 2000, Vol. 36
2000, Vol. 36文章信息
- 李镇宇, 王燕, 陈华盛, 许志春, 路永波.
- Li Zhenyu, Wang Yan, Chen Huasheng, Xu Zhichu, Lu Yongbo.
- 油松对赤松毛虫的诱导化学防御及滞后诱导抗性
- INDUCED CHEMICAL DEFENSES AND DELAYED INDUCED RESISTANCE OF PINUS TABULAEFORMIS CARR. TO DENDROLIMUS SPECTABILIS BUTLER
- 林业科学, 2000, 36(1): 66-70.
- Scientia Silvae Sinicae, 2000, 36(1): 66-70.
-
文章历史
- 收稿日期:1998-12-29
-
作者相关文章


昆虫取食植物, 植物能够调节其体内化学物质的数量或组成, 以对昆虫的取食产生撷抗(亦即诱导化学防御), 降低植物的营养水平, 产生高浓度的有毒物质如单宁、酚酸、生物碱、萜类化合物等(管致和, 1996;Karban, 1995;Haukioja, 1990)。根据不同的作用世代, 诱导化学防御可分为迅速的诱导抗性(RIR)和滞后的诱导抗性(DIR)。前者是指对当前世代植食性昆虫的影响, 后者是指对后继的一到几个世代植食性昆虫的影响(娄永根等, 1997;康乐, 1995)。研究滞后诱导抗性的问题, 在我国尚未见报道农作物大多为一年生产生诱导抗性后不久就趋于生命的结束而滞后诱导抗性只有在木本植物中才有可能, 因此在林业上研究滞后诱导抗性有着特殊的意义和作用。国外在这一方面已有一些研究, 如E.Haukioja等(1987)报道落叶松对灰线小卷蛾(Zeiraphera griseana)的滞后诱导抗性可持续3~4年; S.R.Leather等(1987)报道扭叶松对小眼夜蛾(Panolis flammea)的滞后诱导抗性在一年以上。
赤松毛虫(Dendrolimus spectabilis Butler)是危害油松的一种主要害虫, 研究发现该虫的发生具有周期性的规律, 且在爆发后的后继世代危害较轻。因此本文主要探讨受害油松新长出的一年生针叶化学成分的变化及其对赤松毛虫幼虫发育的影响, 以及油松滞后诱导抗性持续时间的长短, 从而对赤松毛虫的综合治理和持续控制提供理论依据。
1 材料与方法 1.1 赤松毛虫幼虫的室内饲养在室内自然温度、光照、保湿条件下, 用当年已受赤松毛虫危害(1995年4月已有50%针叶受损)和未受危害(针叶损失5%以下)的油松新长出的一年生针叶饲养赤松毛虫初孵幼虫, 每一处理饲养500头, 每25头幼虫放入一个500mL的广口玻璃瓶中饲养, 直至4龄幼虫开始越冬。在不同发育时期用TG-328B光电分析天平称体重, 并定期统计成活率。
1.2 针叶化学成份的测定各种针叶化学成分的分析方法参见《油松对赤松毛虫的诱导化学防御》 (李镇宇等, 1998)。
2 结果与分析 2.1 油松针叶化学成分含量分析 2.1.1 针叶内粗脂肪、糖、单宁、生物碱含量分析受害油松新长出的一年生针叶内除单宁含量降低以外, 粗脂肪、糖、生物碱、糖/氨基酸之比均上升, 尤以生物碱增加最显著, 达138.5%, 其结果与油松二年生针叶成分相似, 见表 1。
|
|
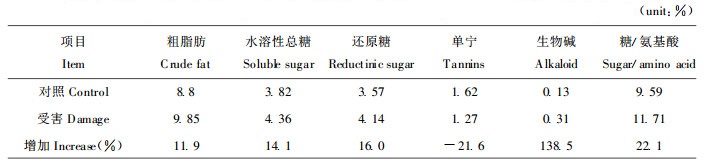
