 2008, Vol. 44
2008, Vol. 44文章信息
- 李亚青, 张钢, 卻书鹏, 祝良, 邸葆, 靳秀梅.
- Li Yaqing, Zhang Gang, Que Shupeng, Zhu Liang, Di Bao, Jin Xiumei.
- 白皮松茎和针叶的电阻抗参数与抗寒性的相关性
- Relation between Electrical Impedance Spectroscopy Parameters and Frost Hardiness in Stems and Needles of Pinus bungeana
- 林业科学, 2008, 44(4): 28-34.
- Scientia Silvae Sinicae, 2008, 44(4): 28-34.
-
文章历史
- 收稿日期:2007-05-18
-
作者相关文章



2. 北京十三陵林场苗圃 北京 102200
2. The Nursery in Beijing Thirteen Tombs Forestry Center Beijing 102200
抗寒锻炼期间,植物组织的细胞发生生理、生化变化。在这个过程中,细胞膜、细胞质和液泡发生的变化比较突出。许多测定植物抗寒性的方法都比较耗时、费力,并且会破坏植物组织,在生产应用上有一定的局限性。因此开发一种测定植物抗寒性的简捷快速的方法很有必要(Repo et al., 1997)。测定植物组织和器官的电阻抗图谱法(electrical impedance spectroscopy, EIS)就是这样一种方法。EIS法用等效电路(equivalent electrical circuit)来表示所测定的组织样本,已被广泛地用于估测植物活力(MacDougall et al., 1987)、果实受害程度(Cox et al., 1993)、抗寒性(Repo et al., 1993; 1997; 2000a; Stout, 1988a; 1988b)、对含盐量敏感度(Mancuso et al., 1996)、根系生长(Ozier-Lafontaine et al., 2005; Repo et al., 2005)等研究中。在所有这些研究中,EIS法提供了非破坏性的测定胞内电阻、胞外电阻和膜变化的手段(Mancuso, 1998)。EIS法采用实部和虚部组成的图谱对电阻抗数据分析来测定植物的抗寒性(Repo et al., 1990)。在对欧洲赤松(Pinus sylvestris)幼树当年生枝条进行的试验中发现,用电导法求出的针叶抗寒性与EIS法用胞外电阻率re求得的茎抗寒性之间存在着很大的相关性(Repo et al., 1997)。因此,在抗寒锻炼期可以通过EIS法得到白皮松(Pinus bungeana)茎和针叶细胞的电学物质变化。在我国,EIS法测定植物抗寒性及其在植物生理学领域的研究尚属全新领域。通过EIS法和常规的电导法测定抗寒锻炼期间不同种源白皮松茎和针叶的抗寒性,并比较2种方法的相关性,以完善EIS法测定植物抗寒性技术。认为,随着抗寒性的提高,植物组织发生生理生化变化,其电阻抗参数也会随之发生变化,因此茎和针叶的抗寒性可以用EIS法测定。
1 材料与方法 1.1 植物材料白皮松1年生枝取自北京市十三陵林场昊林苗圃的白皮松种质资源保存兼种源试验林的8年生实生苗。包括北京蟒山(40°44′ N,116°35′ E)、甘肃两当(33°55′ N,106°40′ E)、山西孝义(37°05′ N,111°45′ E)3个种源。每个种源选择10株树从2006年9月至2007年1月对茎和针叶进行电阻抗和电导的测定。每月采样1次,每株树上随机选取6根1年生枝段,每种源60枝段。
1.2 不经冷冻处理的茎和针叶的电阻抗每次测定时,从每个种源的60枝段中随机选取16枚针叶和16段茎进行电阻抗测定。材料保存于塑料袋里,储存于4 ℃冰箱。每枚针叶从中部切取15 mm,枝段去除针叶后从中部切取15 mm用于测定。采用Ag/AgCl电极,在电极上加电极凝胶,使针叶或茎样本的横切面与凝胶接触。电阻抗仪(HP4284A, USA)设定从80 Hz至1 MHz共42个频率。茎阻抗分析图谱用双-DCE模型(Repo et al., 2000a)表示:
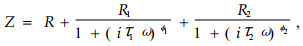
|
(1) |
式中:Z为总阻抗,ω为角速度(ω=2×π×f,其中f为AC频率),i为虚部单位。双-DCE模型共有7个需测定的参数,即3个电阻(R、R1和R2)、2个弛豫时间(τ1和τ2)、2个在不同弛豫时间的分布系数(distribution coefficient, ψ1和ψ2)。图 1为欧洲赤松茎的电阻抗图谱的示意图,展现了双-DCE模型各参数的数学解释(Repo et al., 2000a)。R1和R2分别根据高频弧和低频弧与X轴的交叉点获得,τ1和τ2由阻抗图谱高频弧或低频弧最高值求得,位于X轴下方的圆心分别定义为ψ1和ψ2。等效电路模型的参数值可用复合非线性最小二乘方(complex non-linear least squares, CNLS)下的曲线进行估算(Macdonald, 1987)。

