 2009, Vol. 45
2009, Vol. 45文章信息
- 臧润国, 刘贵峰, 巴哈尔古丽·阿尤甫, 郭仲军, 白志强, 张炜银, 丁易.
- Zang Runguo, Liu Guifeng, Ayoufu Bahaerguli, Guo Zhongjun, Bai Zhiqiang, Zhang Weiyin, Ding Yi
- 天山云杉球果大小性状的地理变异规律
- Geographic Variation of Cone Size of Picea schrenkiana var. tianschanica in the Tianshan Mountains of Xinjiang
- 林业科学, 2009, 45(2): 27-32.
- Scientia Silvae Sinicae, 2009, 45(2): 27-32.
-
文章历史
- 收稿日期:2008-03-10
-
作者相关文章


2. 内蒙古民族大学农学院 通辽 028000;
3. 新疆林业科学院森林生态研究所 乌鲁木齐 830000
2. Agricultural College, Inner Mongolia University for the Nationalities Tongliao 028000;
3. Institute of Forest Ecology, Xinjiang Academy of Forestry Urumqi 830000
天山是亚洲中部最大的山脉,西起乌兹别克斯坦的克孜尔库姆沙漠以东,经哈萨克斯坦和吉尔吉斯斯坦进入我国新疆境内,全长约2 500 km,宽约250~300 km,在我国境内约1 700 km。相对优越的自然条件使得天山森林成为新疆森林生态系统的重要组成部分。天山云杉(Picea schrenkiana var. tianschanica)为天山森林的优势种和建群种,是新疆山地森林中分布最广、蓄积量最大、用途最广的树种。天山云杉林占新疆天然林有林地总面积的44.9%和蓄积量的61.3%,在水源涵养、水土保持、绿洲农业等方面发挥着至关重要的生态作用(新疆森林编辑委员会,1989)。此外,天山是天山云杉在我国的唯一分布区,天山云杉为该地区特有珍贵针叶树种,具有重要的科研价值。
表型多样性是遗传多样性研究的重要内容,是遗传多样性与环境多样性的综合体现(阎爱民等,1999)。表型多样性主要研究种群在其分布区内各种环境下的表型变异。植物形态特征的变异有其固有的遗传基础,受遗传与环境的综合作用,形态特征变化常常可作为遗传变异的表征之一。表型多样性研究主要通过科学有效地采样,合理的数学统计方法,采用总体形态性状、繁殖器官性状、生理生化性状等较稳定的性状,来揭示群体的遗传规律、变异大小,以客观评价其遗传多样性(顾万春,2004)。植物表型变异的研究主要来源于分类学(Ishimitsu et al., 1983; Marcysiak et al., 2007; Medrano et al., 2006; Newell, 1989; Otieno et al., 2006),表型性状是环境与遗传因素共同作用的结果,因此对表型性状进行研究不仅能初步了解种群遗传变异的大小,而且有助于了解生物对不同环境的适应性。表型多样性是生态学及遗传学研究的一个重要领域(Cordell et al., 1998; Cron et al., 2007; Hornung-Leoni et al., 2006; Mizianty et al., 2007; Schoettele et al., 2000)。对表型变异的研究主要集中于对变异模式的探索、变异来源以及变异与生态因子和地理因素的关系等(Hausmann et al., 2001; Joel et al., 1994; Sanou et al., 2006)。利用表型性状研究种群的遗传多样性具有简便、快速和节省费用等优点,至今仍然是重要而且有效的方法(Lagercrantz et al., 1990)。许多针叶树种研究者通过分析针叶、球果和种子的表型性状来探讨性状变异、分化及其与环境因子的关系(Dangasuk et al., 2004; Donahue et al., 1996; 葛颂,1988;Klimko et al., 2007; Linhart, 1978)。球果和种子是植物繁殖系统的重要组分, 它们常常在不同的选择压力下表现出较大的适应性。
天山云杉是我国干旱区山地极其珍贵的树种,具有一系列对环境的适应与响应特征,开展其生态地理学的研究具有重要的理论与实践意义。本文选择位于中国境内天山山脉的5个纬度相近但经度不同的天山云杉林分布的代表性地点(昭苏、巩留、乌苏、乌鲁木齐和哈密),通过测定各地点不同海拔梯度上天山云杉种群的球果长和宽,初步分析种群球果性状的生态地理变异规律,以期为天山云杉种群保护、优良种源选育及其合理利用提供依据。
