2与A、B两层的过氧化氢酶活性之间的相关程度依乔木层→草本层→灌木层的顺序减弱, 与C层过氧化氢酶活性的相关程度依乔木层→灌木层→草本层的顺序减弱(表 4)。从表 4可知, 乔木层植物多样性指数H′、N1和N2与A层过氧化氢酶活性之间的相关程度均达到极显著水平(P < 0.01), 与B层过氧化氢酶活性之间的相关程度分别达到显著(P < 0.05)或极显著水平, 与C层过氧化氢酶活性相关, 但不显著。灌木层植物多样性指数H′、N1和N2与A、B、C三层的过氧化氢酶活性之间的相关程度均未达到显著性水平。草本层植物多样性指数与A层过氧化氢酶活性呈极显著正相关(P < 0.01), 与B、C层的相关程度均未达到显著水平。以上结果表明, 森林生态系统中, 植物的种类和数量对土壤过氧化氢酶活性具有重要作用, 其中, 乔木层植物的种类和数量对土壤过氧化氢酶活性的影响最为显著, 其次是草本植物的种类和数量, 并且这种作用随着土层的加深而减弱。
|
|
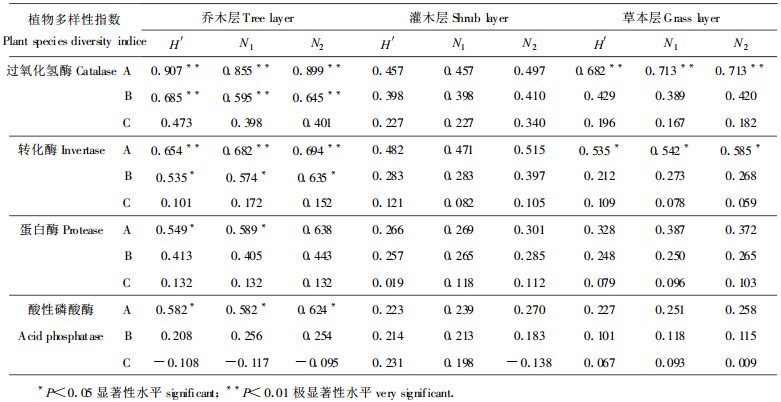
土壤转化酶活性与植物多样性指数之间存在不同程度的相关关系, 且相关程度依A→B→C的顺序减弱(表 4), 即土层愈深, 植物对其影响愈小, 这或许可以解释C层转化酶活性与随群落演替规律并无必然联系的现象(杨万勤等, 1999a)。由表 4可知, 乔木层植物多样性指数H′、N1和N2与A层转化酶活性均呈极显著正相关(P < 0.01), 与B层转化酶活性均呈显著正相关(P < 0.05), 与C层转化酶活性呈正相关, 但不显著。灌木层植物多样性指数与A、B、C三层的土转化酶活性的相关性均未达到显著水平。草本层植物多样性指数H′、N1和N2与A层转化酶活性均呈显著正相关(P < 0.05), 与B层转化酶活性相关, 但不显著, 与C层转化酶活性几乎不相关。以上结果表明, 植物的种类和数量对土壤转化酶活性, 尤其是对A、B两层转化酶活性具有重要作用, 其中, 乔木层植物的种类对A、B两层转化酶活性的作用最为明显, 草本层植物的种类和数量对A层转化酶活性的影响也很明显。灌木层植物的种类和数量对A、B两层的转化酶活性也有一定的作用, 但相对较弱。
3.4 土壤蛋白酶活性与植物多样性的关系相关分析表明(表 4), 土壤蛋白酶活性与植物多样性指数之间存在不同程度的正相关关系, 且相关程度依A→B→C的顺序减弱。从表 4可知, 只有A层蛋白酶活性与乔木层植物多样性指数H′、N1和N2之间的相关性达到显著性水平(P < 0.05), 其余的相关系数均未达到显著性水平。这表明, 乔木层植物的种类和数量与A层蛋白酶活性密切相关, 其对A层蛋白酶活性影响较大, 而灌木层和草本层植物的种类和数量对土壤蛋白酶活性有一定的影响和作用, 但相对较弱。
3.5 土壤酸性磷酸酶活性与植物多样性的关系由表 4可知, A, B两层的酸性磷酸酶活性与植物多样性指数H′、N1和N2之间存在不同程度的正相关关系, 但只有A层酸性磷酸酶活性与植物多样性指数之间的相关性达到显著水平(P < 0.05), 其余的相关系数均未达到显著水平。C层酸性磷酸酶活性与植物多样性指数的相关性很微弱, 甚至出现不显著的负相关。可见, 乔木层植物的种类和数量对A层酸性磷酸酶活性有显著影响, 而对B、C层酶活性的影响较弱。灌木层和草本层植物的种类和数量对土壤酸性磷酸酶活性的影响更微弱。
4 讨论在不同年限的次生林中, 土壤酶活性的变化在一定程度上反映出土壤和植被演替的规律, 演替年限越长, 土壤酶活性越高(张萍, 1995)。但杨万勤等(1999a;1999b)研究表明, 土壤酶活性的高低不仅与群落演替方向有关, 还可能与群落的物种组成有关。本项研究所选择的三个次生演替群落的土壤酶活性受海拔、温度、土壤类型(均为酸性黄壤)等的影响并不显著(杨万勤等, 1999a)。研究结果初步表明, 植物多样性指数H′、N1和N2与所研究的四种土壤酶活性之间存在不同程度的相关关系, 相关程度因土壤层次(或深度)、土壤酶类和植物的生活型(乔、灌、草)而异。总体而言, 植物多样性指数与土壤酶活性的相关程度依A→B→C的顺序减弱, 与土壤过氧化氢酶活性的相关性最强, 其次是与转化酶活性的相关性。