 2001, Vol. 37
2001, Vol. 37文章信息
- 张云跃, 马常耕, 林睦就, 李柏海.
- Zhang Yunyue, Ma Changgeng, Lin Mujiu, Li Baihai.
- 我国银杏遗传变异研究之一———种核性状的群体间和群体内变异
- STUDY ON ONE OF GENETIC VARIATIONS FOR GINKGO BILOBA IN CHINA THE VARIATION OF BREEDING FRUIT-STONE CHARACTERS AMONG AND WITHIN POPULATION
- 林业科学, 2001, 37(4): 35-40.
- Scientia Silvae Sinicae, 2001, 37(4): 35-40.
-
文章历史
- 收稿日期:1999-04-21
-
作者相关文章


2. 中国林业科学研究院林业研究所 北京 100091;
3. 湖南省林业科技推广总站 长沙 410007
2. The Research Institute of Forestry, CAF Beijing 100091;
3. General Station of Forestry Science and Technology Extension, Hunan Province Changsha 410007
银杏(Ginkgo biloba)是我国仅有的著名孑遗树种之一, 由于其多用途特性, 现在已被作为一个多目的树种受到国内外的重视, 集约栽培正在发展中。做为集约栽培基本要素之一的遗传改良工作的我国也已开始。为提高遗传改良效率必须对各地银杏群体遗传结构和性状变异特点有清晰的了解。为此, 我们从种用、叶用和材用方面开展了种核形态学、营养成分、叶中化合物含量和叶片大小、重量、生态适应性、生产力的产区、家系和个体遗传变异研究。本文是这一系列研究结果之一, 目的是揭示种核形态变异规律, 以有助于解决当前我国银杏良种选育中存在的不从选种工作尚处原始群体起步的初始阶段, 应突出无性系品种选育, 却热衷于根据果实和种核形态划分品种(群), 妨碍科学品种选育正常开展的问题。因为品种是人工选择的结果, 选育要突出经济目的, 而不是自然群体诸多特征、特性的简单分群。从适应性角度看, 不同地区相同形态的个体也不能合称一个品种。在开展这一研究中, 我们也充分认识到由于现有银杏的孑遗性, 群体的遗传结构受自然残遗过程和人为活动的重大干扰, 为遗传变异模式的研究带来的不少负面影响, 并力求避免它们。如它的现代分布(生长)空间虽十分辽阔, 大约在北纬22°~41°31′和东经98°~121°30′ (许慕农等, 1993)内的范围, 可是自然群落和天然更新幼龄植株却相当罕见, 而是多数地区以古树星散分布和少数地区集中栽培的独特分布模式。遗传漂变在各地现有群体形成中的作用不可忽视。加之雄株极少, 性比例失调造成基因交流的局限性和不平衡性, 必然造成背离随机交配群体中正常异株授粉植物所普遍表现出的遗传结构模式。考虑到银杏多生长在低海拔农作区域, 所以我们基本以中国自然区划概要(席承藩等, 1984)一书中农业气候区划方案为依据, 对各采种母树进行区域分类和统计处理, 以使研究结果尽量有生态区代表性和选种的实用性。
1 试验材料和研究方法 1.1 收集原则和代表生态区本着力求母树有代表性原则, 采种做到星散分布和集中产区兼顾, 每一集中产区随机由10~15株有广大空间距离的树上采种, 星散分布地区则3~10株不等。为使试样有尽多的原始遗传代表性, 尽可能从100a生以上的实生大树上采种, 为使种核有充分典型性, 采种母株既要大量结实又要果实达充分成熟。采种范围包括15个省(区), 但由于一些地点母株不足5株, 未纳入统计。各气候区包括的采种点和统计分析株数见表 1。考虑到江苏南部为一典型的人工栽培区, 并长期用嫁接方式繁殖而与各实生繁殖区不同, 故独立为一个统计单位, 以资比较。
|
|

研究性状有: (1)种核长度; (2)种核宽度; (3)单核重量; (4)核壳厚度; (5)出仁率。为研究核的形状计算了核形指数(核长/核宽)。
