 2001, Vol. 37
2001, Vol. 37文章信息
- 李建民.
- Li Jianmin.
- 马褂木地理遗传变异和优良种源选择
- GEOGRAPHIC GENETIC VARIATION AND PROVENANCE SELECTION OF LIRIODENDRON CHINENSE
- 林业科学, 2001, 37(4): 41-49.
- Scientia Silvae Sinicae, 2001, 37(4): 41-49.
-
文章历史
- 收稿日期:2000-12-15
-
作者相关文章


马褂木(Liriodendron chinense (Hemsl.) Sarg)自然分布于秦岭、淮河以南, 五岭以北, 横断山脉以东和东海以西11省(区), 包括陕西、安徽、浙江、江西、福建、湖南、湖北、贵州、四川、广西和云南等, 以武陵山、湘西南、桂西北及安徽、浙江.福建和江西等省为分布中心。马褂木适应性强、生长迅速、材质优异, 是优质的胶合板、高档家具和造纸等用材(顾万春, 1992; 叶桂艳, 1996), 是用材林基地建设优先发展的速生优质阔叶树种(李建民等, 2000)。因其花美叶异, 在江浙及其以北地区还是城镇绿化美化的好树种。
近10 a来各地多以开发利用马褂木观赏价值为培育目标, 侧重于育苗(肖水清等, 1994)、扦插繁殖(郭继善, 1995)和种间杂种优势利用叶金山1)、李周岐2)等研究, 较少根据不同培育目标定向选育。1992年世界银行贷款国家造林项目阔叶树研究课题组织开展了马褂木全分布区地理种源区域试验, 苗期结果显示马褂木具有丰富的地理遗传变异, 种源和种源内选择潜力很大(董纯等, 1996; 王启和等, 1997; 李习泉, 1996)。本文将利用设置在福建省邵武市的马褂木种源试验林8 a生测定材料, 首次系统研究马褂木生长、材质材性等主要经济性状在种源间的遗传变异及其地理分异模式, 并分别以纸浆材和胶合板材为培育目标, 为基地造林选择确定相应的优良种源。
1)叶金山.鹅掌楸杂种优势的生理遗传基础。南京林业大学博士学位论文,1998。
2)李周岐.鹅掌楸属种间杂种优势的研究.南京林业大学博士学位论文, 2000。
1 材料与方法 1.1 试材来源和试验设计马褂木种源试验林设置在福建省邵武市水北乡四都村, 林地前茬为马尾松(Pinus massoniana Lamb.)、杉木(Cunninghamia lanceolata (Lamb.) Hook)次生林, 海拔260~350 m, 坡度25°~33°。土壤为花岗岩风化发育的红壤, 立地条件在试验重复间差异较大(从较差至很好)。马褂木参试种源有15个(安徽黄山、安徽大别山、浙江松阳、江西庐山、江西黄冈山、湖北恩施、福建武夷山、湖南绥宁、湖南桑植、湖南浏阳、贵州松桃、贵州黎平、四川叙永、四川酉阳、云南嵩明), 另加5个北美鹅掌楸(L.tulipifera Linn.)种源和1个杂种F1代之后裔(来自中国林科院亚林所)。试验林按完全随机区组设计, 4次重复, 3列8行24株小区。1993年1月用1年生裸根苗造林, 株行距2 m×2 m, 块状整地, 穴规40 cm×40 cm×30 cm, 连续抚育4a至幼林郁闭。