植物与昆虫之间是不断相互选择、协同进化的(钦俊德, 1987)。昆虫不同发育阶段对营养的需求不同, 植物则采取不同的防御策略(周章义等, 1986)。赤松毛虫老龄幼虫主要取食油松二年生针叶, 取食量大, 油松受害后二年生针叶内单宁含量增加, 抗性增强(李镇宇等, 1998)。单宁是干扰昆虫对食物利用的物质, 首先是干扰肠道消化, 使消化酶失去活性, 不能分解利用鞣化的蛋白质; 与淀粉形成络合物, 不能消化淀粉。此外, 植物在合成和积累缩合单宁时, 在代谢上要花很大代价(管致和等, 1996)。由于植物对植物化学防御系统投入的能量和代谢均有一定的限度, 为此植物在进化过程中趋于发展耗能较低和效用较大的防御方法。赤松毛虫低龄幼虫主要以新长出的一年生针叶为食, 取食量少。因此, 受害松树新长出的一年生针叶改变防御策略, 单宁含量降低, 生物碱含量急剧上升。主要作用于昆虫的味觉而引起昆虫忌避或抑制取食, 同时也具抗生作用引起昆虫中毒, 如番茄苷(tomatine)和垂茄碱(demissine)对马铃薯叶甲(Leptiontarsa decemlineata)有较强的拒食作用, 对菜粉蝶有致畸毒性(许美娟等, 1994)。
已经知道芳香油及某些有机酸对昆虫有毒。一年生针叶粗脂肪含量升高, 对昆虫有毒的物质也会随之增多, 这些物质在幼虫体内积累到一定程度就会导致幼虫生长缓慢, 甚至死亡(马尾松毛虫抗性研究组, 1990)
一般认为食物中可溶性糖含量高有利于幼虫的存活及生长发育(周章义等, 1986, Todd, 1971), 而糖/蛋白质比例的失调则会增加昆虫的代谢压力(Harvey, 1974)。一年生针叶内水溶性糖、还原性糖、糖/氨基酸之比均增加, 反而增加了昆虫的代谢压力, 从而影响松毛虫的正常生长和发育。如蚜虫对糖的需求量不大, 排蜜即是在长期进化过程中形成的对体内营养物质平衡的一种调节(钦俊德, 1987)。
2.1.2 针叶脂肪酸含量分析脂肪酸是昆虫体内的重要能源物质。由表 2可见, 受害小的油松二年生针叶及受害大的油松针叶中脂肪酸各组分有大幅度的变化, 针叶中多种脂肪酸如肉豆蔻酸、油酸及脂肪酸总量有较大幅度下降。
|
|
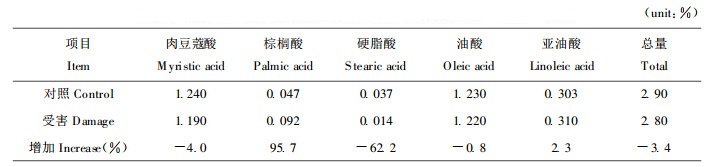
脂肪酸是昆虫体内的重要能源物质。表 2的结果表明, 除亚油酸略有增加, 棕榈酸增加较多以外, 其余几种脂肪酸均减少, 尤以硬脂酸减少最多, 脂肪酸的总量减少3.4%。其中棕榈酸含量增加, 其原因是由软脂酸和硬脂酸去饱和转化的结果(沈同等, 1981)。有些脂类物质虽然并非昆虫所必需, 但它们可能使昆虫对其它营养物质的吸收和利用更为容易(钦俊德, 1976; 1987)。当然不能只分析某一种脂肪酸含量的变化, 因为脂肪酸混合物的效果均优于任何一种单独的脂肪酸(钦俊德, 1987)。
2.1.3 游离氨基酸含量分析氨基酸是昆虫发育的主要营养物质。由图 1可见, 受害油松针叶中大部分游离氨基酸均有大幅度的减少, 氨基酸总量减少6.5%, 尤以昆虫所必需的亮氨酸减少最多, 达66.9%。

|
图 1 油松一年生针叶游离氨基酸含量变化 Fig. 1 Change of free amino acid contents in one-year needles of pine P.tabulaeformis □对照CK; 受害Damaged |
食物中蛋白质的营养价值主要决定于所含氨基酸的种类和比例(Harvey, 1974)。昆虫的组织在整个发育过程中对氨基酸以及其它含氮化合物如核酸等的需要是处于变动状态的, 所以在转氨作用中处于中心位置的那些不稳定的氨基酸, 特别是谷氨酸和天门冬氨酸, 严格说并非昆虫所必需, 但在很多昆虫中却作为补充营养成分而处于重要地位, 这也是要求食物中各种氨基酸按适当比例存在的理由(钦俊德, 1987)。因此, 可以认为氨基酸含量的大幅度变化和总量的减少是油松诱导抗性的一个方面。