1 研究区概况选择我国新疆境内天山山脉从西到东处于不同经度位置5个地点的天山云杉林进行垂直样带调查,5个地点从西至东分别为昭苏北山、西天山国家级自然保护区、乌苏巴音沟、乌鲁木齐水西沟天山森林生态系统定位研究站和哈密白石头乡(表 1)。
|
|
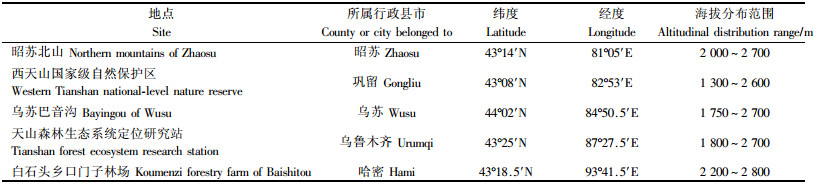
在5个调查地点,从天山云杉林自然分布的下限到上限,每隔50 m划分为一个海拔段。在每个海拔段内随机选取3株成熟(结球果)的天山云杉个体,在每株树木树冠中部随机采集球果20个,每个海拔段采集球果共60枚。采集的球果立即装入塑料密封袋中防止脱水。完成球果采集后,记录每个海拔段的坡度、坡向、坡位等生境因子。球果采集时间是2006年9月上旬—10月上旬。采集的球果带回室内,从每个海拔段内采集的球果中随机选取30个,用游标卡尺测定每个球果的长、宽(最宽处),测量精度为0.001 cm,计算球果长宽比。
2.2 分析方法首先将各调查地点的海拔分别尽可能地等间隔划分为低、中、高3个海拔范围(由于山体高度和经度位置的不同,对各地点海拔的高、中、低范围划分标准不一致),分析天山云杉球果性状随海拔变化的基本特征。另一种是采用不同的函数对球果性状(球果长、宽和长宽比)沿每隔50 m的海拔梯度变化规律进行回归分析。对各性状的差异性采用方差分析,球果性状与经度、纬度、海拔、坡度、坡向、坡位的关系采用相关分析。所有分析在SPSS16.0统计分析程序中完成,显著度设为P < 0.005。
3 结果与分析 3.1 不同海拔范围内天山云杉球果性状的比较处于不同经度位置5个地点的天山云杉种群球果性状随海拔的不同呈现不同的变化规律(图 1)。球果长在昭苏、乌苏、乌鲁木齐3个地点的变化趋势均显著表现为低海拔>中海拔>高海拔。而巩留低海拔和中海拔的天山云杉球果长无显著差异,但均显著高于高海拔;哈密低海拔天山云杉球果长显著高于中海拔和高海拔,但中高海拔之间的无显著差异。球果宽的变化趋势较为复杂,在乌苏(图 1c)和乌鲁木齐(图 1d),球果宽的变化趋势与球果长的变化趋势一致,均显著表现为低海拔>中海拔>高海拔;在昭苏是低、高海拔较大,无显著差异,与中海拔有显著差异(图 1a);巩留在低海拔最小,中、高海拔较大且二者之间无显著差异,与低海拔有显著差异(图 1b);在哈密球果宽是高海拔>中海拔≥低海拔,且中、低海拔之间无显著差异,与高海拔有显著差异(图 1e)。5个地点球果长宽比的变化趋势一致,均为低海拔>中海拔>高海拔,且都有显著差异。

|
图 1 天山5个地点低、中、高3个不同海拔级内天山云杉球果大小(平均值±标准误)性状比较 Figure 1 Comparison of the cone size traits (Mean±SE)of P. schrenkiana var. tianschanica in three altitudinal range (low, middle and high) in five sites of different longitude in Tianshan mountains of Xinjiang |
昭苏、巩留、乌苏和乌鲁木齐的天山云杉球果长随海拔的升高而下降(图 2);哈密天山云杉球果长随海拔梯度的变化不显著,但总体呈现下降的趋势(图 2e)。5个研究地点天山云杉球果长总体表现为随海拔增加而下降的趋势。球果宽随海拔的变化趋势较为复杂(图 3)。昭苏和巩留的天山云杉球果宽随海拔梯度变化不显著;在中部的乌苏和乌鲁木齐球果宽随海拔的升高而下降;而在东部的哈密球果宽随着海拔的上升而增加。球果长宽比随海拔的变化趋势较为一致,在昭苏、巩留、乌苏、乌鲁木齐和哈密5个地点的天山云杉球果长宽比均呈现随着海拔的上升而下降的趋势(图 4)。

|
图 2 天山云杉球果长(平均值±标准误)随海拔梯度的变化 Figure 2 The cone length (Mean±SE)variation of P. schrenkiana var. tianschanica along the altitudinal gradient |

|
图 3 天山云杉球果宽(平均值±标准误)随海拔梯度的变化 Figure 3 The cone width variation (Mean±SE)of P. schrenkiana var. tianschanica along the altitudinal gradient |

|
图 4 天山云杉球果长宽比(平均值±标准误)随海拔梯度的变化 Figure 4 The cone length/width ratio (Mean±SE) variation of P. schrenkiana var. tianschanica along the altitudinal gradient |