1.2 试验林调查和数据采集在苗木定植当年底调查造林成活率, 第8个生长季末对种源试验林全面调查。基于每木胸径生长、枝下高、侧枝形成分叉干特性、树干通直度的观测调查, 在每试验小区内选择4株生长最佳植株调查测定树高、冠幅、分枝性状(1级活枝数、最大侧枝基径、长度和枝角)、下部3m段树干的圆满度(用3m高处直径与胸径之比表示)等指标, 并在4株最佳植株胸高处的上坡方位用6mm直径的生长锥取得由髓心至树皮的无疵木芯, 用以测定木材基本密度。
1.3 统计分析对于造林死亡率、植株枯死率和侧枝形成分叉干率3个性状, 以试验小区测定值为单元进行方差分析检验区组和种源效应, 对于其它性状则以小区内单株测定值为单元进行方差分析以验证区组、种源、种源×区组互作效应, 其中胸径、枝下高和树干通直度用全林调查数据参与统计, 而树高、冠幅、树干圆满度和木材基本密度4个指标则用最佳植株测定数据统计分析。因试验有缺株, 各性状的方差分析均采用SAS/STAT软件中的GLM程序。在方差分析时树干通直度经X-1/2数据转换。
本试验未涉及多点种源测定, 不能验证种源与地点互作。鉴于本测定点各试验重复的立地条件差异较大(4个重复的立地条件可构建一个从较差→一般→较好→很好的1维环境梯度), 可用不同重复的测定值简单估算说明不同种源的遗传稳定性。
通过建立种源性状均值与产地经纬度的线性和非线性回归方程以揭示马褂木主要经济性状的地理分异模式, 并从种群分布、生境片断化等角度阐述地理分异的成因。利用系统聚类分析将参试种源归类以划分种源区。本研究还以纸浆材和胶合板材为选育目标筛选马褂木优良种源, 纸浆材的选育侧重于木材产量及与纸浆性能有关的材性指标, 而胶合板材的选育则侧重于直径生长和干材形质等指标。
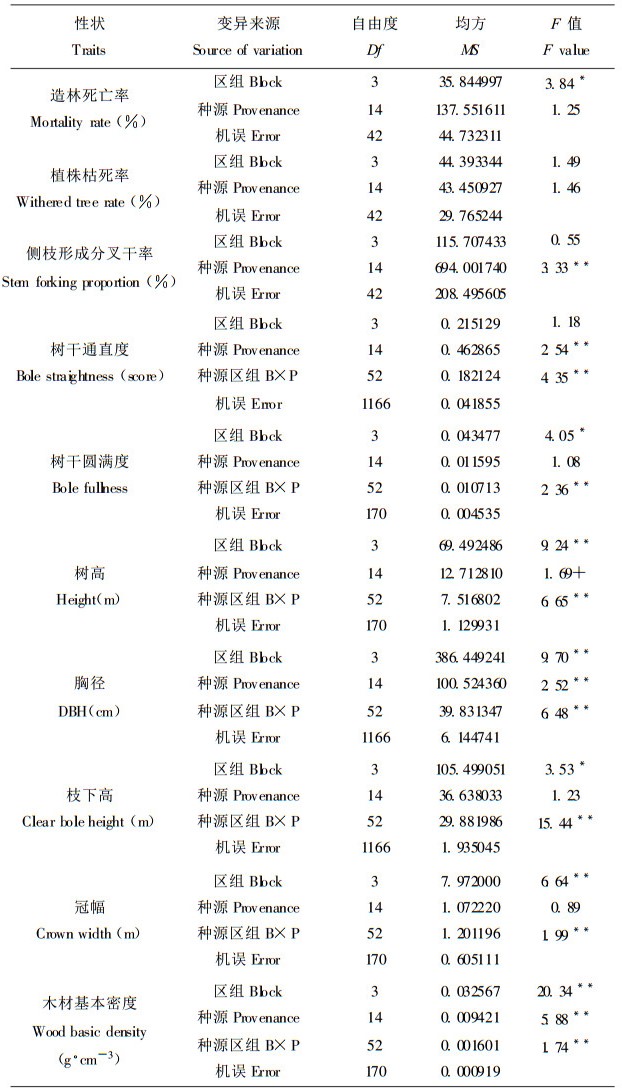
2 结果与分析 2.1 主要经济性状在种源间的遗传变异 2.1.1 造林死亡率和植株枯死率马褂木为落叶树种, 苗木根系欠发达, 但只要把握好造林季节, 当年生苗的造林成活率很高, 平均达到93.25 %。表 1方差分析结果显示马褂木造林死亡率在不同种源间差异不显著。与造林死亡率一样, 植株枯死率也是一个反应种源生态适应性的综合指标, 可说明生长过程中冻害、风折和竞争等环境压力对不同种源的作用。相对于北美鹅掌楸种源的引种造林(另文报道), 8 a生时马褂木不同种源的植株枯死率平均仅为3.31%。虽然马褂木造林死亡率和植株枯死率种源间的差异在统计学上不显著, 但有一定的变幅范围, 分别变化在1.04%~11.46%和0~9.42% (表 2)。
