 2008, Vol. 44
2008, Vol. 44文章信息
- 王利, 邢世岩, 王芳, 韩克杰.
- Wang Li, Xing Shiyan, Wang Fang, Han Kejie.
- 银杏雌株种质遗传关系的AFLP分析
- AFLP Analysis on Genetic Relationship of Ginkgo biloba Female Germplasms
- 林业科学, 2008, 44(4): 48-53.
- Scientia Silvae Sinicae, 2008, 44(4): 48-53.
-
文章历史
- 收稿日期:2007-09-07
-
作者相关文章



银杏(Ginkgo biloba)系第四纪冰川之后惟一在中国保存下来的孑遗种子植物,素有“活化石”之称。孑遗之后的现代银杏,在近200万年的演化过程中,通过杂交和人工选择,银杏种子和叶片发生了明显的变异,使得银杏的种质资源相当丰富(王利等,2006a)。许多学者利用等位酶、ISSR、RAPD、AFLP等对银杏的多态性、性别标记、品种鉴定等进行了大量研究(吴俊元等,1992;王晓梅等,2001a;2001b;沈永宝等,2005;王利等,2006a;2006b)。
葛永奇等(2003)利用ISSR标记对来自5个栽培群体和可能为自然群体的66个样品的遗传多样性进行了研究,其多态带的比例达到70.45%。Kuddus等(2002)对美国宾夕法尼亚、华盛顿和尼加拉河的18株银杏间的遗传变异进行了RAPD分析,表明华盛顿群体内个体间基因组相似性很高,尼加拉河2个植株与华盛顿群体有45%差异性条带。可见,不同的银杏群体其遗传变异程度不同。
Tsumura等(1997)分析了98株银杏的等位酶变异,结果表明中国的银杏与日本的银杏亲缘关系比较近。陈月琴等(1999)比较了2个银杏个体ITS序列,结果发现银杏个体rDNA ITS序列核苷酸差异率高达25%。刘叔倩等(2001)对江苏邳州24个雌株和4个雄株类型的遗传变异进行了RAPD分析,结果表明品种间的多态带的比例仅为30.47%,另外,Shen等(2005)利用PCR-RFLP研究了银杏个体的遗传变异,这些研究表明银杏个体之间存在不同程度的变异。
目前,AFLP被认为是一种十分理想的、有效的分子标记技术,该技术具有DNA需要量少、可重复性好、多态性强、样品适用性广、遗传性稳定等特点,它可以在短时间内提供大量的信息,已被应用于植物遗传关系的研究(Cervera et al., 2005;Ferdinandez et al., 2004)。王利等(2006a;2006b)研究了29个银杏雄株种质和21个观赏品种之间的遗传关系,未见关于国内12个省与国外雌株种质遗传关系AFLP的研究报道。本研究以来自法国、日本、中国12个省的49个种质为试材,利用AFLP标记对它们的遗传关系进行研究,探讨其多态性、遗传变异,为雌株种质的分类、保护、利用等提供理论依据。
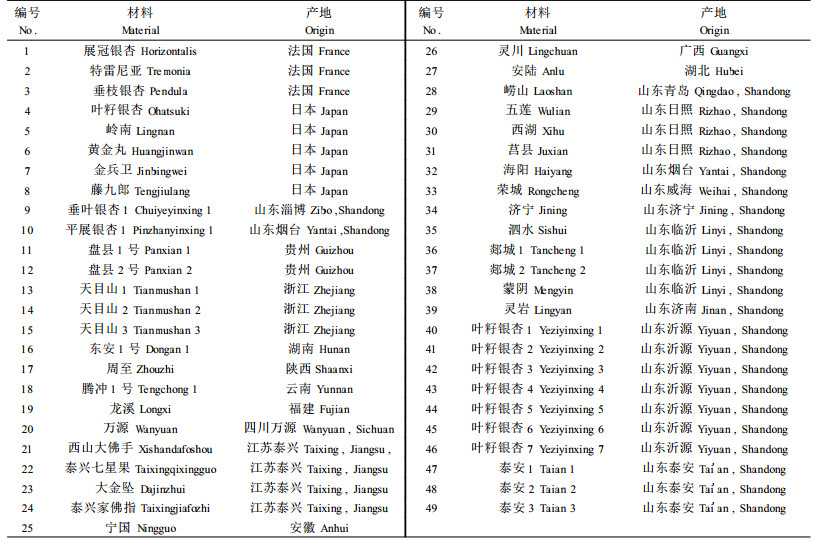
1 材料与方法 1.1 试验材料在山东农业大学的银杏种质资源圃中,采集不同种质(表 1)的幼嫩叶片,放入保鲜塑料袋中带回实验室中,先用去离子水冲洗,再用清洁的吸水纸吸干水分后,置于4 ℃冰箱中保存备用。
|
|
参照Doyle等(1990)的方法,采用CTAB法提取幼嫩银杏叶片总DNA。
