 2009, Vol. 45
2009, Vol. 45文章信息
- 理永霞, 茶正早, 罗微, 林钊沐, 贝美容.
- Li Yongxia, Cha Zhengzao, Luo Wei, Lin Zhaomu, Bei Meirong
- 3种桉树幼苗叶片养分变化及其转移特性
- Dynamics and Transfer of Nutrients in the Seedling Leaves of Three Eucalyptus Varieties
- 林业科学, 2009, 45(1): 152-157.
- Scientia Silvae Sinicae, 2009, 45(1): 152-157.
-
文章历史
- 收稿日期:2007-11-15
-
作者相关文章



2. 南京林业大学 南京 210037
2. Nanjing Forestry University Nanjing 210037
我国桉树人工林面积已超过了100万hm2,成为我国南方面积最大的人工林树种之一,并以每年约10万hm2的速度发展。桉树种植一直以来存在广泛争议,有些学者认为它掠夺地力,并大幅降低地下水位;但亦有不同的证据(徐大平等,2006;杨曾奖等,2006),也有部分学者认为桉树与其他具有同等生产力的树种,如橡胶树(Hevea brasiliensis)和马占相思(Acacia mangium)比较,其养分利用效率是比较高的(余雪标等,2000)。然而,与高效利用特征直接相关的营养利用机理尚没有深入的研究。
营养利用机理的核心是体内的营养转移,即体内养分的再利用,特别是反映在衰老枝叶内的养分转移,它使枝叶的凋落不至于造成大量营养的损失,被认为是物种适应贫瘠土地的重要机制,它提高了体内养分的利用效率,降低了对环境中养分供应的依赖(Chapin,1980;孙书存等,2001;李志安等,2003;王希华等,2004;薛立等,2005)。营养转移是普遍存在的现象,早在1912年Ramann就注意到叶片衰老过程中有部分N、P、K可能运回树体,1927年Gombes证实了这一推论(王文卿等,1999;廖利平,1994)。本文主要从养分转移的角度,探讨巨尾桉(Eucalyptus grandis × E.urophylla)广林-9号、184-1,尾赤桉(Eucalyptus urophylla ×E.camaldulensis)201-2等3个桉树品种在营养利用上的特征,为这几个品种的合理经营、科学施肥提供参考。
1 材料和方法 1.1 试验材料试验在中国热带农业科学院橡胶研究所五队基地自然条件下进行,位于19°32′N,109°14′E,地处热带季风气候区;冬季雨量稀少,夏秋雨水充沛,年均降雨量为1 815 mm;年平均气温23.2 ℃。供试苗木是尾赤桉184-1、201-2、巨尾桉广林-9扦插苗木;试验塑料黑桶内进行,桶规格高和口径是35 cm×30 cm;供试土壤是花岗片麻岩发育的砖红壤,pH值5.65,有机质含量14.2 g·kg-1,全氮含量0.705 g·kg-1,有效磷含量17.26 mg·kg-1,速效钾含量30.8 mg·kg-1,试验所用肥料分别是过磷酸钙(P2O5%≥16%)、氯化钾(K2O%≥60%)、尿素(含氮量46%)、复合肥(总养分≥16%,有机质≥20%)、桉树专用肥(总养分≥15%)。
1.2 试验设计试验苗木于2006年3月定植。试验分3个品种,每个品种分施肥与不施肥2个处理,每个处理重复8次,随机排列。苗木栽植后适时浇灌,分别于2006年6、8、10月及2007年2月共施肥4次,其中6月施的是过磷酸钙和尿素每株分别为4 g和6 g;8月施氯化钾和尿素,每株分别为4 g和6 g;10月施复合肥,每株10 g;2007年2月每株施20 g桉树专用肥。采样在施肥前进行,分别于2006年4、8、10、12月和2007年2、4月进行。