|
|
|
|
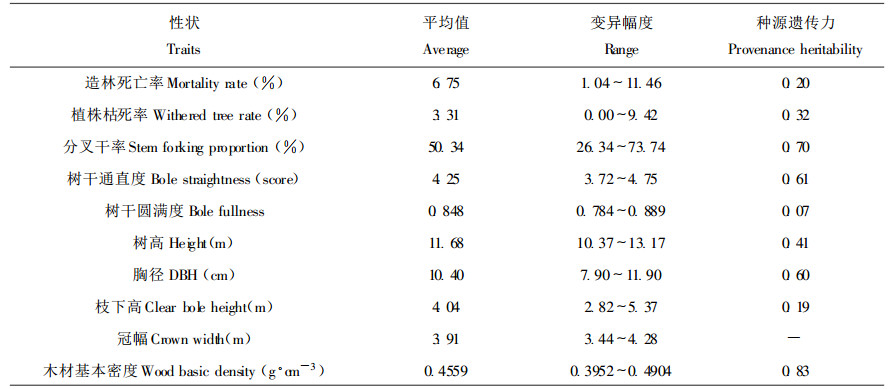
许多阔叶树和一些针叶树具有侧枝形成分叉干的特性, 原因于植苗当年树干基部侧芽异常生长之故。与其它树种相比, 马褂木这一特性尤其明显, 一般形成2~3个分叉干, 多的达5个分叉干。研究发现, 马褂木侧枝形成分叉干率受很强的遗传控制(种源广义遗传力为0.70), 在种源间差异极为显著(表 1), 种源选择时应引起重视。图 1的雷达图则具体展示了不同种源间侧枝形成分叉干率的显著差异性, 福建省武夷山种源的分叉干率为26.34%, 四川叙永种源的分叉干率为73.74%, 两种源相差2.8倍之巨。

|
图 1 马褂木不同种源侧枝形成分叉干率 Fig. 1 Stem forking proportion for different provenances 1.安徽黄山(Huangshan, AH); 2.安徽大别山(Dabieshan, AH); 3.浙江松阳(Songyang, ZJ); 4.江西庐山(lushan, JX); 5.江西黄冈山(Huanggangshan, JX); 6.湖北恩施(Enshi, HB); 7.福建武夷山(Wuyishan, FJ); 8.湖南绥宁(Suining, HN); 9.湖南桑植(Sangzhi, HN); 10.湖南浏阳(Liuyang, HN); 11.贵州松桃(Songtao, GZ); 12.贵州黎平(Liping, GZ); 13.四川叙永(Xuyong, SC); 14.四川酉阳(Youyang, SC); 15.云南嵩明(Songming, YN) |
马褂木是早期速生丰产型树种(李建民等, 2000), 但不同种源间高径生长差异很大。胸径速生型种源如贵州松桃、黎平, 浙江松阳、湖南浏阳和福建武夷山等8a生胸径都在11cm以上, 年生长量1.4cm左右, 而胸径慢生型种源如云南嵩明和四川酉阳的胸径仅分别为8.82cm和7.89cm, 年生长量只有1.0cm, 与速生型种源相差30%~40%。相对于胸径生长, 种源间树高生长差异相对较小, 仅在10%水平上显著, 其变幅范围在10.37~13.17m, 究其原因, 乃是由于利用小区内最佳生长株测定值参与统计分析之故, 但这不影响种源间树高生长的相对比较。对于冠幅生长, 试验未发现其显著的种源效应。
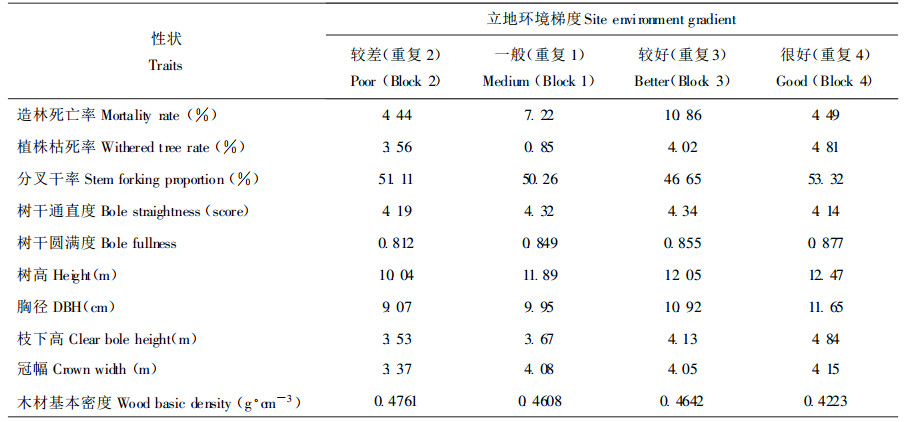