1.3 AFLP分析据北京鼎国生物技术有限责任公司FISH-AFLP(EcoRI/MseⅠ型)试剂盒说明,先进行EcoRI/MseⅠ酶切与T4连接酶连接,取1 μL酶切连接液做模板,用带有1个选择性碱基的引物进行预扩增,再将预扩增液按1:15比例稀释,以2 μL预扩增稀释液为模板,用带有3个选择性碱基的引物对进行选择性扩增。变性产物(含有内标GeneScan-500)上样于4%变性聚丙烯酰胺凝胶,恒定功率50 W、最大电压3 000 V,进行电泳2.4 h,利用ABI 377测序仪对其进行检测。
1.4 数据统计分析利用GeneScan3.1软件对图像进行处理,构建0、1数学矩阵。鉴别效率(identification percentage)=能够被区分的品种数量/品种总数×100%;多态带比例PPB(percentage of polymorphic bands)=(总谱带数-有带数)/总谱带数×100%。采用NTSYS.pc2.11f计算软件,得到Dice相似系数SC(similarity coefficient)矩阵,并用非加权算术平均法(UPGMA)进行聚类分析。
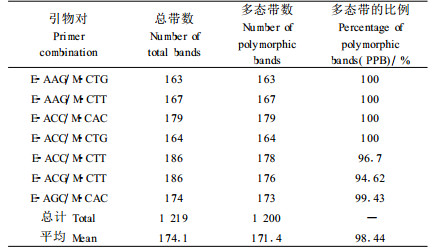
2 结果与分析 2.1 AFLP的选择性扩增结果利用筛选出的7对引物共产生1 219条谱带,多态带1 200条,多态性为98.44%。不同引物对产生的谱带数量存在一定的差异,其中,E-ACC/M-CTT、E-ACG/M-CTT引物对所产生的谱带最多(186条),而E-AAG/M-CTG引物对产生的谱带最少(163条)(表 2)。每对引物的鉴别效率为100%。
|
|
利用7对引物对49个雌株种质进行扩增,其结果表明各种质多态性存在很大差异,其平均多态性为47.80%,‘盘县1’的多态性69.92%,而‘泰安3’27.70%;其中,‘垂枝银杏’、‘岭南’、‘黄金丸’、‘盘县1’的多态性均大于60%。就叶籽银杏而言,其多态性差异很大,‘叶籽银杏5’最大(59.93%),‘叶籽银杏4’最小(33.41%),平均45.42%,小于非叶籽银杏的平均多态性48.26%。
2.3 雌株种质的特异位点对7对引物49个种质的电泳谱带分析,产生132个特异位点(包括单态带和缺失带),其中,单态带121条,缺失带11条。每个种质所产生的特异位点数量不同,‘泰安2’产生的单态带最多,12条,产生缺失带最多的是‘灵岩’,为4条。就雌株所产生的单态带而言,14个雌株种质‘垂枝银杏’、‘平展银杏’、‘盘县1’、‘东安1’、‘泰兴七星果’、‘灵川’、‘安陆’、‘崂山’、‘海阳’、‘荣城’、‘郯城2’、‘沂源叶籽银杏2’、‘莒县’、‘泰安2’的单态带数较多,4~12条,共84条单态带,占单态带总数的69.42%;3个雌株种质‘西山大佛手’、‘海阳’、‘灵岩’的缺失带较多,2~4条,它们共产生8条缺失带,占缺失带总数的72.8%。分别对雌株的特异位点进行分析,结果表明上述16个雌株的单态带或缺失带较多, 它们相对比较特殊。
2.4 相似性分析各雌株种质之间相似系数SC为0.39~0.83,表明各雌株种质之间遗传变异大,遗传关系复杂。‘岭南’与‘黄金丸’之间的相似系数最大(0.83),说明二者遗传差异最小,亲缘关系最近。而‘西山大佛手’与‘叶籽银杏4’之间的相似系数最小(0.39),说明二者遗传差异最大,相似程度最小。
