 2001, Vol. 37
2001, Vol. 37文章信息
- 包维楷, 刘照光, 刘朝禄, 袁亚夫, 刘仁东.
- Bao Weikai, Liu Zhaoguang, Liu Chaolu, Yuan Yafu, Liu Rendong.
- 亚热带次生常绿阔叶林主要乔木种群自然恢复15年来的变化
- FIFTEEN-YEAR CHANGES OF TREE POPULATIONS IN THE SECONDARY SUBTROPICAL EVERGREEN BROAD-LEAVED FORESTS IN SOUTHWESTERN CHINA
- 林业科学, 2001, 37(1): 7-14.
- Scientia Silvae Sinicae, 2001, 37(1): 7-14.
-
文章历史
- 收稿日期:1999-05-12
-
作者相关文章


2. 四川省瓦屋山国家森林公园 洪雅 614374
2. Wawushan National Forest Park, Sichuan Province Hongya 612364
由于人类干扰活动的长期破坏, 目前我国常绿阔叶林分布面积已不足我国亚热带地区面积的5%(陈伟烈等, 1995)。而据卫片分析, 而面积较集中的常绿阔叶林不超过长江流域面积的0.22%(A K Myint et al., 1998)。常绿阔叶林的迅速恢复和重建已成为亚热带地区环境治理和生物多样性保护和维持的关键措施之一。自然更新或人工促进更新是常绿阔叶林恢复和重建与生物多样性维持的有效途径(刘金林等, 1984; 刘照光, 1994; 贺金生等, 1995;余作岳等, 1995; 彭少麟等, 1995; 刘照光等, 1997; 张水松等, 1997)。探讨常绿阔叶林自然恢复过程中的变化, 可能是目前寻找迅速的人工恢复和重建相对稳定的常绿阔叶林的理论和实践依据的最重要的途径之一, 对目前大面积低效纯林改造与人工恢复和重建森林也有积极意义。本文报告瓦屋山中亚热带次生湿性常绿阔叶林自然恢复过程的定位调查结果, 目的在于揭示乔木种群在相互作用下的动态变化特点, 了解各种群在群落恢复过程中的生长行为表现的差异性, 为相对稳定的常绿阔叶林人工恢复和重建提供树种选择的理论和实践依据。
1 材料和方法 1.1 研究对象及基本特点本研究在我国西部的中亚热带湿性常绿阔叶林区代表地段四川洪雅县南部的瓦屋山国家森林公园(前身是洪雅林场)内进行。位于102°51′~103°20′E, 29°31′~29°51′N, 公园面积476 km2(赵崇儒, 1991)。研究样地设置在中山区的木姜杠林班。该区在中国植被分区中该区属亚热带常绿阔叶林区域、中亚热带常绿阔叶林亚区域、中亚热带常绿阔叶林南部亚地带的川滇黔山丘栲类(Castanopsis)、木荷(Schima)林区(中国植被编辑委员会, 1987)。因此, 扁刺栲(Castanopsis platyacantha)、华木荷(Schima sinense)林是该区地带性的植物群落类型之一。共设置相邻的样地3块, 相距不足100 m远, 面积均为20 m × 20 m, 基本概况如表 1所示, 其环境条件基本一致。均为栲、木荷为主的常绿阔叶林1), 在1956年砍伐乔木层而遭到破坏后, 通过封山而自然恢复的次生林, 目前林龄为40~50 a。40多年来, 该区基本无人为干扰(1989年小范围内进行过小径木的择伐, 以取得耳木)。该地段属中亚热带季风性湿润气候, 温和湿润, 雨水多、云雾多、日照短, 湿度大。年均温10 ℃~14 ℃, 年日照不足800 h, 年降水量达2397.7 mm, 降水日达199~226 d, 年均相对湿度达85%~90%。
|
|