2.1.4 干形和木材基本密度树干通直度、圆满度和枝下高等干形指标是优选胶合板材的重要目标性状, 对于纸浆造纸材, 木材密度则是主要的育种目标性状。马褂木顶端优势明显, 树干较通直, 但不同种源的树干通直性差异很大(表 1、2)。从图 1、2的雷达图可看出, 有很多种源诸如福建武夷山、浙江松阳、贵州黎平、松桃、湖北恩施、四川叙永等树干非常通直, 得分都在4.40分以上(5分为最高得分), 而江西黄岗山、安徽大别山和云南嵩明3个种源树干较弯曲, 得分仅3.7~3.8分。方差分析揭示马褂木树干圆满度和枝下高两性状的种源效应在统计学上不显著。本研究发现所有马褂木参试种源树干下部3 m段都较为圆满, 平均圆满度为0.848, 其变幅较小(0.784~0.889)。枝下高的种源效应虽不显著, 但种源间的绝对差值较大, 最大值的四川叙永种源为5.40m, 最小值的四川酉阳种源为2.85m, 相差2.55m, 在胶合板材优良种源选择时要重视枝下高的这种种源差异性。木芯测定结果发现, 马褂木8a生木材基本密度中等(0.4559g·cm-3), 低于桦木而高于杨木、桤木(陈炳星等, 2000), 不同种源间木材基本密度差异极为显著, 并受较强的遗传控制, 最高值和最低值相差0.1g·cm-3左右, 这意味着木材密度的种源选择潜力巨大, 且易于遗传操纵。

|
图 2 马褂木不同种源的树干通直度 Fig. 2 Bole straightness for different provenances 1.安徽黄山(Huangshan, AH); 2.安徽大别山(Dabieshan, AH); 3.浙江松阳(Songyang, ZJ); 4.江西庐山(lushan, JX); 5.江西黄冈山(Huanggangshan, JX); 6.湖北恩施(Enshi, HB); 7.福建武夷山(Wuyishan, FJ); 8.湖南绥宁(Suining, HN); 9.湖南桑植(Sangzhi, HN); 10.湖南浏阳(Liuyang, HN); 11.贵州松桃(Songtao, GZ); 12.贵州黎平(Liping, GZ); 13.四川叙永(Xuyong, SC); 14.四川酉阳(Youyang, SC); 15.云南嵩明(Songming, YN) |
除植株枯死率、侧枝形成分叉干率和树干通直度3个指标外, 其它重要经济性状的立地环境(区组)效应都达到5%和1%的统计显著水平(表 1)。比较不同立地条件下的种源性状均值发现(表 3), 随着立地条件的改善, 马褂木树高、胸径和冠幅生长增加, 枝下高增长, 树干圆满度增大。鉴于这一变化规律, 在培育胶合板材时要求选用较好的立地条件, 以达到速生、树干圆满和长枝下高之培育目标。与高径等性状的变化规律相反, 木材基本密度则随着立地条件的改善而减小。立地环境梯度上造林死亡率差异显著, 但未发现其随立地环境条件变化的规律性。
|
|