来自不同国家的雌株遗传变异程度不同。中国41个雌株之间的相似系数在0.39~0.80之间,平均相似系数为0.56,其变异系数高达14.29%,由此可见,国内41个雌株之间遗传变异较大。在这41个雌株中,‘垂叶银杏’与‘平展银杏’的相似系数较大(0.80),它们的遗传差异较小;而‘西山大佛手’与‘沂源叶籽银杏4’的相似系数较小(0.39),表明它们之间的遗传差异较大。在国外雌株中,日本‘叶籽银杏’、‘岭南’、‘黄金丸’、‘金兵卫’、‘藤九郎’的相似系数在0.61~0.83之间,平均相似系数为0.71,变异系数为8.45%,表明它们之间的遗传差异较小;其中,‘岭南’与‘黄金丸’之间的相似系数较大(0.83),说明它们亲缘关系最近;‘岭南’与‘藤九郎’之间的相似系数最小(0.61)。法国的‘展冠银杏’、‘特雷尼亚’、‘垂枝银杏’3个雌株中,‘特雷尼亚’与‘垂枝银杏’相似程度较大(0.77),相似程度较小的是‘展冠银杏’与‘垂枝银杏’(0.72),平均相似系数为0.75,变异系数却仅有3.21%,表明法国的3个雌株之间的遗传变异比较小,亲缘关系较近。
同一产地的雌株之间遗传变异也存在差异。泰安3个雌株的平均相似系数比较小(0.55),其变异系数为12.73%,其中,‘泰安2’与‘泰安3’的相似系数较小(0.47)。来自沂源的‘叶籽银杏1’、‘叶籽银杏2’、‘叶籽银杏3’、‘叶籽银杏4’、‘叶籽银杏5’、‘叶籽银杏6’、‘叶籽银杏7’7个雌株,平均相似系数较小(0.61),其变异系数为14.75%,表明这7个雌株之间的相似程度较大。对江苏的‘西山大佛手’、‘泰兴七星果’、‘大金坠’、‘泰兴家佛指’4个雌株进行分析,平均相似系数较大(0.64),其变异系数比较小(9.38%)。而来自浙江的‘天目山1’、‘天目山2’、‘天目山3’3个雌株,‘天目山2’与‘天目山3’之间的遗传差异较大(SC=0.58),3个雌株的平均相似系数比较大,为0.69,其变异系数为14.49%,表明这3个雌株之间的遗传变异相对较小。就叶籽银杏而言,它们也没有单独聚为一类,其遗传关系比较复杂。8个种质之间的相似系数在0.47~0.78之间,平均相似系数0.59,变异系数为16.02%,这表明叶籽银杏有较大的遗传变异;‘叶籽银杏1’与‘叶籽银杏4’相似性最小,而日本的‘叶籽银杏’和中国的‘叶籽银杏1’相似性最大,亲缘关系最近。
2.5 聚类分析在相似系数0.51处,供试材料分为2大类(图 1):Ⅰ类包括31个种质,即山东银杏种质‘叶籽银杏1’、‘崂山’、‘西湖’、‘五莲’以及法国、日本的雌株。Ⅱ类有18个种质,包括除了山东‘叶籽银杏1’、‘崂山’、‘西湖’、‘五莲1’之外的山东银杏雌株试材。这揭示出山东大部分雌株种质与其他的雌株种质有显著的遗传差异,亲缘关系较远。在相似系数为0.64时,可将银杏雌株分为7类:第1类包括法国、日本的雌株以及中国的‘叶籽银杏1’、‘垂叶银杏1’、‘平展银杏1’、‘盘县1’、‘盘县2’13个种质;第2类包括浙江的‘天目山1’、‘天目山2’、‘天目山3’、‘万源’、‘泰兴七星果’、‘泰兴家佛指’、‘宁国1’、‘灵川’、‘安陆1’、‘崂山’、‘西湖’11个种质;第3类包括‘东安1’、‘陕西周至’、‘腾冲1’、‘龙溪’、‘西山大佛手’、‘大金坠’和‘五莲’7个种质;第4类包括‘蒙阴’、‘济宁’、‘郯城1’、‘荣城’、‘莒县’、‘郯城2’、‘泗水’、‘叶籽银杏5’、‘泰安2’、‘叶籽银杏6’、‘叶籽银杏2’11个种质;第5类仅含‘泰安1’;第6类包括‘叶籽银杏3’、‘叶籽银杏4’、‘叶籽银杏7’、‘泰安3’;第7类仅含‘灵岩’;第8类仅有‘海阳’。就雌株而言,山东银杏雌株种质试材存在8种类型,占总类型的100%,可见,山东银杏雌株种质存在着较宽的遗传基础。地理距离较近的浙江种质(‘天目山1’、‘天目山2’、‘天目山3’)种质聚在一起,但是有些地理距离较远的种质(如‘灵川’、‘安陆1’、‘崂山’、‘日照西湖’等)也聚为一类,这说明种质的聚类与地理位置并不完全相关。另外,聚类结果也表明,尽管都是叶籽银杏,但是并没有聚为一类,可见,它们的遗传关系比较复杂。