每母株从总收获种核堆中随机抽取2 kg种核, 再按林木种子检验标准取样, 每20粒1组测量各性状, 取4组平均值做为各母株代表值进行各研究性状的统计分析。
1.3 统计参数由于本研究重点是揭示性状的变异强弱和格局, 故突出变异系数的分析, 以每一母株的代表值计算各采种点(群体)的平均表现和变异系数, 然后把同一气候区各采种母树的测得值统一计算出所代表气候区的平均值和变异系数。为探讨选种的综合效应, 还计算了各研究性状间的相关系数。
2 结果与分析 2.1 种核性状的气候区间差异和变异总趋势为比较显示各气候区种核性状的异同, 表 2给出了统计结果。从表 2的数值看到: (1)不同气候区银杏种核的特征除单核重量和核长度外, 其它性状虽有所不同, 但差异不大, 特别是出仁率的地区差异更小。(2)性状的遗传差异无明显地理规律性, 相反却看到明显人为干扰的痕迹, 如2和6区一个是北方主要栽培区, 一个是南方主要栽培区, 地理纬度差近10°, 但单核重却相近, 是分布区内两个单核最重的产区。3a和3b同属一个农业气候区, 虽地理距离较大, 由于都是长期实生繁殖区, 各性状基本一致, 保持着自然群体的遗传结构。3b和7区地理位置十分接近, 但后者由于栽培程度高, 且以嫁接繁殖为主, 人工选择造成二者之间的明显遗传差异。(3)栽培程度高的地区, 核长而重, 非栽培区的则更接近原始群体的性质。因此, 可以认为银杏种核性状的随机变异模式是自然群体残遗过程中遗传随机漂迁和人工选择的结果。对把果实形态这种非适应性性状作为选择目标时, 种源差异就不象以适应性性状为选种目标的用材树种那样有意义, 利用地理区间的整体遗传差异的潜力不如同为广域分布树种的马尾松、白榆那样有效, 而应以基因型选择为根本, 尽管从整体上看有南岭山区银杏因核整体平均重高属大核品种潜在选种区域。另外, 从现有商品性角度看, 江苏南部栽培区种核确有最高商品价值, 表现为核长、大和出仁率高的特点。
|
|
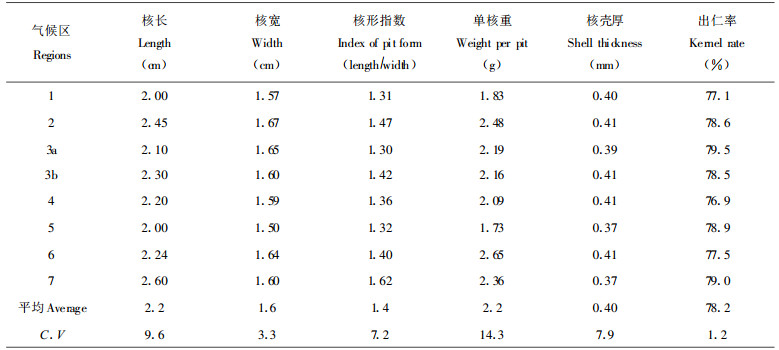
由于种群在残遗过程的偶然因素和人工干扰程度的(有意识和无意识选择)不确定性强烈影响着银杏性状的地理变异现有模式, 就必然存在同一气候区内群体间的遗传差异和基因库价值的不等值性。为证实此点, 把有3对代表性相邻群体的测定值列于表 3中。
|
|

表 3列举地理距离相近又同属一个气候区的3对可比较群体, 它们的种核性状都表现出与气候区间变异强度相近的变量, 如处南岭山地越城岭南北的广西兴安群体和湖南新宁群体遗传差异就极为明显, 前者核长而大, 壳较薄, 出仁率较高; 后者核小, 壳厚, 出仁率比广西兴安的低2.3%。从变异系数(表示性状遗传变异幅度大小)也可看出相邻群体由于自然和人工干扰作用的不同, 各性状的异质度也不尽相同, 预示在不同群体中改良相同性状可能有不同的潜力。如浙江临安和富阳两群体单核重的变异系数为12.8%和7.6%, 而且临安的单核重群体平均值也高于富阳群体, 可以断定通过选择从临安群体获得核重改良的机遇将比从富阳群体选择为高。综合表 2和表 3的结果, 认为群体的选择比气候区间选择更有实用价值。
2.3 个体的遗传变异前两项分析表明, 银杏种核性状在气候区间虽有一定差异, 但无地理规律性, 因同一气候区内相邻群体各性状的平均表现可有很大不同。果用植物选种需强调产品的一致性, 因它有利于加工和提高商品价值, 这只有以个体(基因型)为基础的选择, 推广无性系品种才能达到。所以研究个体的性状变异是提高银杏核用选种效率的根本。表 4给出了各气候区及全分布区范围内核性状个体间变异的特点, 以有助于识别各地区和总体的遗传资源价值和选种潜力。
|
|

从表 4数值看出, 在全分布区范围, 银杏核性状的遗传变异幅度是各不相同的, 这预示着通过选择改良核特征的可能性或难易程度因性状而异。变异系数所揭示的各性状变异幅度由大到小依次为单核重 > 壳厚度 > 果形 > 核长 > 核宽 > 出仁率。不同气候区间各性状的变异系数的不同预示着在不同气候区对同一性状选择其效率也不相同。从总体上看, 通过选择提高单核重最易成功, 而且在3a和6区内选择会更易于达到目的, 因为不仅这两区的核重变异系数最大, 而且单核最重的个体也出现在那里。出仁率是变异幅度最小的性状, 这与一些学者的研究结果一致(邢世岩等, 1994; 刘燕君, 1992), 表明通过现有个体选择要大幅度提高出仁率是困难的, 因此各地不必花大力气于这一性状的改良上, 通过提高单株产量达到提高总仁量比提高出仁率更为可行。
