 2008, Vol. 44
2008, Vol. 44文章信息
- 邓文红, 沈应柏, 李镇宇.
- Deng Wenhong, Shen Yingbai, Li Zhenyu
- 马尾松毛虫取食、茉莉酸甲酯和萜烯混合物处理对马尾松18种氨基酸含量的影响
- Effects of Dendrolimus punctatus, MeJA and Terpenes-Treatment on theContents of Eighteen Amino Acids in Pinus massoniana Seedlings
- 林业科学, 2008, 44(10): 69-75.
- Scientia Silvae Sinicae, 2008, 44(10): 69-75.
-
文章历史
- 收稿日期:2008-03-03
-
作者相关文章



2. 北京林业大学生物科学与技术学院 北京 100083;
3. 北京林业大学森林培育与保护教育部重点实验室 北京 100083
2. College of Biological Science and Biotechnology, Beijing Forestry University Beijing 100083;
3. Key Laboratory of Forest Silviculture and Conservation of Ministry of Education, Beijing Forestry University Beijing 100083
植物受到害虫危害后会产生一系列生理生化反应(Bristow, 1989;戈峰, 1992),通过调节体内化学物质的数量和组成,产生诱导化学防御(Haukioja, 1990;Karban et al., 1993;Leather et al., 1987;康乐, 1995;李镇宇等, 2000;娄永根等, 1997)。如松粉蝶(Neophasia menajpia)取食松树后会使植物体内的松粉蝶营养物质含量下降,导致取食后的松粉蝶营养不良(李镇宇等, 1998)。氨基酸是昆虫生长发育的主要营养物质,食物中蛋白质的营养价值主要决定于所含氨基酸的种类和比例(Harvey,1974)。李镇宇等(1998)研究了油松(Pinus taebulaeformis)受赤松毛虫危害后,发现油松针叶中游离氨基酸总量明显减少,用受害油松针叶饲养的幼虫摄食量和产卵量均减少,死亡率升高;而昆虫的组织在整个发育过程中对氨基酸的需要是处于变动状态的,因此,可以认为氨基酸含量的大幅度变化和总量的减少是松树诱导抗性的一个方面(李镇宇等, 2000)。昆虫体内GABA是一种重要的抑制性神经递质,植食性昆虫的活动对植物叶子的机械刺激和破坏可以诱导GABA的合成和积累,植物组织快速积累GABA可以影响植食性昆虫的摄食习性(Ramputh et al., 1996;Wallace et al., 1984)。研究发现(Farmer et al., 1992; Hayden et al., 2002; Stratmann et al., 1997; Wierstra et al., 2000; 娄永根等, 2000; 任琴等, 2006b),植物在受到伤害后不仅在受伤部位产生防御反应,在伤口远端的组织中也迅速产生类似的反应,使整株植物产生系统诱导抗性,阻止昆虫的进一步危害。
马尾松毛虫(Dendrolimus punctatus)是我国南方主要造林树种马尾松(Pinus massoniana)的重要害虫,也是我国林业上发生面积最大、危害最重的食叶害虫,在我国周期性暴发成灾,戈峰等(2002)、任琴等(2005;2006a)及马尾松抗松毛虫抗性研究组(1990)等曾对马尾松的诱导抗性进行研究,但马尾松毛虫对马尾松苗木体内GABA和17种游离氨基酸含量的系统性影响尚未见报道。本文以盆栽马尾松为试验材料,研究虫害、10 μmol·L-1茉莉酸甲酯、10 μmol·L-1萜烯混合物熏蒸马尾松苗4 h后,马尾松损伤枝、姊妹枝和系统枝中GABA和17种游离氨基酸含量变化的规律,旨在了解GABA和17种游离氨基酸在树木伤害防御反应中的作用,并为进一步揭示MeJA和萜烯类化合物作为伤信号分子的作用机制提供理论依据。