王利等(2006b)研究了银杏观赏品种的特异种质。本研究分别对雌株的特异位点进行分析,13个雌株种质‘垂枝银杏’、‘平展银杏1’、‘盘县1’、‘东安1’、‘泰兴七星果’、‘灵川’、‘安陆’、‘崂山’、‘海阳’、‘荣城’、‘郯城2’、‘叶籽银杏2’、‘莒县’、‘泰安2’的单态带数较多(每个雌株4~12条),3个雌株种质‘西山大佛手’、‘海阳’、‘灵岩’的缺失带较多(每个雌株2~4条),分别占单态带和缺失带总数的69.42%、72.80%,它们比较特殊。
‘垂枝银杏’、‘平展银杏1’的相似性高达76%,且‘垂枝银杏’的多态性(60.29%)高于‘平展银杏’(58.38%);‘叶籽银杏2’与‘泰安2’同属于第4类,而且‘泰安2’的多态性、特异位点数分别比‘叶籽银杏2’大16.53%、5.79%。另外,‘崂山’与‘泰兴七星果’亲缘关系比较近(SC=0.77),且‘崂山’多态性、特异位点数分别比‘泰兴七星果’大2.14%、3.31%;‘岭南’与‘黄金丸’都为日本品种,相似程度高(SC=0.83),且又属于第1类,‘黄金丸’的多态性(63.26%)、特异位点数(3)分别高于‘岭南’的多态性(63.14%)、特异位点数(2)。可见,‘泰安2’、‘崂山’、‘垂枝银杏’更应优先得到保护。从聚类分析可以看出,‘泰安1’、‘灵岩’、‘海阳’分别聚为一类,表明它们比较特殊。
结合多态性、相似性、特异位点、聚类结果等对所试材料进行分析,法国的‘垂枝银杏’、日本的‘黄金丸’、中国的‘盘县1’、‘灵川’、‘安陆’、‘东安1’、‘西山大佛手’、‘荣城’、‘莒县’、‘郯城2’、‘泰安1’、‘泰安2’、‘泰安3’、‘崂山’、‘海阳’、‘灵岩’16个雌株分别属于不同的大类群与亚类群,具有一定的代表性,为极其重要的特异种质,应加以保护和利用。尽管这些种质数量仅占所试种质的32.65%,但其多态性却达到91.32%,占所试种质多态性的92.77%,这为银杏核心种质构建提供了依据。鉴于各个雌株种质具有不同的遗传特性,为了扩大银杏的遗传基础,丰富当地种质资源,在不同产地之间,应该有针对性地进行银杏的交流与引进。
3.2 银杏的遗传关系Tsumura等(1997)研究了98株银杏的遗传变异,中国与日本银杏亲缘关系近,认为日本银杏来自中国。华盛顿群体内12个个体间亲缘关系近,尼加拉河2个植株与华盛顿12个个体亲缘关系远,推测银杏物种水平的遗传多样性可能比较高(Kuddus et al., 2002)。21个银杏观赏品种的多态带比例为88%;各品种之间相似系数为0.45~0.85,表明它们存在不同程度的遗传差异和分化(王利等,2006b)。本研究49个雌株种质的多态带比例98.44%,高于21个银杏观赏品种的多态性(王利等,2006b),表明它们遗传差异大,遗传多样性丰富,这证明了Kuddus等(2002)的推测是正确的。研究还发现,大多数山东雌株种质聚在Ⅰ类中,表明它们亲缘关系近,这是由于山东是银杏的主要分布区之一,其银杏资源丰富,存在区域内相互引种;另外,该区受到暖温带季风气候的影响,使此区内的资源发生特定的趋同变异,致使区域内各种质之间相似性增加。至于山东银杏种质的起源与进化还有待于从细胞和分子水平上进行深入探讨。