统计分析表明, 马褂木生长、干形指标和木材基本密度具有显著的种源与立地环境(区组)互作效应(表 1), 在不同立地条件上应用优选的种源时要重视种源与立地环境互作和种源的遗传稳定性。表 4分别列出了胸径和木材基本密度两性状各参试种源的基因型值和Wricke's生态价, 比较发现, 各种源的遗传稳定性差异很大(详见表 4所注)。浙江松阳、贵州松桃和四川叙永为胸径速生稳定型种源, 在不同的立地上都能推广, 福建武夷山、湖南浏阳和贵州黎平则是胸径速生不稳定型种源, 适宜在条件较好立地上选用; 在上述6个胸径速生型种源中, 福建武夷山属高密度稳定型种源, 贵州黎平为高密度中等稳定型种源, 浙江松阳和湖南浏阳分别属于中等密度中等稳定型种源和中密度不稳定型种源, 而贵州松桃和四川叙永为低密度不稳定型种源。
|
|
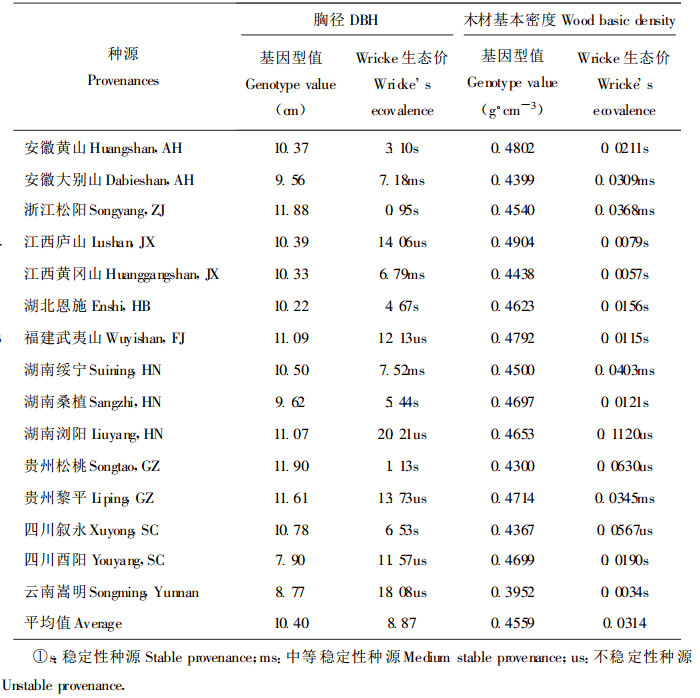
表 5的相关系数显示, 马褂木多数经济性状与产地经纬度的相关系数不显著, 不存在典型的地理变异模式。造林死亡率和枝下高两性状虽分别与产地的经度和经纬度有显著的相关性, 但因其种源效应较小而实际意义不大。分析发现木材基本密度受产地径度控制, 呈明显的经向变异模式, 随着产地经度的增加其木材基本密度有增大之势。二维反应面(Response surface)分析可形象地揭示性状在纬度和经度二维平面上的变化趋势, 从分析结果来看(表 5), 只有造林死亡率和植株枯死率两性状的回归方程显著, 拟合优度较高, 但同样因其种源效应不显著而意义不大。
|
|
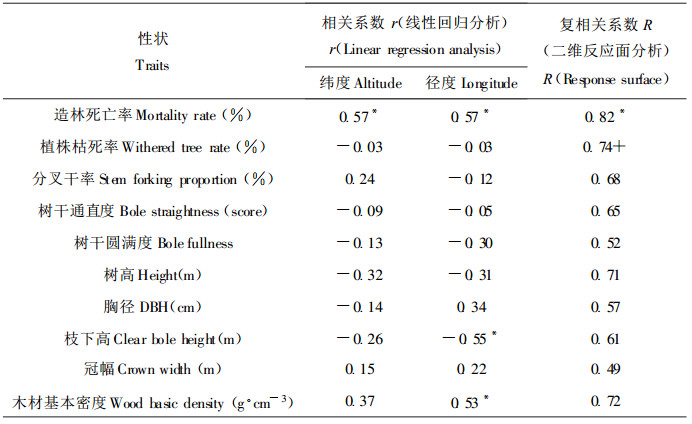
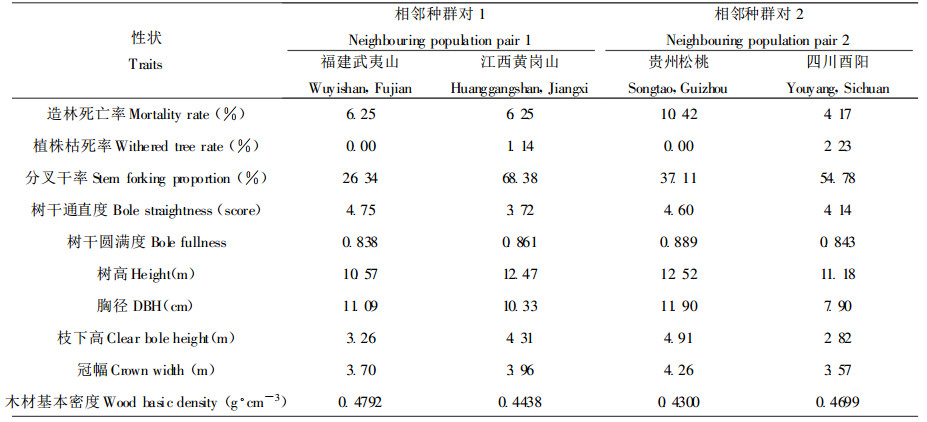
马褂木地理分异模式的成因, 可从其种群分布等角度解释。与种群大、呈连续分布的北美鹅掌楸迥然不同是, 马褂木星散间断分布于海拔600~1800 m的山坡或沟谷。除西部种群外, 多数种群的个体数较小, 在1~20株之间(郝日明等, 1995)。这种生境片断化、小种群效应(遗传漂变、近交等)和距离隔离效应的结果, 将使得马褂木种群内的遗传变异程度降低, 而种群间的遗传分化程度加大(陈小勇, 2000)。这里发现即使相邻的两个马褂木种群其遗传分化程度也很高。例如, 参试的福建武夷山和江西黄冈山2个种群分别位于武夷山脉的东西两侧, 相距仅数十公里, 但差异非常显著, 两者8 a生胸径分别为11.09 cm和10.33 cm, 侧枝形成分叉干率分别为26.34%和68.38%, 相差2.6倍, 木材基本密度分别为0.4792 g.cm-3和0.4438 g·cm-3, 相差0.035 g·cm-3。同样, 相距甚近的贵州松桃和四川酉阳两种群间差异更大(表 6)。因小种群效应和距离隔离效应等, 除木材基本密度等少数性状外, 马褂木多数经济性状无明显的地理变异规律, 而呈随机分布。
|
|