1.3 样品采集取样过程中,为了减少淋溶的影响,一般在取样前72 h内没有降雨。收集的树叶样品所处的方位、叶龄、层次应尽可能的保持一致。绿叶选择中上部长势正常的稳定叶,落叶选择整株叶色已经变红或变黄的衰老叶。收集的方法,轻摇树枝,收集摇晃后脱落的叶色发生变化的叶片,这些叶片可以被认为已经和植物体相脱离,或根据凋落叶的颜色从地面上选取新鲜凋落叶。
1.4 样品处理和分析方法样品采回后,用洁净的纱布将叶片两面擦试干净,剪去叶柄、叶尖和病斑等不正常部分。然后,放在105 ℃烘箱中杀青15~30 min,然后在70 ℃烘箱烘4~6 h至干,再用植物磨碎机粉碎,过1 mm筛,连同标签一起装入密封塑料袋中,待分析其养分含量。
常量元素含量采用常规方法测定。叶片N含量采用浓H2SO4-H2O2消煮、凯氏定氮法测定,消煮液同时用以P的测定;叶片P含量采用酸溶-钼锑抗比色法测定;采用火焰光度计法测定叶片K含量,用原子吸收法测定Ca和Mg含量。
营养转移的计算式有多种,有的以面积为基数计算,亦有以质量为基数计算, 本研究采用后者进行计算,养分转移率(%)=(衰老前绿叶养分含量-落叶养分含量)/衰老前绿叶养分含量×100%(王文卿等,1999;李志安等,2000;2003)。这种计算方式假定叶子脱落前后的质量不变,尽管实际情况不完全如此,但有研究发现从绿叶到脱落叶的质量损失一般不超过10%。
2 结果与分析 2.1 叶的养分含量动态变化3种桉树幼苗的绿叶与落叶养分含量1年内的变化动态图 1可以看出,各营养元素含量在落叶和绿叶养分的变化总体趋势一致,N表现为在4—6月和8—10月有所上升,然后又下降;P和K表现在6—8月有所上升其余月份缓慢下降,K全年变化幅度大于P;Ca在4—12月变化比较平缓,12月到第2年的4月份有下降趋势;Mg在4—6月和第2年的2—4月有明显下降趋势,其余各月份有所上升。

|
图 1 叶养分含量动态变化 Figure 1 Dynamic changes of nutrient content in leaf |
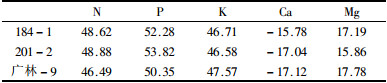
利用变异系数对不同养分全年变化的幅度进行量度结果表明(表 1):不同品种,以及同一品种的绿叶和落叶的不同元素的变异系数各不相同,对N、P、K、Mg 4种元素来说,绿叶的变异系数大于落叶的变异系数,Ca的相反,落叶的变异系数大于绿叶的变异系数。绿叶和落叶都表现为N和Mg变异系数较大,K和Ca的变异系数较小。
|
|
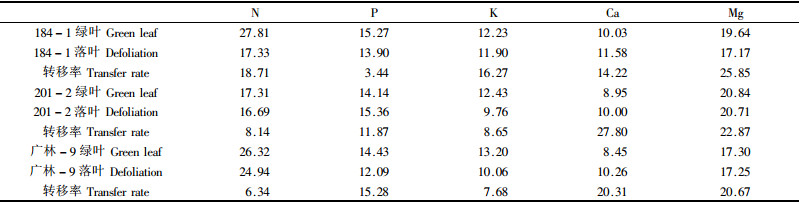
养分的变化动态表现主要与品种特性和不同的物候生长期有关。因为养分含量的变化是由叶对不同元素的不同输入输出比例引起的,叶在不同生长阶段的结构和功能直接影响到叶片对养分吸收的选择性,不同品种的叶的结构和功能又有一定的差异。对每一个品种来说,苗木生长迅速阶段,都表现为细胞具有较强的分裂能力,需要大量的蛋白质和核酸,因此对各养分的选择性吸收较多,含量表现为逐渐上升,随后由于光合作用产生的碳水化合物的增加以及养分的转移引起稀释效应;不同营养元素又表现不一致,因为不同的元素的季节动态受到其在细胞内存在状态和功能的严格制约,同时与苗木的生长需肥特性有关,另外含量下降并不能由稀释效应完全解释,特别是在降雨量比较大的生长季节,如在生长后期(8—12月),由于K的高度流动性,受降水的淋溶作用和其他器官对养分的争夺而表现为含量持续下降;Ca前期的逐渐上升可能与它的不转移和逐渐积累有关;同样Mg 6月后表现持续上升,也与它在生长后期及落叶前后有一定积累有关。