乔木层植物多样性指数与A、B两层的过氧化氢酶活性、转化酶活性及A层的蛋白酶活性和酸性磷酸酶活性之间的相关程度均达到显著(P < 0.05)或极显著水平(P < 0.01), 草本层植物多样性指数与A层的过氧化氢酶活性和转化酶活性的相关性达到显著或极显著水平; 灌木层植物多样性指数与各个土壤层次的各种酶活性的相关均不显著。可见, 中亚热带地区的森林生态系统中, 乔木层植物的种类和数量对于森林土壤酶活性的影响最为显著, 其次是草本层植物的种类和数量, 灌木层植物的影响最弱。森林土壤酶活性是表征森林土壤肥力的重要参数之一(关松荫, 1986), 而森林土壤肥力是林业可持续发展的基础, 本项研究结果是否能为森林土壤培肥提供一条新的思路, 即森林土壤培肥的植物(尤其是乔木和草本植物)多样性, 这是有待探讨的问题。另一方面, 生物多样性保护措施中, 通过森林土壤培肥是否是一项有力措施, 也有待深入探讨。
关松荫. 1986. 土壤酶及其研究法. 北京: 农业出版社, 1-327.
|
劳家柽. 1988. 土壤农化分析手册. 北京: 农业出版社, 67-340.
|
杨万勤, 钟章成, 李瑞智等. 缙云山天然次生林的土壤酶活性的分布特征研究. 生态学研究论文集(董鸣、J. W. Werger主编). 重庆: 西南师范大学出版社, 1999a: 171~ 179
|
杨万勤, 钟章成, 韩玉萍. 1999b. 缙云山森林土壤酶活性的分布特征、季节动态及与四川大头茶的关系研究. 西南师范大学学报(自然科学版), 24(3): 318-324. |
张萍. 1995. 西双版纳次生林土壤微生物生态分布及其生化特性的研究. 生态学杂志, 14(1): 21-26. |
张其水. 1992. 福建杉木连载林地营造不同混交林后土壤酶活性的季节动态. 土壤学报, 29(1): 104-108. |
郑文教, 王良睦, 林鹏. 1995. 福建和溪亚热带雨林土壤酶活性研究. 生态学杂志, 14(6): 16-20. DOI:10.3321/j.issn:1000-4890.1995.06.006 |
钟章成. 1988. 常绿阔叶林生态学研究. 重庆: 西南师范大学出版社, 103-114.
|
Busman L M, Tabatabai M A. 1985. Factors effecting enzymic and nonenzymic hydrolysis of trimetaphosphate in soils. Soil Science, 140(4): 421-428. |
Cox G W(蒋有绪译). 普通生态学实验手册. 北京: 科学出版社, 1979: 229~ 299
|
Lajtha K, Bloomer S H. 1988. Factors affecting phosphate sorption and phosphate retention in a desert ecosystem. Soil Science, 146(3): 160-167. DOI:10.1097/00010694-198809000-00003 |
Ludwig J A等(李育中等译). 统计生态学———方法和计算入门. 呼和浩特: 内蒙古大学出版社, 1990: 54~ 66, 94~ 100
|
Sarathchander S U, Perrott K W. 1984. Assay of β-Glucosidase activity in soils. Soil Science, 138(1): 15-19. |
Stott D E, Dick W A, Tabatabai M A. 1985. Inhibition of pyrophosphatase activity in soils by trace elements. Soil Science, 139(2): 112-117. DOI:10.1097/00010694-198502000-00003 |
Tate R L. 1984. Function of protease and phosphatase activities in subsidence of Pahokee muck. Soil Science, 138(4): 271-278. DOI:10.1097/00010694-198410000-00003 |