‘叶籽银杏’也有称‘叶胚珠’银杏(phyllosporous或phyllospory),1927年Makino首次把叶籽银杏定为一个变种:Ginkgo biloba var. epiphylla Mak.,该变种以后被多人先后引用(李世美等,2007)。在日本叶籽银杏称为‘Ohatsuki phenomena’,‘Ohatsuki’这一名称被美国育种学家Santamour等引用,并作为一个重要的观赏品种在世界各地栽培(Santamour et al., 1983)。刘叔倩等(2001)构建了28个不同品种(或类型)银杏的RAPD和AFLP分子标记的遗传图谱,发现鸭尾银杏(Ginkgo biloba cv. yaweiyinxing)和叶籽银杏与其他26个样品之间的遗传差异相对较大。‘叶籽银杏1’比日本的‘叶籽银杏’更原始(邢世岩等,2007)。本研究对来自日本、中国的8株叶籽银杏的遗传关系进行AFLP分析表明,叶籽银杏并不是很原始的种质,‘海阳’位于聚类图的底部,且扩增的多态带较少(271条),与其他种质的相似系数0.44~0.61,平均相似系数0.50,该种质可能比较原始,这有助于进一步研究雌株种质的起源与进化。日本的‘叶籽银杏’与‘叶籽银杏1’相似性77%,该结果与以前研究的结果相一致(王利等,2006b),这与公元六世纪银杏从中国传至日本有关(邢世岩等,2004);而‘叶籽银杏1’与‘叶籽银杏4’亲缘关系最远,它们并没有聚为一类,其结果表明它们的起源不同。可见,尽管同为叶籽银杏,但是,它们之间亲缘关系比较复杂,其远近程度存在比较大的差异。生物性状的变异主要是通过基因突变、基因重组、基因互作以及染色体结构与数目的变异所产生的,另外,还受到环境条件的影响。已有研究表明叶籽银杏染色体数目基本上保持稳定2n=24(邢世岩等,2007),叶籽银杏的变异并不是由染色体数目的变异引起的,可能是由基因突变和环境条件共同作用的结果,至于其原因有待于进一步从细胞学、分子生物学等方面进行深入探讨。
陈月琴, 庄丽, 屈良鹄, 等. 1999. "活化石"植物银杏形态与分子进化(Ⅰ). 中山大学学报: 16-19. DOI:10.3321/j.issn:1672-3554.1999.01.004 |
葛永奇, 邱英雄, 丁炳扬, 等. 2003. 孑遗植物银杏群体遗传多样性的ISSR分析. 生物多样性, 11(4): 276-286. DOI:10.3321/j.issn:1005-0094.2003.04.002 |
李世美, 邢世岩, 李保进, 等. 2007. 叶籽银杏的发生及其个体与系统发育研究述评. 林业科学, 43(5): 90-98. |
刘叔倩, 马小军, 郑俊华. 2001. 银杏不同变异类型的RAPD指纹研究. 中国中药杂志, 26(12): 822-825. DOI:10.3321/j.issn:1001-5302.2001.12.009 |
沈永宝, 施季森, 赵洪亮. 2005. 利用ISSR DNA标记鉴定主要栽培银杏品种. 林业科学, 41(1): 202-204. DOI:10.3321/j.issn:1001-7488.2005.01.034 |
王利, 邢世岩, 韩克杰, 等. 2006a. 银杏雄株亲缘关系的AFLP分析. 中国农业科学, 39(9): 1940-1945. |
王利, 邢世岩, 杨克强, 等. 2006b. 银杏观赏品种遗传关系的AFLP分析. 遗传学报, 33(11): 1020-1026. |
王晓梅, 宋文芹, 刘松, 等. 2001a. 利用AFLP技术筛选与银杏性别相关的分子标记. 南开大学学报:自然科学版, 34(1): 5-9. |
王晓梅, 宋文芹, 刘松, 等. 2001b. 与银杏性别相关的RAPD标记. 南开大学学报:自然科学版, 34(3): 116-117. |