对养分含量及其变化进行方差分析结果表明(表 2):各月份之间,5种养分含量差异达到极显著水平, N、P、K、Mg各营养元素绿叶中的含量都极显著高于落叶的养分含量,Ca是落叶中养分含量显著高于绿叶,这是树叶脱落前N、P、K、Mg养转移而Ca不发生转移的结果。各品种间之间Ca含量差异显著,K和Mg的含量差异极显著,N和P含量的差异不显著,具体是,184-1的N和Ca含量最高,其中Ca含量显著高于其他2个品种;201-2的P、K、Mg含量最高,其中P的含量显著高于广林-9,K和Mg含量极显著高其他2品种;广林-9的N处于3品种之间外,其他4种元素含量均为最低,各品种同等条件下对各养分元素的吸收能力和吸收量也不同,3个品种相比201-2的吸收能力最强,广林-9的相对较差,184-1的介于两者之间,这与它们的品种特性有关。
|
|
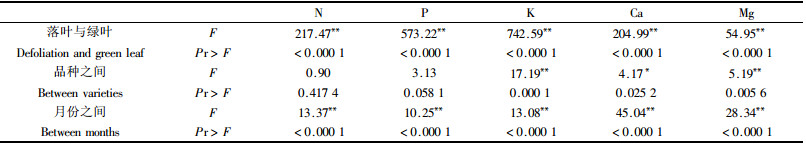
落叶中许多大量元素养分的含量显著低于绿叶养分,一般认为这是植物转移养分现象。由于苗木前期落叶量较少,所以本结果是施肥与不施肥的平均值。从表 3看,具有明显转移现象的4种元素,转移率是不同的,P的转移率最高,Mg的转移率最低,N、P、K、Mg 4种元素的转移率差异达到极显著(F=192.19,P < 0.01),即N、P、K 3种元素差异显著,它们分别极显著高于Mg的转移率。针对3个桉树品种,184-1的N、K、Mg的转移率较最高,201-2的P的转移率最高,而Mg的转移率最低,广林-9 N、P、K的转移率最低,但3个品种的转移率差异不显著,同一品种对不同营养元素的相对转移率不同,不同品种对同一营养元素的养分转移也有一定的差异。
|
|
养分转移率的变化见图 2,从图可以看出N的转移率在4—6月处于上升趋势,6—12月一直具有相对较高的转移率。2个品种又有所差异,184-1在8月份转移率最高;201-2在6月份到第2年的2月转移率表现最高;广林-9全年变化幅度较小。说明养分转移率的变化一方面与苗木生长季节有关,苗木在生长相对较慢的时期,需要的养分减少,这时期N的转移率也低,而在生长较快的时期,有机物迅速积累,需要大量营养元素满足生长发育的需要,所以这时期N元素转移率呈上升趋势;另一方面与品种特性有关,即不同品种在各个季节内的养分转移率也不同。

|
图 2 叶片养分相对转移率 Figure 2 Relative nutrient transfer rate in leaf |
P的转移率出现先上升后下降的趋势,春季到夏季整个过程,苗木生长逐渐加快,P元素的需要量也逐渐上升,直至P元素的营养临界期,这时期植株需要P元素合成有机物来构建营养器官,这个时期P元素需要比较迫切,所以它的转移率升高。随后7—12月,P元素的转移率呈下降趋势,由于P元素的移动性比N、K的小,所以P的转移率下降幅度小于K转移率下降幅度。
K的转移率与P的变化基本一致,单峰曲线,K的转移率在秋冬季节下降幅度大于磷,且201-2和广林-9在6月份就开始下降,这可能与桉树苗木生长前期(1—5月)需K量相对较少有关,另外可能受K的高度流动性和降水淋溶的影响。
Ca的是负值,全年波浪式变化,表明Ca不发生转移,其变化的大小主要与绿叶片的含量和苗木的生长特性有关。Mg的转移率较小,3个桉树品种均表现为4—6月和8—12月下降,6—8月和12月至次年4月上升趋势,这与单位叶面积叶绿素含量有关,因为Mg是叶绿素合成的重要元素。
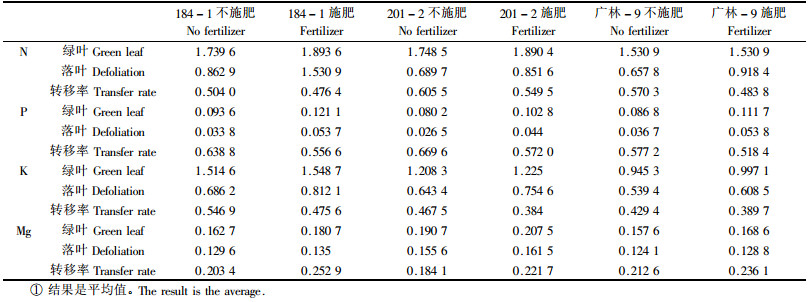
2.3 施肥对叶片养分及养分转移的影响一般认为养分含量与土壤有效养分含量呈正相关,依据这种相关性,再根据绿叶与落叶中养分含量的相关性大小,即可判断土壤养分含量对叶养分转移率的影响。对施肥与不施肥叶片养分含量和养分转移率的比较结果表 4可以看出,施肥苗木绿叶养分含量显著高于不施肥叶片的养分含量(t=2.97>t 0.025(6)=2.446 9);施肥落叶的养分含量也显著高于不施肥的叶养分含量(t=2.24>t 0.05(6)=1.943 2)。有关养分转移率,施肥苗木叶片N、P、K的养分转移率反而低于不施肥处理;对Mg的转移率来说,施肥处理高于不施肥处理,差异也达显著水平(t=2.77>t 0.025(6)=2.446 9)。进一步相关分析结果表明,落叶养分含量与绿叶养分含量正相关(相关系数r=0.94,p < 0.01),叶养分转移率与叶养分含量相关不显著。以上结果说明施肥对叶片养分含量影响显著,但对叶片养分转移率影响不显著。