采用主要经济性状的种源均值, 数据经标准化转换后利用欧氏距离进行聚类分析。对比分析显示, 系统聚类中各种方法的聚类效果相似, 这里仅给出了利用可变类平均法的聚类结果。根据图 3的聚类系谱图可大致按地理区域将参试种源划分成2个聚类大组。第1聚类大组主要包括马褂木自然分布区的西南部种源, 如贵州松桃、黎平、四川叙永和云南嵩明。有趣的是, 浙江松阳和江西黄冈山这2个种源表现较为特别, 与西南部种源相近, 与中、东部种源差异较大。第1聚类大组可细分为2个聚类亚组, 分别包括贵州松桃、黎平种源和四川叙永、云南嵩明种源。第2聚类大组包括马褂木中部和东部的大部分种源, 也可再分为2个聚类亚组, 1个聚类亚组从西至东包括湖北恩施、湖南桑植、浏阳和福建武夷山种源, 这4个种源大致处在相同的纬度范围内; 另1个聚类亚组包括自然分布区东北部的安徽大别山、黄山、江西庐山种源和中部的湖南绥宁和四川酉阳种源。从聚类结果来看, 在各聚类大组和聚类亚组有些种源在地域上不相连, 这样仅能粗略地而不能精确地进行种源区的划分, 但这也有力地证实了小种群效应和距离隔离效应对马褂木种群遗传分化的作用, 清楚地说明了马褂木地理变异呈随机分布的特性。

|
图 3 马褂木15个参试种源的聚类系谱图 Fig. 3 Dendrogram of 15 seed sources tested of Liriodendron chinense |
不同材种的育种目标是不同的。从马褂木基本密度变化值(0.3952~0.4904 g·cm-3)来看, 其木材硬度适中, 作为纸浆用材可用以发展CTMP和BCTMP等浆种。纸浆材优良种源的选择要求速生丰产, 单位时间单位面积纸浆收获量最大, 这里主要以干物质指数作为选择标准, 可以选择确定浙江松阳、贵州黎平和松桃为3个纸浆材优良种源(表 7), 其干物质指数较当地福建武夷山种源高20%以上。在所有参试种源中, 福建武夷山种源的遗传值相对较高, 因此还选用种源群体均值作为参照, 这样3个入选种源的干物质指数增益将大于30%。
|
|
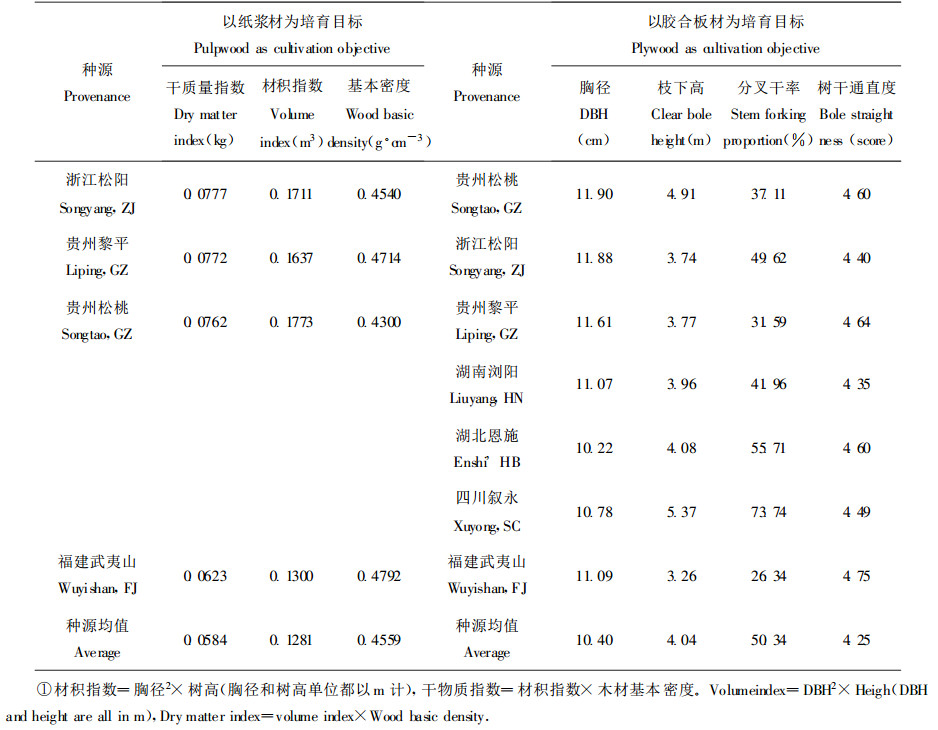
对于胶合板材优良种源的选择, 要求径生长量大、枝下高长、树干通直圆满、侧枝形成分叉干率低。由于树干圆满度在种源间的绝对差值很小, 种源优选时可不考虑这一干形指标。侧枝形成分叉干虽在一定程度上能提高单位面积的生物产量, 但会严重影响干材的径生长、降低干材培育的等级。同样以当地福建武夷山种源和群体均值作参照, 比较发现贵州松桃种源表现最佳; 四川叙永种源胸径生长迅速、枝下高最长、树干通直, 但侧枝形成分叉干率较高, 若作为胶合板材种源选用时应辅以人工抹芽和去萌; 此外, 在种子来源缺乏时, 诸如浙江江松阳、贵州黎平、湖南浏阳和湖北恩施等种源也可选用。
3 结论与讨论鹅掌楸属树种是古老的孑遗植物, 在日本、格陵兰、意大利和法国等曾发现它的化石。在第三纪时本属尚有十余种, 广布于北半球温带地区, 但经第四纪冰川的摧残, 现仅存马褂木和北美鹅掌楸2种(王诗云等, 1995)。种源试验发现, 北美鹅掌楸生长、形态和生态适应性等性状从北至南呈典型的倾群变异模式, 分布区最南端的种源生长最快(Parks et al., 1994; Smith et al., 1988; Roeder, 1980)。根据Parks等(1994)对北美鹅掌楸50个种群体23个位点同工酶的研究结果, 可以很容易地将所有种群从地理上划区归类。