1) 刘国祥(洪雅林场).1987, 在实践中对次生林效益的认识.
1.2 研究方法对各固定样地上受破坏后恢复起来的该群落乔木层进行了4次(1982-04, 1983-12, 1989-07, 1997-06)调查。调查指标有树高3 m或以上的活立木种类、每木胸径DBH、树高H等。根据测定结果, 统计各乔木种群的密度、平均胸径、平均树高。乔木材积蓄积量与其生物量生产力成正比已经被国内外研究证实, 并已经成功用于生物量生产力的估算(Browns et al., 1984; 方精云等, 1996)。本文未进行生物量测定, 这里利用该区前期建立的各主要树种的一元材积表及其公式1), 计算出各林木的材积。以乔木材积总和代替各乔木种群生物生产力进行分析, 称材积生产力。
1) 四川洪雅县林业局和四川省第一森林勘察大队.1986.四川洪雅县林业区划、林业规划报告.
2 结果和分析 2.1 群落乔木层种群组成与地位及其变化3个固定样地乔木层(高于3 m)最近15 a共有22个种群存在(包括出现或消失的种)。在样地1中, 调查初期的几年存在的刺楸(Acanthapana × chinensis)、稠李(Prunus brachypode)、杜鹃(Rhododerdron sp.)和虎皮楠, 在1989年从群落中消失了, 随后分别出现了峨眉泡花树(Meliosma yunnanensis var. fisheriana)和大叶冬青, 但在1997年时也消失了, 而仅出现峨眉木姜子(Lindera fabri); 在样地2中, 调查初期的几年存在的绿叶石栎(Lithocarpus hancei)、小叶青冈(Cyclobalanopsis myrsinaefolia)、杜鹃、虎皮楠, 在最近调查时从群落中消失了, 随后出现了峨眉木姜子; 在样地3中, 调查初期的几年存在的绿叶石栎、峨眉木姜子、漆树(Toxicodendron succedaneum)、小叶青冈、野桐(Mallotus tenuifolius), 在最近调查时从群落中消失了, 随后出现了川鄂山茱萸(Cornus chinensis)、冬青(Llex sp.)、大叶冬青(Llex macrocarpa)、峨眉木姜子、木瓜红(Rehdodendron macrocarpa)、野樱(Maddenia hypoleuca)、川钓樟(Lindera pulchemima var. hemsleyana)、省沽油(Staphylea bumalda)、野桐。总体而言, 有11个种群从群落中消失, 而有10个种群出现, 基本保持了群落树种多样性的动态发展。
重要值是衡量树种在群落中的地位和作用大小的重要指标(王伯荪, 1987)。主要种群的重要值及其变化列于表 2。优势种群均为扁刺栲、华木荷(和绿叶石栎), 除样地3已经由调查初期的以扁刺栲、华木荷为优势种演变为扁刺栲、绿叶石栎、华木荷为优势的多优种群落, 而其它两个样地优势种地位基本不变, 但重要值发生了显著变化, 表明群落正从单优种向双优或多优势种群转变。其它种重要值均较小, 仍在群落中居于伴生地位。因此本文重点分析表 2中的主要种群的变化。
|
|
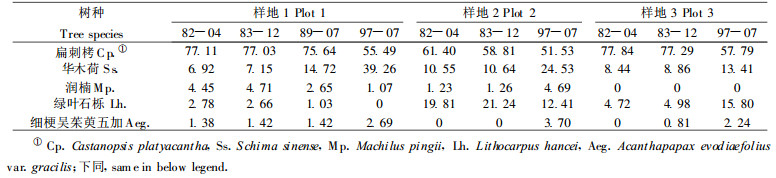
3样方中的优势种群密度变化如图 1所示。各主要种群在不同样方中的变化趋势是有差异的。第1优势种扁刺栲密度最大, 在最近15 a的自然恢复过程中, 密度表现出两种变化特点:初查密度较高, 则有下降趋势在1989年调查时达最高)(样地1);初查密度较低, 则先略有下降而后但都上升(样地2和3)。尽管最初密度差异较大, 但最终密度都有趋向接近由该区环境资源决定的某一密度值的趋势; 第2优势种群华木荷在自然恢复过程中, 均表现为增大趋势。绿叶石栎和润楠均表现出两种变化特点:种群密度下降或上升, 只是在样地2中的表现正相反。这些主要种群密度的变化反映了它们对群落环境的适应能力的差异和相互作用过程中的竞争能力强弱。扁刺栲种、华木荷、润楠、石栎都被认为是常绿阔叶林的顶极建群种(中国植被编写委员会, 1980), 可居于各群落优势地位。因而群落中的各种群密度的消长变化可能决定于它们的初始密度和环境的变化。

|
图 1 四川瓦屋山自然恢复的次生常绿阔叶林三个固定样地内主要乔木种群密度变化 Fig. 1 The Density ch anges of main tree population in secondary evergreen broad-leaved forest in Mt Wawushan, Sichuan
|
3个固定样地上各种群胸高直径DBH分布及其变化如图 2, 3, 4所示。表明在同一年龄的森林内, 各种群DBH在不同的样地中的分布特点差异较大, 类型多样。只有明显优势的种群扁刺栲的DBH呈正态分布或近似正态分布, 并随演变时间而整体的增长变化。其它种群DBH分布特点差异较大, 变化特点不一。各样地中, 在相互作用下各种群平均DBH值平均增长速度表现出较大差异(表 3)。平均DBH生长最快的是细梗吴茱萸叶五加(Acanthopanax evodiafolius var. gracilis), 依次为青冈(Cyclobalanopsis myrsinaefolia)、石栎、华木荷, 扁刺栲和润楠生长最慢(表 4)。