|
|
叶片养分元素含量可作为植物养分供应水平的一个指标。叶片衰老过程中由于呼吸消耗及碳水化合物、核酸、脂类、蛋白质等降解后小分子物质外运,使叶片的质量及N、P、K等元素含量下降,因此叶养分含量的变化是植物遗传因素和植物物候期及外界环境共同作用的结果。184-1、201-2、广林-9等3种桉树幼苗绿叶N、P、K、Mg的养分含量及其变化系数显著高于落叶,且成正相关,Ca相反;3个品种中N、P、K、Ca、Mg含量表现分别是N水平:184-1>广林-9>201-2;P、K、Mg水平:201-2>184-1>广林-9;Ca水平:184-1>201-2>广林-9。
叶的养分转移在生态系统中的重要意义已被许多研究所证实(曾德慧,2005)。许多研究(Gime, 1979;Chapin, 1980;Berendse et al., 1987;Aerts,1990;May et al., 1992)表明,在叶片衰老过程中养分转移减少了由于凋落物而引起的养分损失,低的养分损失率能够使植株种群保持健康生长,减少了植物对土壤养分吸收的依赖性(Aert, 1996;Van Heerwaarden et al., 2003)。试验条件下184-1、201-2、广林-9等3种桉树幼苗养分转移率及其变化动态表明:Ca为非转移元素;N、P、K、Mg 4种营养元素则表现为P>N>K>Mg,4种元素差异达到极显著(F=192.19,P < 0.01);对单个品种来说,184-1的N、K、Mg的转移率最高;201-2的P的转移率最高,Mg的转移率最低;广林-9 N、P、K的转移率最低,但3个品种的转移率差异不显著。
就施肥与绿叶片含量及其在叶片衰老过程中转移率的关系而言,施肥桉树叶片养分含量显著高于不施肥叶片养分含量(t=2.97>t 0.025(6)=2.446 9),施肥条件下养分转移率比不施肥的低。这与Chapin(1980)的研究结果有相同之处。土壤养分含量低者(即生长于贫瘠土壤中者)的转移率反而高,其原因可能是可溶性蛋白的含量相对较高。但这方面的研究至今没有定论,还有待进一步深入研究。
李志安, 王伯荪. 2000. 植物营养转移研究进展. 武汉植物学研究, 18(3): 229-236. DOI:10.3969/j.issn.2095-0837.2000.03.011 |
李志安, 林永标, 彭少麟. 2000. 华南人工林凋落物养分及其转移. 应用生态学报, 11(3): 321-326. DOI:10.3321/j.issn:1001-9332.2000.03.001 |
李志安, 邹碧, 曹裕松, 等. 2003. 华南两种豆科人工林体内养分转移特性. 生态学报, 23(7): 1395-1402. DOI:10.3321/j.issn:1000-0933.2003.07.018 |
廖利平. 1994. 国外林木养分内循环研究. 生态学杂志, 13(6): 34-38. DOI:10.3321/j.issn:1000-4890.1994.06.003 |
孙书存, 陈灵芝. 2001. 东灵山地区辽东栎叶养分的季节动态与回收效率. 植物生态学报, 25(1): 76-82. DOI:10.3321/j.issn:1005-264X.2001.01.013 |
王文卿, 林鹏. 1999. 树木叶片衰老过程中养分元素内吸收研究. 武汉植物学研究, 17(增): 117-122. |