1 材料与方法 1.1 材料试验所用树种为盆栽马尾松,来自江西省高安市, 盆栽马尾松每2周浇1次1/2Hoagland营养液,防止营养不良, 育苗过程中经常浇水,避免产生干旱胁迫,影响试验结果;马尾松毛虫由中国科学院动物研究所提供,5龄幼虫,生长良好。
1.2 样品采集选取树龄相同、长势良好且具有6个轮枝的盆栽马尾松3株作标记,分别接入一定量的马尾松毛虫幼虫,取食1 d后造成轻度伤害(针叶损失量达25%~40%),待达到所要求的受害程度后,移去马尾松毛虫幼虫,开始计时。
选取与受虫害一致的盆栽马尾松2株,用10 μmol·L-1茉莉酸甲酯熏蒸马尾松健康植株,并计时。
选取与受虫害一致的盆栽马尾松2株作标记,用10 μmol·L-1萜烯(α-蒎烯、β-蒎烯、柠檬烯、3-蒈烯、月桂烯)混合物熏蒸马尾松健康植株,并计时。
同时设置3株树作为非危害对照,在处理后的4 h收集处理枝、姊妹枝、系统上枝、系统下枝4个部位的马尾松针叶。采样完毕迅速将针叶投入液氮中,-80 ℃保存备用。
1.3 氨基酸的提取准确称取0.5 g马尾松针叶鲜样,用70%乙醇研磨成匀浆,4 ℃冰箱中提取过夜,离心,上清液用0.1 mol·L-1 HCl调pH值为2.0,用等体积的石油醚:乙醚萃取3次,弃去有机相,水相60 ℃条件下旋转蒸发至干,用500 μL0.01 mol-1HCl溶解样品,过0.45 μm微孔滤膜,待上HPLC。
1.4 氨基酸含量的测定 1.4.1 仪器美国Agilent公司HP1100高效液相色谱仪,系统配置为G1315B二极管阵列检测器,G1316A柱温箱,G1313自动进样器,G1311A四元泵,G1322A在线脱器机以及Agilent色谱工作站。
1.4.2 试剂17种氨基酸标准品(天门冬氨酸、谷氨酸、丝氨酸、组氨酸、甘氨酸、苏氨酸、丙氨酸、精氨酸、酪氨酸、胱氨酸、缬氨酸、蛋氨酸、苯丙氨酸、异亮氨酸、亮氨酸、赖氨酸、脯氨酸)、邻苯二甲醛、9-芴基甲基氯甲酸酯以及pH10.4的0.13 mol·5L-1硼酸缓冲液,购自Agilent公司;γ-氨基丁酸购自Sigma公司;醋酸钠、三乙胺、四氢呋喃为国产分析纯;甲醇、乙腈为美国Fisher公司生产;水为去离子水用MILLI-Q(美国Millipore公司)超纯水仪处理。
1.4.3 色谱条件色谱柱:Agilent Hypersil AA-ODS, 5 μm 200 mm×2.1 mm柱和保护柱; 流动相A:20 mmol·5L-1 NaAc+0.018%TEA, 用1%~2%醋酸调pH至7.2;流动相B:20%的20 mmol·5L-1 NaAc,用1%~2%醋酸调pH至7.2+40%乙腈和40%甲醇; 流速:0.45 mL·min-1, 梯度洗脱,见表 1; 柱温:40 ℃; DAD检测器:信号=338/10 nm,参考=390/20 nm,16min时信号=262/16 nm,参考=324/8 nm。
|
|
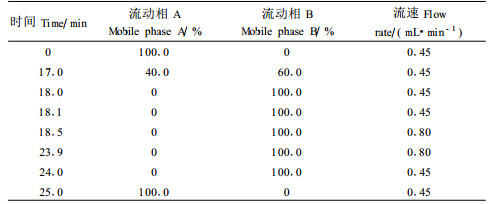
采用自动进样器进行OPA-FMOC柱前衍生,吸取硼酸缓冲液5 μL后,吸取1 μLOPA,洗针,吸取1 μL氨基酸标准溶液或样品,混合,洗针,吸取1 μLFMOC,洗针,混合后进样。