对照北美鹅掌楸, 马褂木虽然同样分布广泛, 但呈间断星散分布。按照郝日明等(1995)对其自然分布区的研究, 马褂木在地理上呈“一带五岛”分布形式。马褂木这种生境的片断化应主要归因于第四纪冰川的作用, 加之马褂木物种本身特有的致濒机制以及长期以来人为的选择压力, 现有的多数种群很小。本文利用马褂木全分布种源试验林8 a生测定材料, 首次系统研究了生长、干形和木材基本密度等在种源间的遗传差异及地理分异模式, 发现多数经济性状的种源效应非常显著, 未发现典型的倾群变异模式, 除木材基本密度受产地经度控制外, 其它性状多呈随机分布。根据15个种源的系统聚类谱图可看出, 福建武夷山和江西黄冈山、贵州松桃和四川酉阳两相邻种群的遗传距离较大, 分别归并在两个大类中, 这充分说明了小种群效应和距离隔离效应等对马褂木种群遗传分化的巨大作用。本文根据生长、形质等性状的研究结果似与朱晓琴等(1995)和罗佐光等(2000)利用分子标记的研究报道不同, 后两者分别利用等位酶和RAPD分子标记来研究马褂木的遗传多样性, 认为马褂木的变异主要来源于种群内, 而种源间的遗传多样性较小。
马褂木对立地环境条件的反应很敏感, 在较好的立地上, 马褂木树高、胸径和冠幅生长快、枝下高长、树干圆满, 但木材基本密度略低, 这种环境效应为不同材种培育在立地选择上提供了原则依据。本文通过系统聚类可将马褂木参试种源粗略地划分成2个聚类大组和4个聚类亚组, 但由于有些种源表现特异, 虽归在某一类中, 但与类内其它种源在地域上是不相连的, 因此利用系统聚类难以精确进行种源区的划分。按照纸浆材和胶合板材2种培育目标, 分别选择确定了相应的优良种源。从优良种源分布情况来看, 贵州松桃、黎平和四川叙永相对集中, 这应是一个优良种源区, 但其它入选优良种源的分布则较为分散, 无一定的规律。
陈炳星, 李光荣, 周志春, 等. 2000. 引种四川桤木木材化学组分的分析与评价. 中国造纸, 19(4): 20-23. DOI:10.3969/j.issn.0254-508X.2000.04.004 |
陈小勇. 2000. 生境片断化对植物种群遗传结构的影响及植物遗传多样性保护. 生态学报, 20(5): 884-892. DOI:10.3321/j.issn:1000-0933.2000.05.027 |
董纯, 谭德仁, 汪长江, 等. 1996. 马褂木地理种源试验苗期及幼林生长测定报告. 湖北林业科技, (2): 6-15. |
郭继善. 1995. 关于杂种马褂木的扦插繁殖. 林业科技开发, (2): 2-3. |