王希华, 黄建军, 闫恩荣. 2004. 天童常绿阔叶林若干树种的叶片营养转移研究. 广西植物, 24(1): 81-85. DOI:10.3969/j.issn.1000-3142.2004.01.020 |
徐大平, 张宁南. 2006. 桉树人工林生态效应研究进展. 广西林业科学, 35(4): 179-187. DOI:10.3969/j.issn.1006-1126.2006.04.002 |
薛立, 徐燕, 吴敏, 等. 2005. 4种阔叶树种叶中氮和磷的季节动态及其转移. 生态学报, 25(3): 251-256. |
杨曾奖, 李尚均, 徐大平, 等. 2006. 钾素营养对浅海沉积土桉树生长的影响. 生态环境, 15(2): 337-340. DOI:10.3969/j.issn.1674-5906.2006.02.029 |
余雪标, 李维国, 王尚明, 等. 2000.桉树人工林的若干生态问题及其研究进展//余雪标.桉树人工林长期生产力管理研究.北京: 中国林业出版社, 19-26.
|
曾德慧, 陈广生, 陈伏生, 等. 2005. 不同林龄樟子松叶片养分含量及其再吸收效率. 林业科学, 41(5): 21-27. DOI:10.3321/j.issn:1001-7488.2005.05.004 |
Aerts R. 1990. Nutrient use efficiency in evergreen and deciduous species from heathlands. Oecologia, 84: 391-397. DOI:10.1007/BF00329765 |
Aerts R. 1996. Nutrient resorption from senescing leaves of perennials: are there general patterns. J Ecol, 84: 597-608. DOI:10.2307/2261481 |
Berendse F, Aerts R. 1987. Nitrogen use efficiency: a biologically meaningful definition. Funct Ecol, 1: 293-296. |
Chapin F S Ⅲ. 1980. The mineral nutrition of wild plants. Ann Rev Ecol Syst, 2: 233-260. |
Grime J P. 1979. Competition and the struggle for existence//Anderson B M, Turner B D, Taylor L R. Population dynamics, Proc 20th Symp B E S. Oxford: Blackwell Scientific Publications, 123-139.
|
May J D, Killingbeck K T. 1992. Effects of preventing nutrient resorption on plant fitness and foliar nutrient dynamics. Ecology, 73: 1868-1878. DOI:10.2307/1940038 |
Van Heerwaarden L M, Toet S, Aerts R. 2003. Nitrogen and phosphorus resorption efficiency and proficiency in six subarctic bog species after 4 years of nitrogen fertilization. J Ecol, 91: 1060-1070. DOI:10.1046/j.1365-2745.2003.00828.x |