 2008, Vol. 44
2008, Vol. 44文章信息
- 有祥亮, 余新晓, 张德顺, 郗金标.
- You Xiangliang, Yu Xinxiao, Zhang Deshun, Xi Jinbiao.
- 欧洲银叶椴种子快速解除休眠的技术
- Techniques of Quickly Breaking Seed Dormancy of Tilia tomentosa
- 林业科学, 2008, 44(4): 161-166.
- Scientia Silvae Sinicae, 2008, 44(4): 161-166.
-
文章历史
- 收稿日期:2007-07-20
-
作者相关文章


2. 北京林业大学 北京 100083;
3. 上海建桥学院 上海 201319
2. Beijing Forestry University Beijing 100083;
3. Jian-qiao Institute of Shanghai Shanghai 201319
银叶椴(Tilia tomentosa)又名银毛椴、绒毛椴等,为椴树科椴树属植物。银叶椴主产巴尔干半岛,向北可达匈牙利及乌克兰西部,向东经土耳其延伸至叙利亚西北部(北纬约36°30′)(唐亚等,1996)。银叶椴是优良的城市绿化树种,树冠塔形,枝叶浓密,叶色暗绿,叶背面银白色,在阳光下熠熠生辉;初夏淡黄色小花馥郁芬芳,沁人心脾,秋季叶色金黄,赏心悦目,非常适合做城市行道树及公园、庭院栽植;耐修剪,适于做绿篱(Wandell,1989)。银叶椴适应能力强,是优良的水土保持、防风固沙和水源涵养等防护林树种(Burgess,1991)。银叶椴经济价值高,木材可做胶合板和家具;树皮纤维用途广泛;花香浓郁,是重要的蜜源植物;嫩叶和花制作茶等饮料;嫩叶和花茶具有止痉挛、促发汗及止痛等药效(Burgess,1991)。
一般认为椴树(Tilia spp.)种子休眠属综合性休眠,但休眠原因,没有统一观点(史锋厚等,2006)。Spaeth(1934)认为种皮是导致椴树种子休眠的重要原因;心叶椴(Tilia cordata)种子休眠是由种皮和胚休眠造成的(Heit,1977); 美洲椴(T. americana)种子休眠的原因在于胚乳中含有发芽抑制物质(Vanstone,1982)和珠心膜限制气体出入(美国农业部林务局,1984); 银叶椴是由物理休眠(physical dormancy)和生理休眠形成的复合休眠类型(Baskin et al., 2001)。
针对椴树种子休眠问题,许多学者提出了不少破眠方法。如欧洲椴(T.europaea)去掉果皮;心叶椴种子酸蚀(或暖层积)和低温层积处理;紫椴(T.amurensis)种子混沙变温处理;美洲椴种子酸蚀、擦伤或热水浸泡,激动素(kinetin)和GA3溶液浸种而后低温层积等,皆可促进种子发芽(美国农业部林务局,1984;Heit,1977;邹学忠等,1991;杨顺等,1992;Pitel et al., 1989)。不过,这些破眠方法大多需要5个月以上时间(史锋厚等,2006)。而银叶椴在种皮具有透水性后,也需要低温层积一个冬季,第2年春天才能发芽(Heit, 1969)。
为了缩短发芽时间,提高发芽率,幼苗生长整齐、健壮,为银叶椴这一具有生态、经济、社会效益的优良植物资源在我国引种驯化成功提供苗木保障,笔者针对其种子具有的复合休眠、繁殖困难的特点,在对休眠原因进行前期探讨的基础上,深入研究快速解除种子休眠、促进萌发的苗木繁殖技术。
1 材料与方法 1.1 试验材料银叶椴种子2003年11月采于匈牙利,净度98.8%,千粒质量为43.8 g,出仁率为99.2%。