顾万春. 1992. 主要阔叶树速生丰产培育技术. 北京: 中国科学技术出版社.
|
郝日明, 贺善安, 汤诗杰, 等. 1995. 鹅掌楸在中国的自然分布及其特点. 植物资源与环境, 4(1): 1-6. |
李建民, 封剑文, 谢芳, 等. 2000. 鹅掌楸人工林的丰产特性. 林业科学研究, 13(6): 622-627. DOI:10.3321/j.issn:1001-1498.2000.06.008 |
李习泉. 1997. 马褂木属地理种源试验苗期初步研究. 湖南林业科技, 24(1): 4-7. |
罗佐光, 施季森, 尹佟明, 等. 2000. 利用RAPD标记分析北美鹅掌楸与鹅掌楸种间遗传多样性. 植物资源与环境学报, 9(2): 9-13. DOI:10.3969/j.issn.1674-7895.2000.02.003 |
王启和, 干少雄, 龚毅红, 等. 1997. 鹅掌楸种源造林试验初报. 四川林业科技, 18(1): 14-17. |
王诗云, 赵子恩, 彭辅松, 等. 1995. 华中珍稀濒危植物及其保存. 北京: 科学出版社.
|
肖水清, 廖龙泉. 1994. 五种优良阔叶树育苗技术及苗木生长规律初报. 江西林业科技, (1): 16-18. |
叶桂艳. 1996. 中国木兰科树种. 北京: 中国农业出版社.
|
朱晓琴, 马建霞, 姚青菊, 等. 1995. 鹅掌楸遗传多样性的等位酶论证. 植物资源与环境, 4(3): 9-14. |
Parks C R, Wendel J F, Sewell M M, et al. 1994. The significance of allozyme variation and introgression in the Liriodendron tulipifera complex (Magnoliaceae). American Journal of Botany, 81(7): 878-889. DOI:10.1002/j.1537-2197.1994.tb15569.x |
Roeder K R. 1980. Provenance performance of Liriodendron tulipifera in South Africa. South African Forestry Journal, (112): 42-44. |
Smith A L, Campbell C L, Walker D B, et al. 1988. Geographic variation in the essential oil monoterpenes of Liriodendron tulipifera L.. Biochemical Systematics and Ecology, 16(7-8): 627-630. DOI:10.1016/0305-1978(88)90075-0 |