不同气候区间性状的绝对值和变异幅度不同, 说明我国各地银杏基因库在质与量上的差异及育种上的不等值性。一般平均表现好的区域或群体预示选种有利基因的频率高, 因而育种价值也高。如南岭山地的广西灵川和兴安最具选出大核品种的潜力, 江苏南部群体则较易选出长核形个体。考虑到单一性状最突出的个体不必然出现在平均表现最高的群体中, 如3a区平均单核重为2.9 g, 但却出现了所测定200余株母树中单核最重的个体; 南岭山地群体平均出仁率偏低(77.5%), 但却出现了出仁率最高(82.8%)的个体。这告诉我们在制定银杏改良计划时, 既要考虑群体的平均表现, 又不排斥在平均表现不高的地区(群体)作独特基因型的选择。由于现有银杏残遗历史过程中的基因随机漂迁, 从基因资源角度考虑, 不可忽视星散残遗个体的遗传考察。长期以来以嫁接方式为主的银杏栽培区, 核性状的总体表现虽最优异, 但本研究中所发现的性状极端个体却不总出现在其中, 故其育种价值并不高于实生繁殖区。
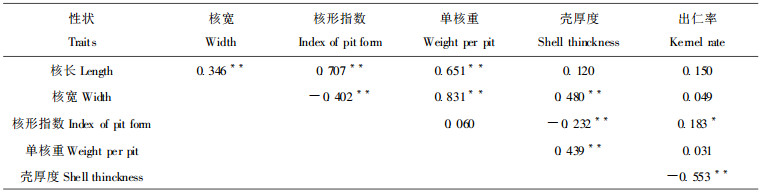
2.4 种核性状间的相关关系选种目的性状间的关系对制定选种计划有重大影响, 如性状间存在正向连应, 通过一个性状的选择可同时改良其它性状, 如确知两性状间存在负向连应, 就应采取能把这种负效应减至最小程度的选择方法。
从表 5看到, 核长极显著地正向影响着核形和核重, 对出仁率也有一定的正向作用。核宽虽也极正向影响着单核重, 却对壳厚度有不利影响, 而壳厚又显著对出仁率有不利影响。所以把长核形作为选择标志之一时, 预示可能会同时选出利于加工处理的核大和出仁率高的基因型。选核宽而近似圆形的核很可能会降低出仁率。这大概就是目前银杏栽培区的优良基因型, 多为称作佛指或马铃型而较少圆形的原因(郭善基, 1993)。总之, 在单核重相等条件下, 选择长核形果似更有经济意义些。
|
|
银杏核用选种的总目标虽是单位面积的总经济产量, 株形、结实特征和核出仁率等有其决定意义, 但核形和大小也影响着产品的商品价格, 所以把核的特性作为选种目的性状之一也是必要的, 这也是本研究的实用性效应。
3 结论我国银杏核性状在个体上存在明显的遗传差异, 但气候区间的平均表现除单核重外差异不明显, 且地区内相邻群体间的变异甚至大于地区间的, 因而种核性状变异没有地理规律性, 这可能是历史残遗过程中遗传随机漂迁的结果。因而以核性状为选择目标时, 研究重点应放到广泛的基因型调查上, 群体和地区表现只有参考意义。
银杏不同性状的遗传变异强度不同, 在所研究的性状中以单核重的变异水平最高, 预示其改良潜力最大, 出仁率是遗传差异最小的性状, 把它做为选种性状的意义不大。
不同气候区现存银杏的种质资源价值并不完全相同, 且群体平均表现和极端基因型的出现并不必然相关, 所以今后的遗传改良要在全分布区内广泛开展, 不应局限于少数栽培区。从未来杂交育种需要考虑, 具极端性状基因型的搜集比现有综合性状表现优良基因型选择更为重要。
由于核长与核重和出仁率有正的相关, 与壳厚相关不显著, 核宽却与壳厚极显著正相关, 从而影响出仁率, 所以选长核基因型比圆核基因型经济上更为有利些。
郭善基主编.中国果树志银杏卷, 北京: 中国林业出版社, 1993
|
刘燕君.湖北省安陆市银杏品种资源调查及良种选择初报.全国银杏首届学术研讨会论文集, 武汉: 湖北科技出版社, 1992
|
席承藩, 丘宝剑等主编.中国自然区划概要.北京: 科学出版社, 1984
|
邢世岩, 孙露.泰山银杏实生优良单株研究.全国第二次银杏学术研讨会论文集, 武汉: 湖北科技出版社, 1994
|
许慕农, 胡大维主编.银杏栽培和产品加工技术.北京: 中国林业出版社, 1993
|