1.2 试验方法1) 种子的处理 银叶椴核果果皮厚度0.7~0.8 mm,表面有角质层,中果皮为致密木栓层,严重阻碍种子吸水膨胀。种皮表面角质化,中层致密的厚壁栅状(柱状)石细胞阻碍胚与外界的水、气交换;同时,在果皮、种皮和胚乳内积累了大量的ABA等抑制物质,使幼胚进入深度休眠状态(有祥亮,2006)。
针对银叶椴种子休眠的原因,笔者曾对快速解除种子休眠做过一些前期研究,包括保留果皮和机械去果皮的种子吸涨率对比;为增加种皮透性采用高温水烫种或浓硫酸腐蚀(分别设不同的时间梯度)进行比较研究;为解除ABA等抑制物质,分别进行不同浓度赤霉素(GA3)和细胞分裂素(6-BA)浸种试验和层积试验等(有祥亮,2006)。参考试验结果和前人研究(杨孝汉,1989;程广有等,2004),设计以下正交组合。
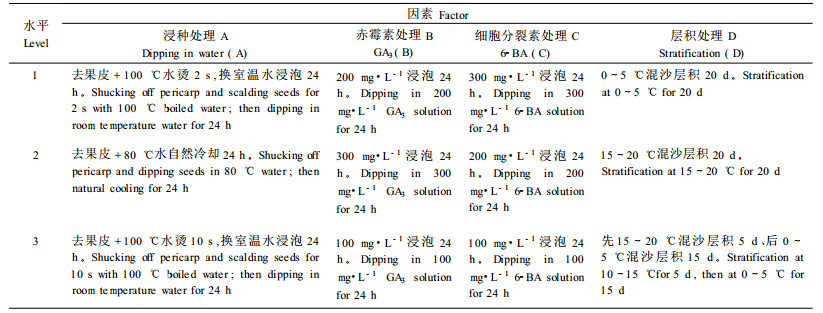
① 热水浸种处理:银叶椴果实机械去果皮,然后风选得无果皮的纯净种子。对纯净种子取出3组,每组重300 g,分别做如下处理:100℃开水速烫2、10 s,然后迅速放入室温水中浸泡24 h;80℃热水自然冷却后浸泡24 h。
② 赤霉素(GA3)处理:上述各组中取出已经吸胀的种子,按正交试验设计要求分别用100、200、300 mg·L-1赤霉素溶液,在室温下浸泡24 h。
③ 细胞分裂素(6-BA)处理:种子经赤霉素浸种处理后,按正交试验设计要求分别用100、200、300 mg·L-1细胞分裂素溶液,在室温下浸泡24 h。
④ 层积处理:种子经细胞分裂素浸种处理后,按正交试验设计要求分别进行:a种子与湿沙按1:3(体积比)的比例混匀后,装入搪瓷盘中,上覆湿纱布以保持水分,定期定量洒水和翻动,在0~5℃冰箱中放置20 d; b种子与湿沙按1:3(体积比)的比例混匀后,装入搪瓷盘中,上覆湿纱布以保持水分,定期定量洒水和翻动,在室内(15~20℃)放置20 d; c种子与湿沙按1:3(体积比)的比例混匀后,装入搪瓷盘中,上覆湿纱布以保持水分,定期定量洒水和翻动,先在室内(15~20℃)放置5 d,然后在0~5 ℃冰箱中放置15 d。
2) 试验设计 以上各组种子处理,均按正交试验设计选用正交表L27(313)(贾乃光,1993)安排试验组合。各因素和水平情况见表 1。
|
|
3) 发芽率计算、播种及出苗前后的抚育管理 层积结束后,计算各组处理的发芽率(发芽标准:长出正常胚根,幼根长为该种粒长度的一半以上)。对正交试验结果进行方差分析,找出打破休眠、促进萌发的最优方案。根据最优方案,重新取一批种子进行催芽处理,待种子有超过1/3露白时,于2004年5月28—29日播在摆放于低床上的塑料容器中,在大棚内进行育苗,60 d后调查场圃出苗率。容器基质为草炭:土=1:1(体积比),每个容器播1粒种子,播后覆盖草帘,并在大棚上覆盖遮阳网。出苗前经常洒水保持床面湿润,幼苗出土后,分次撤除草帘。
试验重复3次,以大棚内南北向低床(6 m×1 m)按长度等分为3部分,每小区2 m2,在小区内整齐排放塑料容器。
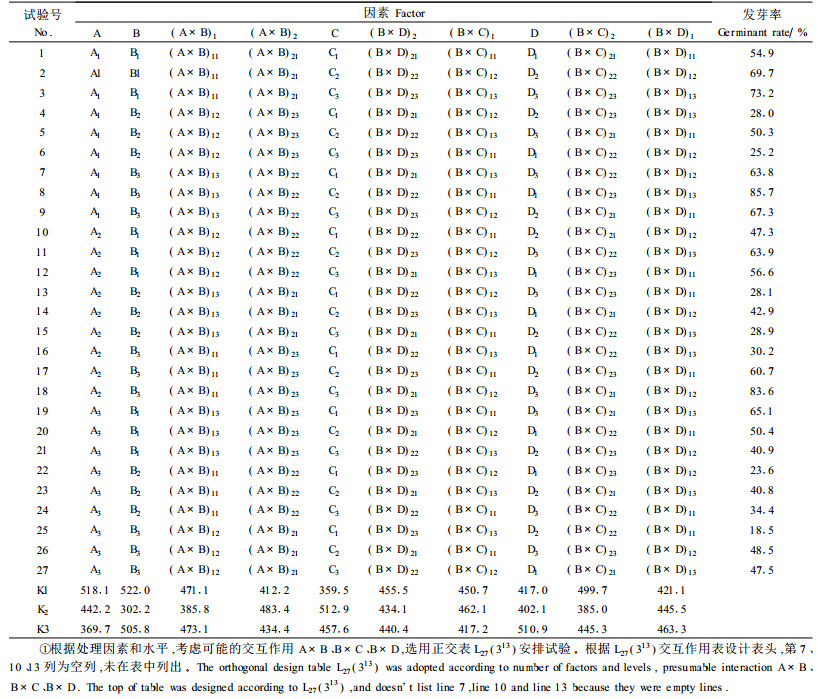
2 结果与分析 2.1 影响种子发芽率的主导因子银叶椴种子发芽正交试验结果见表 2、3。方差分析结果表明(表 3),对发芽率有极显著或显著影响的因素是赤霉素处理、细胞分裂素处理、浸种处理和层积处理,其中赤霉素处理、细胞分裂素处理的不同水平之间有极显著差异,其他2种处理的不同水平之间有显著差异。而赤霉素处理和其他3个处理的交互作用对发芽率没有显著影响。
|
|
|
|
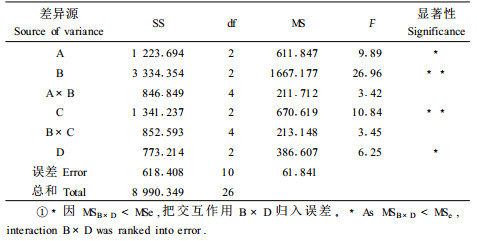