2.1.4 油松针叶内酚酸含量分析酚酸是一类重要的次生抗虫化合物, 存在于大麦和其它植物中的香草酸、没食子酸、丁香酸、香豆酸、咖啡酸等的含量影响麦二叉蚜(Schizsphis graminum)的生长和繁殖儿茶酚在洋葱中有抗病作用这种成分对麦二叉蚜有毒并能抑制小地老虎(Agrotis ypsilon Rott.)的生长(Meyer, 1974)。酚酸总量有所下降, 其原因可能是所测酚酸对赤松毛虫作用不大。如金丝梅(Hypericum)中的金丝梅蒽酮(hypericin)也是对动物有毒性的酚酮, 但对金叶甲(Chrysolina brunsvicensis)则是助食剂(Rees, 1969)。

|
图 2 油松一年生松针酚酸含量变化 Fig. 2 Change of phnolics contents in one-year needles of pinc P.tabulaeformis □对照CK; 受害Damaged. |

|
图 3 油松一年生针叶饲养幼虫死亡率变化 Fig. 3 Change of mortality of larvaes fed on pine needles ……对照CK; ———受害Damaged |

|
图 4 油松一年生针叶饲养幼虫体重变化 Fig. 4 Change of weight of larvaes fed on pine needles —□—对照CK; —△—受害Damaged |
室内养虫结果表明, 受害油松长出的新针叶饲养的赤松毛虫均比对照体重减轻, 死亡率增加。9月15日, 受害油松长出的新叶饲养赤松毛虫死亡率比对照高出12.2%, 至10月24日高达19.2%。说明当年4月油松受到赤松毛虫危害, 会产生诱导化学防御以使针叶化学成分发生变化, 这种变化不仅对当前世代的赤松毛虫的生长发育产生不利影响(李镇宇等, 1998), 而且存在着时滞效应, 对后继世代的赤松毛虫产生影响。
3 结论与讨论研究植物的诱导化学防御和滞后诱导抗性, 不仅能在理论上加深对植食性昆虫种群动态机制、昆虫与植物相互关系、昆虫种间和种内相互作用以及昆虫群落构建机制等的认识, 而且还能在实践上从一个侧面补充和完善害虫综合治理理论(娄永根等, 1997)。另外, 对减少化学农药或不使用化学农药防治害虫的研究都有重要意义。因此, 近年来植物诱导化学防御和滞后诱导抗性的研究受到高度重视。本文通过对当年受赤松毛虫危害和未受危害的油松新长出的一年生针叶化学成份以及用其饲养的赤松毛虫的发育情况的分析, 发现受害油松针叶化学成份仍有较大幅度的变化, 氨基酸总量及多数游离氨基酸含量下降; 水溶性总糖、粗脂肪、生物碱含量及总糖/氨基酸比例上升; 脂肪酸、单宁、酚类化合物均有较大幅度的减少; 用其饲养的赤松毛虫幼虫体重减轻、死亡率增加。证明油松对赤松毛虫存在着滞后诱导抗性, 持续期至少在半年以上。
植物体内的生化反应是有机联系的整体, 代谢途径繁多, 代谢产物之间的关系复杂。植物本身就处于一种动态变化过程中即基础代谢和次生代谢的交替进行而植物的抗性功能正是存在于这种动态的变化过程中(王海波, 1993)。同时, 植物的营养物质变化, 甚至水分的变化均可以起到防御昆虫取食的作用, 并且, 化学物质和营养物质往往是协同作用, 共同对昆虫起着防御的作用(康乐, 1995)。另外, 植物的诱导防御是一个有机的整体, 包括许多方面与环节, 而不是单纯依赖某一些物质的增加或减少。因此, 植物体内的各种物质在诱导化学防御中的地位及其协调作用还有待于进一步的探讨。同时, 由于各种原因, 本文只对油松受害的二年生针叶和当年新生针叶进行了测定和分析, 而没能进行多年跟踪测定对于受害油松的滞后诱导抗性到底能持续多久还有待于深入研究.