|
图 2 样地1上优势种种群胸径级结构及其变化 Fig. 2 Size structures and changes of main tree population in plot 1 |

|
图 3 样地2上优势种种群径级结构及其变化 Fig. 3 Size structures and changes of main tree population in plot 2 |

|
图 4 样地3上优势种种群径级结构及其变化 Fig. 4 Size structures and changes of main tree population in plot 3 |
|
|

|
|

从样地上主要种群的垂直高度分布格局来看(图 5), 最初期调查时(1982-04), 乔木层的下层由所有的润楠、小叶青冈个体以及小部分扁刺栲个体构成, 高度在4.5 m以下, 而乔木层的中上层则由大部分扁刺栲个体、所有的华木荷和绿叶石栎个体构成, 乔木层的立体空间分化不甚明显(3.5~6.7 m); 第2次调查时(1983-12), 除有少量润楠个体进入乔木层的中间亚层外, 各种群个体在乔木层的垂直高度分布格局没有变化, 而是整体性的增高, 乔木层的立体空间分化已较明显(3.8~7.5 m); 在最近调查时(1997-07), 华木荷种群的个体在乔木层的上中下亚层均有分布, 润楠个体则仍然主要在乔木层的中下亚层存在, 乔木层的立体空间分化十分明显(3.4~15 m)。调查时也发现, 在自然恢复过程中种群垂直高度分布格局的变化, 除受个体高生长的程序稳态控制外, 也受到种群内和种群间各个体由相互竞争而导致的稀疏作用影响, 一方面有存在的个体死亡, 也有灌木层的苗木生长进入乔木层的补充。

|
图 5 样地1主要种群个体高度垂直剖面及其变化 Fig. 5 The change and vertical distribution of main tree populations in plot 1
|
从各种群平均高生长变化来看, 均稳定增长, 但各种群间也差异明显(图 6)。年平均高生长率最高的是细梗吴茱萸叶五加, 依次为华木荷、小叶青冈、扁刺栲、润楠, 石栎高生长最慢(表 5)。

|
图 6 3个固定样地上主要种群平均高度变化 Fig. 6 Growth changes of the average height of main tree populations in three plots during recent 15 years

|
|
|

3固定样地上主要种群材积生产力高低不一, 样地1中, 华木荷、扁刺栲、润楠种群材积生产力分别为98.93 m3·hm-2、107.95 m3·hm-2和0.46 m3·hm-2, 样地2中, 华木荷、扁刺栲种群材积生产力分别为67.10 m3·hm-2和132.14 m3·hm-2; 样地3中, 华木荷、扁刺栲、石栎种群材积生产力分别为24.41m3·hm2, 143.833 m3·hm-2, 28.19 m3·hm-2。而1997年3固定样地材积量均在218~235 m3·hm-2之间。主要种群材积生产力过去15年的变化如图 7所示, 固-1样地优势种扁刺栲种群材积生产力并不是一直增长, 而是在后期下降了; 次优势种华木荷增长速度更快。润楠种群不仅没增长, 反而以枯立木形式几乎从群落中消失了。另两个样地主要种群材积生产力总体呈相同的增长规律。