从表 2可以看出,在不考虑交互作用的情况下,发芽率最高的最佳试验因素、水平组合(优方案)应取各因素最大K值所对应的水平,即为A1B1C2D3。从方差分析的结果可以看出,MSB×D<MSe,这说明交互作用B×D对发芽率的影响较小,可以将其归入误差,交互作用A×B、B×C亦对发芽率无显著影响,所以在确定因素A、B、C、D优水平时可以不考虑交互作用,即优方案为A1B1C2D3。也就是说,浸种处理以去果皮+100℃水烫2 s,换室温水浸泡24 h(水平1)发芽率最高,赤霉素处理以200 mg·L-1溶液浸泡24 h(水平1)效果最佳,细胞分裂素处理以200 mg·L-1溶液浸泡24 h(水平2)效果最好,层积处理以先15~20℃、后0~5℃混沙层积各10 d(水平3)效果最好。当这些水平配合适当时,就为解除休眠创造有利的条件。
2.2 不同浸种处理对发芽率的影响从前面方差分析中已经知道,各种浸种处理对发芽率有显著影响。去掉果皮、100℃水烫2 s,换室温水浸泡24 h处理后,平均发芽率为57.6%,去果皮+80℃水自然冷却24 h,平均发芽率为49.1%,而去掉果皮、100℃水烫10 s,换室温水浸泡24 h处理后,平均发芽率最低,为41.1%。
2.3 不同浓度赤霉素和细胞分裂素浸种处理对发芽率的影响表 3方差分析结果表明,赤霉素浸种对发芽率有极显著的影响。赤霉素溶液浓度为100、200、300 mg·L-1各自浸泡24 h,平均发芽率分别为56.2%、58%、33%。可见不同浓度赤霉素溶液对发芽率的影响不同,较低浓度有利于种子萌发,高浓度则使发芽率下降。适宜浓度为200 mg·L-1。
同样,细胞分裂素浸种对发芽率也有极显著的影响。细胞分裂素溶液浓度为100、200、300 mg·L-1各自浸泡24 h,平均发芽率分别为50.8%、57%、39.9%。可见不同浓度细胞分裂素溶液对发芽率的影响也不同,较低浓度有利于种子萌发,高浓度则使发芽率下降。适宜浓度为200 mg·L-1。
2.4 层积处理对发芽率的影响方差分析表明,层积处理对种子发芽率有显著影响。0~5℃混沙层积20 d,15~20 ℃混沙层积20 d,先15~20℃混沙层积5 d、后0~5℃混沙层积10 d平均发芽率分别为46.3%、44.7%、56.8%。可见,不同层积方法,发芽率不同,其中以先暖温、后低温的变温层积效果最好,其最大值达到了83.6%。
2.5 场圃出苗率在获得较高种子发芽率之后,其苗木长势及保存率如何,是至关重要的问题,它决定了技术措施的生产适用性及其推广价值。
试验表明,正交试验种子催芽最优方案的田间育苗获得了较高的场圃出苗率。5月29日播种,6月11日,不到半月时间平均发芽率达到23.5%,6月21日,不到1月时间平均发芽率达到44.7%,播种2个月后的7月29日调查场圃出苗率,平均达到71.2%。可见,正交试验所确定的种子催芽最优方案出苗早,场圃出苗率高,出苗迅速而整齐。
2.6 苗木质量2004年10月19日对上述试验进行苗木质量调查,得到苗高、地径、根长、干重平均值分别为42.3、0.85、25.6和11.6 g·株-1。获得了健壮苗木。
3 结论与讨论通过以上的试验结果,经过综合分析,得到快速解除银叶椴种子休眠的综合技术为:脱去果皮、用高温水烫种增加种皮透水性、用赤霉素和细胞分裂素混合处理、先暖温后低温的变温层积。具体的催芽处理程序流程图见图 1。

|
图 1 银叶椴种子催芽处理程序图 Figure 1 Process of seed accelerative germination on Tilia tomentosa |