管致和主编.植物医学导论.北京: 中国农业大学出版社, 1996
|
康乐. 1995. 环境胁迫下的昆虫-植物相互关系. 生态学杂志, 14(5): 51-57. |
李镇宇, 陈华盛, 袁小环, 等. 1998. 油松对赤松毛虫的诱导化学防御. 林业科学, 34(2): 43-49. DOI:10.3321/j.issn:1001-7488.1998.02.007 |
娄永根, 程家安. 1997. 植物的诱导抗虫性. 昆虫学报, 40(3): 321-327. |
马尾松毛虫抗性研究组. 1990. 抗性植株的抗性机制研究. 林业科学, 26(2): 133-140. |
钦俊德. 1976. 昆虫学研究进展. 北京: 科学出版社.
|
钦俊德. 1987. 昆虫与植物的关系. 北京: 科学出版社.
|
钦俊德. 1995. 昆虫与植物关系的研究进展和前景. 动物学报, 4(1): 12-19. DOI:10.3321/j.issn:0001-7302.1995.01.008 |
沈同, 等. 1981. 生物化学(下). 北京: 高等教育出版.
|
王海波. 1993. 昆虫胁迫下的植物应急反应模式. 生态学杂志, 12(6): 46-48. DOI:10.3321/j.issn:1000-4890.1993.06.012 |
许美娟, 管致和. 1994. 番茄植株中生物碱类对菜青虫(Pieris rapae L.)活性组分的分离和提纯. 北京农业大学学报, 20(2): 153-156. |
康乐. 1995. 植物对昆虫的化学防御. 植物学通报, (4): 22-27. |
周章义, 等. 1986. 林业营养与虫害油松施N肥的抗虫效应. 昆虫学报, 29(3): 283-290. |
Harvey G T. 1974. Nutritionalstudies of eastern spruce budworm Ⅰ.soluble sugars. Can.Ento mol., (93): 594-602. |
Haukioja E, Neuvonen S.Insect population on oviposition growth and induction of plant resistance : the testing of hypotheses in : Barbosa P.Schultz T.C.eds, Insect outbreaks.San Diego : Academic, 1987, 411~430
|
Haukioja E. 1990. Theories of plant chemical defense in trees. Ann.Rev.Entomol., 36: 25-42. |
Karban R, Nhho C. 1995. Induced resistance and suscetibility to herbivory :Plant memory and altered plant development. Ecology, 76(4): 1220-1225. DOI:10.2307/1940928 |
Karban R. 1993. Costs and benefits of induced resistance and plant density for a native shrub, Gossypium thurberi. Ecology, 74(1): 9-19. DOI:10.2307/1939496 |
Leather S R, Watt A F, et al. 1987. Insect-induced chemical changes in young lodgpole pine (Pinus contorta); the effect of previous defoliation onoviposition, growth and survival ofthe pine beauty moth, Panolis flammea. Ecol.Entomol, (12): 275-281. |
Meyer H J & Norris D M.Lignin intermedites and simple phenolics as feeding stimulants for Scolytus multistriatus.J.Insect physiol., 1974, 20:2015~2021
|
Rees C J C. 1969. Chemoreceptor specificity associated with choice of feeding site by the beetle Chrysolina bruusvicens onits footplant. Hypericumhirsutum.Ent.expt.appl, 12: 565-583. DOI:10.1111/j.1570-7458.1969.tb02554.x |
Todd G W, Getahun A & Cress D C.Resistancein barley to the greenbug, Schizaphis graminum.I.Toxicity of phenolic and flavonoid co mpoudsand related substances.Ann.Ent.Soc.Amer., 1971, 64:718~722
|