|
图 7 三个固定样地中主要种群材积生产力及其变化 Fig. 7 The changes on volume productivity of main tree population in three plots
|
从所研究群落主要乔木种群胸径和高生长动态来看, 生长速度差异明显。平均高生长速度在24~48 cm·a-1, 由高至低的排序为小叶青冈、细梗吴茱萸叶五加、石栎、华木荷, 扁刺栲、润楠, 平均DBH生长率在0.34~1.10 cm.a-1, 由高至低的排序为细梗吴茱萸叶五加, 华木荷、小叶青冈、扁刺栲、润楠, 石栎(表 4)。综合分析, 可将这些树种按生长速度划分为适应性强的速生树种如细梗吴茱萸叶五加, 华木荷(平均DBH生长速度在0.70 cm·a-1以上), 中等速度的小叶青冈、扁刺栲(平均DBH生长速度在0.40 cm·a-1以上), 慢生种如润楠, 石栎(平均DBH生长速度在0.40 cm·a-1以下)。这是各种群在相同环境下和相互作用(竞争或排斥)下的生长差异结果, 能客观反映各种群生长对具体环境的适应能力及其相互作用, 与从解析木分析得出的结果有所不同(张水松等, 1997)。上述分析结果也为人工恢复和重建常绿阔叶林提供了较准确的依据, 如将细梗吴茱萸叶五加与华木荷混交可望获得较高的生产力。
3.2 乔木各种群密度与各种群生长及其材积生产力的关系3个固定样地上的乔木层各主要种群密度、生长速度和材积生产力差异较大。密度与种群平均高、平均DBH和生长和材积生产力的相关关系如表 5所示。除固-3的扁刺栲种群平均树高与其密度、固-1的扁刺栲种群材积生产力与其密度相关性不明显外, 其它均存在明显相关性。群落中的第1优势种扁刺栲初始密度大, 其种群密度变化与平均树高生长呈负相关, 与平均DBH生长变化呈正或负相关, 与材积生产力变化呈正相关; 对密度较小的第一伴生种群华木荷而言, 其种群密度变化与平均树高、平均DBH生长和材积生产力变化均呈显著正相关; 润楠和石栎种群除在固-1样地中平均树高、平均DBH生长和材积生产力与密度变化呈显著负相关外, 在其它两个样地中它们的平均树高、平均DBH生长和材积生产力增长与密度变化均呈显著正相关。表明在群落相同环境条件下和各种群相互作用下, 密度是各种群生长和材积生产力高低的关键因素。在群落恢复和重建过程中为了提高种群和群落生长量与材积生产, 应予以充分考虑群落和各种群的合理密度控制, 尤其是优势种群的密度。
3.3 乔木物种组成丰富度与材积生产力的关系1997年3固定样地材积量分别为218~235 m3·hm-2, 均高于相同林龄的江西次生常绿阔叶林材积生产力(张水松等, 1997), 而接近于福建省低山区人工促进天然更新和抚育管理的36年生栲树(Casatanopsis fargesii)纯林的材积生产力下限214~433 m3·hm-2(童书振等, 1997), 也与江西70~100 a的部分原生的木荷常绿阔叶林材积生产力(197~310 m3·hm-2和栲树常绿阔叶林材积生产力(220~294 m3·hm-2)下限相当, 而低于该区甜楮木的材积生产力(321~379 m3·hm-2)(詹有生, 1997)。表明自然恢复的常绿阔叶林不仅保持有较高的树种多样性(表 2), 也具有较高的材积生产力, 并不逊色于人工培育的常绿阔叶纯林; 在森林重建过程中, 追求较高的材积生产力与生物多样性的恢复和保护是可以兼顾的。这对于现阶段人工纯林和低效低质林改造, 对现有防护林建设中树种配置都有指导意义。
陈伟烈, 贺金生.中国亚热带地区的退化生态系统: 类型、分布、结构特征及恢复途径.见: 陈灵芝, 陈伟烈主编.中国退化生态系统, 北京: 中国科学技术出版社, 1995, 61~93
|
方精云, 刘国华, 徐松龄. 1996. 我国森林植被的生物量和净生产量. 生态学报, 16(5): 497-508. |
刘照光.岷江上游生态环境现状与恢复植被的途径.见: 姜恕, 陈昌笃主编.植被生态学研究.北京: 科学出版社, 1994, 399~408
|
刘照光, 张雨成, 吴宁.长江上游森林生态系统与可持续发展.见: 牛德水主编.农业生物学研究与农业持续发展.北京: 科学出版社, 1997, 227~232
|
刘金林, 等. 1983. 浙江午潮次生林演替过程分析. 植物生态学与地植物学丛刊, 7(1): 8-19. |
彭少麟, 方炜. 1995. 热带人工林生态系统重建过程物种多样性的发展. 生态学报, 15(增刊): 18-30. |
彭少麟, 方炜. 1996. 广州白云山次生常绿阔叶林结构动态. 应用与环境生物学报, 2(1): 22-29. |
王伯荪. 1987. 植物群落学. 广州: 高等教育出版社.
|
杨玉坡主编.长江上游(川江)防护林研究.北京: 科学出版社, 1993
|
中国植被编辑委员会. 1987. 中国植被. 北京: 科学出版社.
|
余作岳, 彭少麟. 1995. 热带亚热带退化生态系统的植被恢复及其效应. 生态学报, 15(增刊): 1-17. |
赵崇儒. 1991. 洪雅林场志. 成都: 四川人民出版社.
|
张水松, 林光, 等. 1997. 次生常绿阔叶林抚育改造技术研究. 林业科学研究, 10(5): 506-513. DOI:10.3321/j.issn:1001-1498.1997.05.010 |
张水松, 詹有生, 陈长发, 等. 1997. 常绿阔叶林树种组成株数比例和材积比例变化规律的研究. 林业科学, 33(4): 289-297. DOI:10.3321/j.issn:1001-7488.1997.04.001 |
詹有生. 1997. 江西天然次生米楮林组成及生产力研究. 林业科学研究, 10(4): 402-407. DOI:10.3321/j.issn:1001-1498.1997.04.011 |
Browns, Lugo A E. 1984. Biomass of tropical forest:An estimate based on forest volumes. Science, 223: 1290-1293. DOI:10.1126/science.223.4642.1290 |
Myint A K, Hofer T. 1998. Forestry and Key Asian Watersheds. ICIM OD: 44-49. |