2 结果与分析 2.1 伤害对马尾松针叶中氨基酸总量的影响 2.1.1 虫害后马尾松针叶中氨基酸总量的系统性变化与对照相比,马尾松被马尾松毛虫危害后,其受害针叶、姊妹针叶、系统上枝针叶和系统下枝针叶中氨基酸总量均降低(表 2)。说明马尾松枝条受到马尾松毛虫危害后,不仅受害部位氨基酸含量减少,使自己免受进一步伤害,还会把受害信息传递给系统枝条针叶,产生系统诱导抗性,从而引起系统枝条针叶中氨基酸含量的降低。
|
|
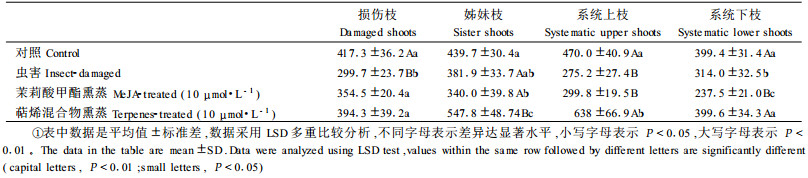
10 μmol·L-1茉莉酸甲酯熏蒸处理马尾松苗木4 h后,其熏蒸针叶、姊妹针叶和系统针叶中氨基酸总量与马尾松虫害后的结果相同,均呈下降趋势(表 2)。说明茉莉酸甲酯可以代替虫害等外部刺激,启动相似的信号防御系统,诱导健康植株针叶中氨基酸含量的降低,产生系统诱导抗性,起到防御外界伤害的作用。
2.1.3 萜烯混合物熏蒸后马尾松针叶中氨基酸总量的系统性变化10 μmol·L-1萜烯混合物熏蒸处理马尾松苗木4 h后,熏蒸枝和系统下枝中氨基酸总量略有下降,而姊妹枝和系统上枝中氨基酸含量升高(表 2)。说明萜烯混合物虽然可以代替虫害等外部刺激,启动相似的信号防御系统,产生系统诱导抗性。但是它与虫害和茉莉酸甲酯熏蒸处理之间的作用又有些不同,而且植物体能够区分,从而产生不同的反应。
2.2 伤害对马尾松针叶中各种氨基酸含量的影响 2.2.1 虫害后马尾松针叶中各种氨基酸含量的系统性变化从图 1可以看出,马尾松被马尾松毛虫取食后,虫害枝、姊妹枝、系统上枝和系统下枝中天门冬氨酸、谷氨酸、组氨酸、赖氨酸含量全部下降;丝氨酸、甘氨酸、苏氨酸、γ-氨基丁酸、酪氨酸、脯氨酸含量升高;胱氨酸、异亮氨酸、亮氨酸除系统下枝含量升高外,其他降低,精氨酸姊妹枝中含量升高,而在其他3个部位中全部降低,其他氨基酸在所测4个部位中表现出有升有降的趋势。LSD多重比较显示(P<0.05),虫害后天门冬氨酸、谷氨酸、丝氨酸、甘氨酸、苏氨酸、γ-氨基丁酸、胱氨酸在虫害针叶、姊妹针叶和系统针叶中含量变化差异显著;精氨酸、酪氨酸和苯丙氨酸除虫害后系统上枝不显著外,其他部位含量变化差异显著。说明虫害对氨基酸含量的变化有显著影响。

|
图 1 不同处理的马尾松针叶中氨基酸含量的系统性变化 Figure 1 Systematic changes of amino acids contents in needles of P. massoniana by different treatments |
10 μmol·L-1茉莉酸甲酯熏蒸处理马尾松苗木4 h后,在所检测的18种氨基酸中有8种氨基酸的变化规律与马尾松虫害后的结果相同。其中天门冬氨酸、谷氨酸和赖氨酸含量在熏蒸枝、姊妹枝、系统上枝和系统下枝中全部下降;胱氨酸除系统下枝含量升高外,其他部位降低;丝氨酸、甘氨酸、苏氨酸、γ-氨基丁酸含量升高。其他氨基酸的变化规律与虫害略有不同。LSD多重比较结果表明(P<0.05),天门冬氨酸、谷氨酸、丝氨酸、甘氨酸、苏氨酸、γ-氨基丁酸、酪氨酸、苯丙氨酸在虫害针叶、姊妹针叶和系统针叶中含量变化差异显著;胱氨酸、亮氨酸和脯氨酸除系统下枝针叶中变化不显著外,其他部位含量变化差异显著。说明马尾松能够识别虫害和熏蒸,而且其受害机制也可能不同。MeJA熏蒸只能模拟虫害的结果,并不能模拟虫害的过程,MeJA作为创伤信号的传递体之一,在植物创伤中参与了局部和系统的抗性(Doke et al., 1991; Orozco-Cardenas et al., 1999)(图 1)。
2.2.3 萜烯混合物熏蒸后马尾松针叶中各种氨基酸含量的系统性变化10 μmol·L-1萜烯混和物熏蒸处理马尾松苗木4 h后,在所检测的18种氨基酸中有6种氨基酸的变化规律与马尾松虫害后的结果相同。其中天门冬氨酸和谷氨酸含量在熏蒸枝、姊妹枝、系统上枝和系统下枝中全部下降;丝氨酸、甘氨酸、苏氨酸和γ-氨基丁酸含量升高。LSD多重比较结果表明(P<0.05),天门冬氨酸、丝氨酸、甘氨酸、苏氨酸、精氨酸、γ-氨基丁酸、酪氨酸、胱氨酸、蛋氨酸、苯丙氨酸在虫害针叶、姊妹针叶和系统针叶中含量变化差异显著;谷氨酸和赖氨酸除姊妹枝中变化差异不显著外,其他部位含量变化差异显著。说明作为针叶树主要抗虫物质的萜烯类化合物,当植物遭受损伤后可能作为细胞内应答伤信号的信号分子,起到保护健康植株免受伤害的预先报警信号,产生系统诱导抗性(图 1),但与茉莉酸甲酯熏蒸处理的结果有所不同。