椴树种子被坚硬果皮包裹,去掉果壳成为解除种子休眠的第一步。Heit(1977)认为心叶椴和大叶椴果壳可以利用机械擦伤去除,欧洲椴果壳可以手工去除,美洲椴的坚硬外壳则需要用磨咖啡机处理或者酸蚀处理。由于银叶椴果皮坚韧,用手无法搓掉,可用机械擦伤果皮,再用手剥去;也可以用浓硫酸处理果皮,随着处理时间的加长,果皮逐层脱落,大约经过2 h处理,轻搓种子,果皮既与种子相分离,达到脱去果皮的目的(有祥亮,2006)。
去掉果皮的种子,可以通过酸蚀或开水烫种增加种皮的透性,浓硫酸处理以浸种30 min比较好,种子吸胀率达89%,而生活力达到87%,其他浸种时间相对差些。热水浸种,以2 s比较适宜,吸胀率达到81%,而生活力达到94%。不过,经过酸蚀后的银叶椴种子在层积过程中发芽率低且易腐烂,尤其是在暖温层积期间(有祥亮,2006),罗丽芬等(1990)处理紫椴种子,也存在这一问题。而热水烫种则很少有腐烂现象。具体原因有待深入研究。基于此,本次正交试验设计的浸种处理都采用开水浸种,而不用酸蚀。
赤霉素和细胞分裂素对尽快打破休眠、促进种子萌发有重要作用。在种子层积处理之前,利用GA3或GA3与6-BA混合浸泡东北红豆杉种子,可以使种子萌发时间提前,同时发芽率也有所提高(程广有等,2004)。本研究用GA3与6-BA先后浸泡银叶椴种子,也得出相似结果。郑光华等(1990)认为:GA3可代替需要的低温层积,解除休眠促进发芽。激素调节3因子学说认为,GA3、ABA与CK在种子萌发中起着抑制及允许萌发的作用,GA3为萌发最初所需,CK的存在归因于GA3,其作用是减缓ABA的抑制,促使GA3的作用得以实现,进而启动萌发。
在本次正交试验中,采用综合处理方法,银叶椴种子层积比单一低温层积催芽(Heit, 1969)所需发芽时间大大缩短,这是一个很突出的特点。国外随种子寄来的银叶椴催芽方法是“袋装层积法”:种子用室温水浸泡48 h,然后将种子与等量的基质进行混合,装入塑料袋,然后放入0~2℃的冷库中保存52周,最后将其放入18~20℃保存2周,种子就会发芽多且整齐。该方法实际效果不清楚,仅就其需要52周的时间才能打破休眠进行萌发,则耗时太长。而通过正交试验,能在20 d左右即能达到理想的发芽率。
综合处理方法能快速解除银叶椴种子休眠,大大缩短发芽时间,这是因为:1)去掉果皮,消除了果皮的透水障碍、内源抑制物质可能的不利影响和坚硬果皮的机械束缚。2)浸种处理,尤其是100℃开水烫2 s,再转入室温水浸泡24 h,既有较高的吸胀率(81%),又保持较高的生活力(94%)。迅速吸胀和高生活力为外源激素发挥作用打下良好基础,室温水浸泡24 h也有利于萌发抑制物质的溶解。3)外源赤霉素(GA3)和细胞分裂素(6-BA)的使用是萌发时间大为缩短的关键。外源GA3和6-BA的使用,使种子内部激素比例发生改变,由原来抑制萌发转变为有利于种胚发育。赤霉素能诱导酶的合成,促进RNA合成的同时促进酶分泌,使贮藏物质水解成可供胚生长的小分子化合物,促进萌发。因此,赤霉素可代替所需要的低温层积,尽快解除休眠促进发芽。外源细胞分裂素(CK)的使用也很重要,CK减缓ABA的抑制,促使GA3的作用得以实现,进而启动萌发。4)先暖温、后低温的变温层积为种子快速萌发创造了良好的环境条件。前期暖温有利于种子萌发所需各种酶的活化和合成,后期低温有利于贮藏物质水解伴随而来的能量或热量的释放。
程广有, 唐晓杰, 高红兵, 等. 2004. 东北红豆杉种子休眠机理与解除技术探讨. 北京林业大学学报, 26(1): 5-10. DOI:10.3321/j.issn:1000-1522.2004.01.002 |
贾乃光. 1993. 数理统计. 2版. 北京: 中国林业出版社.
|
罗丽芬, 王凤霞, 赵垦田, 等. 1990. 紫椴种子快速催芽育苗试验研究. 东北林业大学学报, 18(2): 103-108. |
美国农业部林务局. 1982. 美国木本植物种子手册. 李霆, 译. 北京: 中国林业出版社.