天山云杉球果形态性状在5个研究地点存在显著差异(表 2)。除乌鲁木齐和哈密之间无显著差异外,其余不同地点之间球果长均存在显著差异,西部巩留球果长最大,中部乌苏球果长最小。除西部的昭苏和巩留球果宽之间无显著差异外,其余不同地点之间均存在显著差异,其中乌鲁木齐球果宽最大,乌苏最小。不同地点之间天山云杉球果长宽比均存在显著差异,西部的巩留球果长宽比最大,中部的乌苏最小。天山云杉球果长、球果宽和球果长宽比3个性状的平均变异系数分别为17.01%,10.99%和15.50%,表明球果宽较球果长稳定性高。
|
|

天山云杉球果长与球果宽、球果长宽比呈极显著正相关(表 3)。球果长与海拔呈极显著负相关,与坡度呈显著负相关,与经度呈负相关但不显著。球果宽与经度呈极显著正相关,与海拔呈负相关趋势但没有达到显著水平。球果长宽比与海拔、经度、坡度都呈极显著负相关。
|
|

本文研究表明5个研究地点天山云杉球果长与海拔呈现显著的负相关,但球果宽则没有一致的变化规律,在不同研究地点表现出不同的相关性。国内对其他云杉球果性状研究表明球果长和球果宽与海拔也表现出不同的相关性。例如云杉(Picea asperata)和秦岭冷杉(Abies chensiensis)球果长、球果宽与海拔都呈负相关(罗建勋等,2003;孙玉玲等,2005a;2005b)。但江洪(1992)通过分析云杉种群性状分化与海拔梯度的关系发现,球果长与海拔梯度的关系为一种典型的负相关,而球果宽与海拔梯度则为一种明显的正相关。本研究中仅昭苏和乌鲁木齐2个地点的天山云杉球果宽与海拔显著相关。植物性状与海拔的关系不仅与海拔变化带来的地形变化相关联,而且还受到森林群落分布的局域环境的影响。新疆天山云杉林为浅根系树种,抗风能力较差,因而土壤结构和风力强度能够显著影响天山云杉生长状况。高海拔地区较短的生长周期、较差的土壤条件和较强的风力可能会影响高海拔范围天山云杉的营养生长,因而降低了天山云杉对生殖器官的能量分配。也有一些研究得出了与此不同的结果,如加纳利松(Pinus canariensis)球果和种子在较高海拔林分更大,这可能是对不利环境条件(寒冷和干旱)的适应(Gil et al., 2002)。徐化成(1992)在油松(Pinus tabulaeformis)的研究中也发现,干旱寒冷的区域球果更大。岷江柏(Cupressus chengiana)球果大小随着海拔的增加而增加(徐亮等,2004)。天山云杉主要分布在阴坡即北坡、西北坡和东北坡,因此坡向对球果生长没有显著影响。
天山云杉球果长和宽的变异可能与气温和降水相关。海拔梯度的变化反映了气候条件的变化,随着海拔的上升,气温相应地下降,但降水则随基带气候条件和地形的变化而不同。球果长与经度虽没有达到显著相关水平,但也呈负相关趋势,整体呈现西部>中东部的趋势;而球果宽则呈现中东部>西部的趋势;球果长宽比呈现西部>中东部的趋势。在天山山脉,西部伊犁地区温暖湿润,降雨量丰富;中部乌鲁木齐地区气候干燥,雨量适中;东部哈密地区干旱炎热,雨水较少。沿天山山脉,从西至东,降水量总体上呈现逐渐减少的趋势,但不同地点内不同海拔范围内的降水格局会发生不同的变化,天山云杉球果性状的变异会随区域大气候梯度和局域小气候及其他环境梯度而变化。球果宽性状较球果长性状的稳定性高,这与天然红松(Pinus koraiensis)种群的研究结果一致(张恒庆等,1999)。本文的调查研究表明天山云杉球果形态特征随海拔梯度变化的规律性比随经度梯度变化的规律性强。
葛颂. 1988. 同工酶与林木群体遗传变异研究. 南京林业大学学报, (1): 68-77. |
顾万春. 2004. 统计遗传学. 北京: 科学出版社.
|
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社.
|
罗建勋, 李晓清, 孙鹏, 等. 2003. 云杉天然群体表型变异. 东北林业大学学报, 31(1): 9-11. DOI:10.3969/j.issn.1000-5382.2003.01.003 |
孙玉玲, 李庆梅, 杨敬元, 等. 2005a. 秦岭冷杉球果与种子的形态变异. 生态学报, 25(1): 176-181. |
孙玉玲, 李庆梅, 谢宗强. 2005b. 濒危植物秦岭冷杉结实特性的研究. 植物生态学报, 29(2): 251-257. |
新疆森林编辑委员会. 1989.新疆森林.乌鲁木齐: 新疆人民出版社; 北京: 中国林业出版社.
|
徐化成. 1992. 油松地理变异和种源选择. 北京: 中国林业出版社.
|
徐亮, 包维楷, 何永华. 2004. 4个岷江柏种群的球果和种子形态特征及其地理空间差异. 应用与环境生物学报, 10(6): 707-711. DOI:10.3321/j.issn:1006-687X.2004.06.007 |