3 讨论氨基酸是昆虫生长发育不可缺少的生物活性物质,除参与蛋白质合成外,还具有缓冲调节血液渗透压、参与能量代谢和脂肪酸合成等功能,其含量过低或各种氨基酸的相对比例不均衡,都会影响生物的正常生长发育和繁衍(Harvey,1974;彩万志等,2001;王荫长,2001)。研究表明,马尾松受到轻度虫害4 h后,虫害针叶、姊妹针叶和系统针叶中氨基酸总量明显降低,说明马尾松被马尾松毛虫咬食后,伤口部位迅速产生了某种信号分子,由它把受伤信息传递给姊妹枝条和系统枝条针叶,启动植物体内一系列防御基因的表达,诱导健康植株针叶中游离氨基酸总量降低,产生系统诱导抗性,使松毛虫取食后得不到正常生长发育所需要的足够营养,从而防止进一步被伤害。GABA是一种广泛存在于各种植物中的四碳非蛋白质氨基酸,在信号转导和防御反应中有重要作用,笔者首次研究了马尾松受害后GABA含量的变化情况,结果表明,虫害后虫害针叶、姊妹针叶和系统针叶中GABA含量明显增加,Shelp等(1999)报道,植物在逆境胁迫下会迅速积累GABA,反过来摄食GABA又能破坏昆虫的正常生长和发育。同时由于GABA作为一种可以移动的小分子氨基酸,推测GABA可能作为细胞内应答伤信号的信号分子,起到保护健康植株免受伤害的预先报警信号,产生系统诱导抗性,防止进一步被伤害。
与对照相比,马尾松被马尾松毛虫取食后,天门冬氨酸、谷氨酸、组氨酸、赖氨酸虫害针叶、姊妹针叶和系统针叶中含量均降低,胱氨酸、异亮氨酸、亮氨酸在虫害针叶、姊妹针叶和系统上枝针叶中降低,精氨酸在虫害针叶和系统针叶中降低;尤以天门冬氨酸和谷氨酸降低最多,据报道,谷氨酸和天门冬氨酸由于在转氨作用中处于中心地位,在昆虫中可作为补充营养(管致和, 1996);异亮氨酸、亮氨酸为昆虫体内的必需氨基酸(戈峰等,1997);赖氨酸、组氨酸与生物碱的合成有关,而生物碱已被证明是重要的抗虫物质(罗万春等,1997);精氨酸是合成植物多胺的前体物质,胁迫条件下细胞内大量积累植物多胺(汪沛洪,1990)。各种氨基酸含量的变化可能与这些生物过程紧密相关。丝氨酸、甘氨酸、苏氨酸、γ-氨基丁酸、酪氨酸和脯氨酸含量升高,已证明丝氨酸、酪氨酸和脯氨酸对棉蚜虫不利(钦俊德,1987;刘旭明等,1993;邹运鼎,1991);甘氨酸和丙氨酸与柑桔(Cirtus reticulata)的抗蚜性呈明显的正相关(叶兴祥等,1993)。马尾松苗木系统诱导抗性与氨基酸各组分更进一步的关系,则有待于深入的研究。
近年来,MeJA作为创伤信号分子的研究备受关注,经常被用来模拟昆虫取食,研究植物的诱导抗性(Baldwin et al., 1997;McConn et al., 1997;Orozco-cardenar et al., 2001)。萜烯类是近年来研究较多的一类重要的树木抗虫物质,在针叶树中尤为重要。从氨基酸含量变化的情况来看,10 μmol·L-1外源MeJA和10 μmol·L-1萜烯混合物熏蒸虽然可以代替虫害等外界刺激启动防御系统,产生系统诱导抗性。但是3种不同处理方式引起的氨基酸含量的变化规律不完全相同,推测不同处理方式可能启动了不同的生物合成途径。由于各种原因,本文只对马尾松受害4 h的受害针叶、姊妹针叶、系统上枝针叶和系统下枝针叶中氨基酸的含量进行了测定和分析,而没能进行时间动态的测定,这还有待于进一步研究。
彩万志, 庞雄飞, 花保祯, 等. 2001. 普通昆虫学. 北京: 中国农业大学出版社, 127.