|
唐亚, 诸葛仁. 1996. 椴树属的地理分布. 植物分类学报, 34(3): 254-264. |
史锋厚, 卢芳, 沈永宝, 等. 2006. 椴树属植物研究进展. 林业科技开发, 20(1): 12-15. DOI:10.3969/j.issn.1000-8101.2006.01.004 |
杨顺, 林英飞. 1992. 紫椴种子变温催芽法. 内蒙古林业, (2): 28-29. |
杨孝汉. 1989. 玉兰种子休眠的解除及其生化机理研究. 华中农业大硕士学位论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y080551
|
有祥亮. 2006. 欧洲栓皮槭和银叶椴引种的适应性、繁殖技术与经济价值研究. 北京林业大学博士论文. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y958801
|
郑光华, 史忠礼, 赵同芳, 等. 1990. 实用种子生理学. 北京: 农业出版社.
|
邹学忠, 阎忠林, 刘清田, 等. 1991. 紫椴种子休眠原因及催芽处理方法的研究. 辽宁林业科技, (5): 6-10. |
Baskin C C, Baskin J M. 2001. Seeds: ecology, biogeography, and evolution of dormancy and germination. San Diego: Academic Press.
|
Burgess K S. 1991. Tilia tomentosa. The Journal of the American Association of Botanical Gardens & Arboreta, 6(1): 39-42. |
Heit C E. 1977. Propagation from seed: 27. Collecting, testing and growing tilia linden species. American Nurseryman, 146(7): 10-11. |
Heit C E. 1969. Propagation from seed:7.Germinating six hardseeded groups. American Nurseryman, 125(12): 44-45. |
Pitel J A, Cheliak W M, Wang B S P. 1989. Some biochemical changes associated with stratification and germination of basswood seeds. Seed Science and Technology, 17(1): 57-71. |
Spaeth J N. 1934. A physiological study of dormancy in Tilia seed. Cornell University Agricultural Experiment Station Memoir, 169: 1-71. |
Vanstone D E. 1982. Seed germination of American basswood in relation to seed maturity. Canadian Journal of Plant Science, 62(3): 709-713. DOI:10.4141/cjps82-103 |
Wandell W N. 1989. Sterling silver linden. Plant Patent, 65(3): 1-5. |