阎爱民, 陈文新. 1999. 苜蓿、草木樨、锦鸡儿根瘤菌的表型多样性分析. 生物多样性, 7(2): 1-8. |
张恒庆, 安利佳, 祖元刚. 1999. 天然红松种群形态特征地理变异的研究. 生态学报, 19(6): 932-938. DOI:10.3321/j.issn:1000-0933.1999.06.029 |
Cordell S, Goldstein G, Mueller-Dombois D, et al. 1998. Physiological and morphological variation in Metrosideros polymorpha, a dominant Hawaiian tree species, along an altitudinal gradient: the role of phenotypic plasticity. Oecologia, 113: 188-196. DOI:10.1007/s004420050367 |
Cron G V, Balkwill K, Knox E B. 2007. Multivariate analysis of morphological variation in Cineraria deltoidea (Asteraceae, Senecioneae). Botanical Journal of the Linnean Society, 154(4): 497-521. DOI:10.1111/j.1095-8339.2007.00664.x |
Dangasuk O G, Panetsos K P. 2004. Altitudinal and longitudinal variations in Pinus brutia (Ten.) of Crete Island, Greece: some needle, cone and seed traits under natural habitats. New Forests, 27(3): 269-284. DOI:10.1023/B:NEFO.0000022227.33131.f0 |
Donahue J K, Upton J L. 1996. Geographic variation in leaf, cone and seed morphology of Pinus greggii in navite forests. Forest Ecology and Management, 82: 145-157. DOI:10.1016/0378-1127(95)03677-6 |
Gil L, Climent J, Nanos N, et al. 2002. Cone morphology variation in Pinus canariensis Sm. Plant Systematics and Evolution, 235(1): 35-51. DOI:10.1007/s00606-002-0218-9 |
Hausmann S, Lotter A F. 2001. Morphological variation within the diatom taxon Cyclotella comensis and its importance for quantitative temperature reconstructions. Freshwater Biology, 46(10): 1323-1333. DOI:10.1046/j.1365-2427.2001.00752.x |
Hornung-Leoni C, Sosa V. 2006. Morphological variation in Puya (Bromeliaceae): an allometric study. Plant Systematics and Evolution, 256: 35-53. |
Ishimitsu M, Tateoka T. 1983. Morphological variation and taxonomy of the Poa macrocalyx complex (Poaceae) in Hokkaido. Journal of Plant Research, 96(3): 245-267. |
Joel G, Aplet G, Vitousek P M. 1994. Leaf morphology along environmental gradients in Hawaiian Metrosideros Polymorpha. Biotropica, 26(1): 17-22. DOI:10.2307/2389106 |