|
戈峰. 1992. 昆虫对植物的胁迫作用//万方浩, 夏云龙. 青年生态学者论从(二). 北京: 中国科学技术出版社, 87-91.
|
戈峰, 李典谟, 邱业先, 等. 1997. 松树受害后一些化学物质含量的变化及其对马尾松毛虫种群参数的影响. 昆虫学报, 40(4): 337-342. DOI:10.3321/j.issn:0454-6296.1997.04.001 |
戈峰, 李镇宇, 谢映平, 等. 2002. 我国主要松树诱导抗虫性的一些规律比较. 北京林业大学学报, 24(3): 61-65. DOI:10.3321/j.issn:1000-1522.2002.03.013 |
管致和. 1996. 植物医学导论. 北京: 中国农业大学出版社.
|
康乐. 1995. 环境胁压下的昆虫-植物相互关系. 生态学杂志, 14(5): 51-57. |
李镇宇, 陈华盛, 袁小环, 等. 1998. 油松对赤松毛虫的诱导化学防御. 林业科学, 34(2): 43-50. DOI:10.3321/j.issn:1001-7488.1998.02.007 |
李镇宇, 王燕, 陈华盛, 等. 2000. 油松对赤松毛虫的诱导化学防御及滞后诱导抗性. 林业科学, 36(1): 66-70. DOI:10.3321/j.issn:1001-7488.2000.01.011 |
刘旭明, 杨奇华. 1993. 棉花抗蚜的生理生化机制及其与棉蚜种群数量消长关系的研究. 植物保护学报, 20(1): 25-29. DOI:10.3321/j.issn:0577-7518.1993.01.020 |
娄永根, 程家安. 1997. 植物的诱导抗虫性. 昆虫学报, 40(3): 320-330. DOI:10.3321/j.issn:0454-6296.1997.03.018 |
娄永根, 程家安. 2000. 虫害诱导的植物挥发物:基本特性、生态学功能及释放机制. 生态学报, 20(6): 1097-1106. DOI:10.3321/j.issn:1000-0933.2000.06.031 |
罗万春, 慕立义, 李云寿. 1997. 植物源生物碱的杀虫作用. 农药, 36(7): 11-15. |
马尾松抗松毛虫抗性研究组. 1990. 马尾松抗松毛虫植株的抗性机制研究. 林业科学, 26(2): 133-140. |
钦俊德. 1987. 昆虫与植物的关系. 北京: 科学出版社, 38-61.
|
任琴, 李镇宇, 胡永建, 等. 2005. 受害马尾松、湿地松挥发性化学物质的释放. 生态学报, 25(11): 2928-2932. DOI:10.3321/j.issn:1000-0933.2005.11.019 |
任琴, 金幼菊, 胡永建, 等. 2006a. 马尾松诱导挥发性有机化合物的快速变化. 林业科学, 42(4): 65-70. |
任琴, 杨莉, 胡永建. 2006b. 受害马尾松针叶内脱落酸含量的变化. 北京林业大学学报, 28(5): 99-101. |
汪沛洪. 1990. 植物多胺代谢的酶类与胁迫反应. 植物生理学通讯, (1): 1-7. |
王荫长. 2001. 昆虫生物化学. 北京: 中国农业出版社, 60, 92-96.