吴俊元, 陈品良, 汤诗杰. 1992. 天目山银杏群体遗传变异的同工酶分析. 植物资源与环境, 1(2): 20-23. |
邢世岩, 高进红, 姜岳忠, 等. 2007. 银杏特异种质核型进化趋势. 林业科学, 43(1): 21-28. |
邢世岩, 郭彦彦, 王利, 等. 2004. 银杏种质遗传多样性研究述评. 经济林研究, 22(4): 67-70. |
Cervera M T, Stome V, Soto A. 2005. Intraspecific and interspecific genetic and phylogenetic relationships in the genus Populus based on AFLP markers. Theoretical and Applied Genetics, 111: 1440-1456. DOI:10.1007/s00122-005-0076-2 |
Doyle J J, Doyle J L. 1990. Isolation of plant DNA from fresh tissue. Focus, 12: 13-15. |
Ferdinandez Y S N, Coulman B E. 2004. Genentic relationships among smooth bromegrass cultivars of different ecotypes detected by AFLP mar kers. Crop Science, 44(1): 241-247. DOI:10.2135/cropsci2004.2410 |
Kuddus R H, Kuddus N N, Dvorchic I. 2002. DNA polymorphism in the living fossil Ginkgo biloba from the Eastern United States. Genome, 45: 8-12. DOI:10.1139/g01-115 |
Santamour F, He S A, McArdle A J. 1983. Checklist of cultivated Ginkgo. J Arboriculture, 9(3): 88-92. |
Shen Lang, Chen Xiaoyong, Zhang Xin, et al. 2005. Genetic variation of Ginkgo biloba L (Ginkgoaceae) based on cpDNA PCR-RFLPs: inference of glacial refugia. Heredity, 94(4): 396-401. DOI:10.1038/sj.hdy.6800616 |
Tsumura Y, Ohaba K. 1997. The genetic diversity of isozymes and the possible dissemination of Ginkgo biloba in ancient times in Japan//Hori T, Ridge R W, Tuleeke W, et al. Ginkgo biloba—a global treasure. Spinger-Verlag, Tokyo, 159-172. https://link.springer.com/chapter/10.1007%2F978-4-431-68416-9_13
|