Klimko M, Boratynska K, Montserrat J M, et al. 2007. Morphological variation of Juniperus oxycedrus subsp. oxycedrus (Cupressaceae) in the Mediterranean region. Flora-Morphology, Distribution, Functional Ecology of Plants, 202(2): 133-147. DOI:10.1016/j.flora.2006.03.006 |
Lagercrantz U, Ryman N. 1990. Genetic structure of Norway spruce (Picea abies): concordance of morphological and allozymic variation. Evolution, 44(1): 38-53. |
Linhart Y B. 1978. Maintenance of variation in cone morphology in California closed-cone Pines: the roles of fire, squirrels and seed output. The Southwestern Naturalist, 23(1): 29-40. DOI:10.2307/3669977 |
Marcysiak K, Boratyński A. 2007. Contribution to the taxonomy of Pinus uncinata (Pinaceae) based on cone characters. Plant Systematics and Evolution, 264(1): 57-73. |
Medrano M, Castellanos M, Herrera C. 2006. Comparative floral and vegetative differentiation between two European Aquilegia taxa along a narrow contact zone. Plant Systematics and Evolution, 262(3): 209-224. |
Mizianty M, Frey L, Bieniek W, et al. 2007. Variability and structure of natural populations of Hordelymus europaeus (L.) Jess. ex Harz and Leymus arenarius (L.) Hochst. as revealed by morphology and DNA markers. Plant Systematics and Evolution, 269(1): 15-28. |
Newell S J. 1989. Variation in leaflet morphology among populations of Caribbean cycads (Zamia). American Journal of Botany, 76(10): 1518-1523. DOI:10.1002/ajb2.1989.76.issue-10 |
Otieno D, Balkwill K, Paton A. 2006. A multivariate analysis of morphological variation in the Hemizygia bracteosa complex (Lamiaceae, Ocimeae). Plant Systematics and Evolution, 261(1): 19-38. |
Sanou H, Picard N, Lovett P, et al. 2006. Phenotypic variation of agromorphological traits of the shea tree, Vitellaria paradoxa C.F. Gaertn., in Mali. Genetic Resources and Crop Evolution, 53(1): 145-161. DOI:10.1007/s10722-004-1809-9 |
Schoettele A W, Rochelle S G. 2000. Morphological variation of Pinus flexilis (Pinaceae), a bird-dispersed pine, across a range of elevations. American Journal of Botany, 87(12): 1797-1806. DOI:10.2307/2656832 |