|
叶兴祥, 叶琪明, 王强, 等. 1993. 柑桔对桔棉蚜的抗性与其叶片游离氨基酸含量的关系. 浙江农业学报, 5(3): 184-186. |
邹运鼎, 杨义和. 1991. 棉花苗期棉株内含物与棉蚜种群消长动态及蚜翅型分化的关系. 昆虫知识, 28(1): 14-17. |
Baldwin I T, Zhang Z P, Diab N. 1997. Quantification, correlations and manipulations of wound-induced changes in jasmonic acid and nicotine in Nicotiana sylvestris. Planta, 201: 397-404. DOI:10.1007/s004250050082 |
Bristow C M. 1989. Host development offers new insight into insect plant interaction. Trends Ecol Evol, 5(4): 123-124. |
Doke N, Miura Y, Chai H B, Kawakita K. 1991. Involvement of active oxygen in induction of plant defense response against infection and injury// Pell E, Steffen K. Active Oxygen/Oxidative stress and Plant Metabolism. American Society of Plant Physiologists, 184-196.
|
Farmer E E, Ryan C A. 1992. Octadicanoid precursors of JA activate the synthesis of wound-inducible proteinase inhibitors. Plant Cell, 4: 129-134. DOI:10.1105/tpc.4.2.129 |
Harvey G T. 1974. Nutritional studies of eastern spruce budworm Ⅰ.Soluble sugars. Can Entomo, 93: 594-602. |
Haukioja E. 1990. Induction of defenses in trees. Annual Review of Entomology, 36: 25-42. |
Hayden K J, Parker I M. 2002. Plasticity in cyanogensis of Trifolium repens L: inducibility, fitness costs and variable expression. Evol Ecol Res, 4: 151-168. |
Karban R, Nhho C. 1993. Induced resistance and plant density for a native shrub, Gassypium thurberi. Ecology, 74(1): 9-19. DOI:10.2307/1939496 |
Leather S R, Watt A F. 1987. Insect-induced chemical changes in young lodgpole pine(Pinus contorta):the effect of previous defoliation on oviposition, growth and survival of the pine beauty moth, Panolis flammea. Ecological Entomology, 12: 275-281. DOI:10.1111/j.1365-2311.1987.tb01006.x |
McConn M, Creelman R A, Bell E, et al. 1997. Jasmonate is essential for insect defense in Arabidopsis. Proc Natl Acad Sci USA, 94: 5473-5477. DOI:10.1073/pnas.94.10.5473 |
Orozco-Cardenas M, Ryan C A. 1999. Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the oct adecanoid pathway. Proceedings of the National Academy of Sciences, USA, 96: 6553-6557. DOI:10.1073/pnas.96.11.6553 |
Orozco-Cardenas M L, Narvaez-Vasquez J, Ryan C A. 2001. Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in reponse to wounding, systemin and methyl jasmonate. Plant Cell, 13: 179-191. DOI:10.1105/tpc.13.1.179 |
Ramputh A I, Bown A W. 1996. Rapid r-aminobutyric acid synthesis and the inhibit ion of the growth and development of oblique-banded leaf-roller larvae. Plant Physiol, 111: 1349-1352. DOI:10.1104/pp.111.4.1349 |
Shelp B J, Bown A W, Mclean M D. 1999. Metabolism and functions of gamma-aminobutyric acid. Trends in Plant Science, 4: 446-452. DOI:10.1016/S1360-1385(99)01486-7 |
Stratmann J W, Ryan C A. 1997. Myelin basic protein kinase activity in tomato leaves is induced systemically by wounding and increase in response to systemin and loigosaccharide elicitors. Proc Natl Acad Sci USA, 94: 11085-11089. DOI:10.1073/pnas.94.20.11085 |
Wallace W, Secor J, Schrader L E. 1984. Rapid accumulation of r-aminobutyric acid and alanine in soybean leaves in response to an abrupt transfer to lower temperature, darkness, or mechanical manipulation. Plant Physiol, 75: 170-175. DOI:10.1104/pp.75.1.170 |
Wierstra I, Kloppstech K. 2000. Differential effects of methyl jasmonate on the expression of the early light-inducible proteins and other light-regulated genes in barley. Plant Physiol, 124: 833-844. DOI:10.1104/pp.124.2.833 |