|
图 1 欧洲赤松茎的电阻抗图谱示意图(引自Repo et al., 2000a) Figure 1 A schematic drawing of an impedance spectrum of stem of Pinus sylvestris (from Repo et al., 2000a) 分布模型(Double-DCE)参数的数学解释详见Repo et al., 2000a图 1和附录1,图谱从80 Hz到1 MHz共42个频率(从右至左)。 The mathematical interpretation of the distributed model parameters (double-DCE) see Repo et al., 2000a Fig. 1 and Appendix 1. The spectrum is composed of 42 different frequencies ranging from 80 Hz to 1 MHz from right to left, respectively. |
低频电流不能通过细胞膜,但可在胞外间隙流动,所以胞外电阻Re(extracellular resistance)可用下式求得:
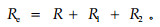
|
(2) |
而高频电流可以通过细胞膜,并相应地既可在胞外间隙也可在胞内流动,因此胞内电阻Ri(intracellular resistance)可用下式求得:
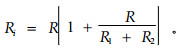
|
(3) |
由于样本的横切面面积及长度会成比例地分别降低和提高阻抗,需要考虑组织的几何形状。也就是说,要对测定的电阻参数求出其电阻率rx:
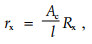
|
(4) |
式中:rx是电阻率(x表示胞外e或胞内i),单位为Ωm;Rx是测定的电阻(x表示胞外e或胞内i),单位为Ω;Ac是样本横切面面积,单位为m2(对于白皮松茎,则Ac=π×d2/4,d为茎的直径);l是样本长度,单位为m (l=0.015 m) (Repo et al., 1990; 2000a)。
针叶叶肉细胞用模型A (Zhang et al., 1995; 2002)表示,总阻抗(ZModel-A)是:
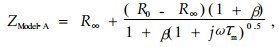
|
(5) |
式中:R∞和R0分别是在极高频与极低频下的电阻;τm是细胞膜的时间恒量,τm=R3×Cm, 其中R3和Cm分别为细胞膜的电阻及电容;系数β是控制图谱偏度及凹陷中心的一个因子(Zhang et al., 1995);j=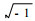
低频电阻R0对应于胞外电阻Re。而胞内电阻Ri由下式求得:
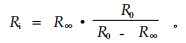
|
(6) |
针叶的电阻率rx也由公式(4)求出。白皮松针叶的横切面被看作是圆心角为60°的扇形,故A针叶=π×d2/3,式中d表示针叶的厚度,样本长度l为15 mm。
双-DCE模型和模型A利用LEVM v8.06软件拟合数据,得出模型参数(电阻抗参数的缩写词见表 1)。
|
|

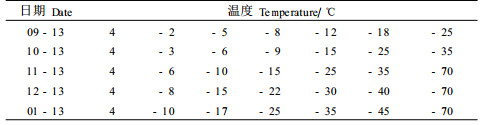
样本用清水清洗3次,去除样本表面污染物,再用去离子水洗3次。洗后用吸水纸吸干水分,分别装于塑料袋中,每袋放6个枝段,每种源需塑料袋7个。袋中喷少量去离子水,以免材料发生过冷。冷冻处理设7个温度(表 2),以4 ℃为对照,置于冰箱内,其他6个温度包括使样本全部成活和全部死亡的温度。降温速率6 ℃·h-1,到达每个温度时保持4 h,每种源取出1袋,置于4 ℃冰箱中缓慢解冻24 h。
|
|
冷冻处理的茎和针叶缓慢解冻后,每种源每温度设8个重复,进行电阻抗的测定,方法同1.2。
用电导法(electrolyte leakage, EL)测定抗寒性(张钢, 2005; Repo et al., 2000a; Zhang et al., 2003):缓慢解冻后,每枚针叶从中部切取10 mm,用去离子水洗净后放入试管中,每个试管放8段针叶。每段枝从中部切取10 mm,再纵切成4段,用去离子水洗净后放入试管中,每个试管放4段茎段。每个温度设4个重复。每试管加13 mL去离子水,用胶塞封口,放入摇床中振荡24 h。用DDSL-308型电导仪(上海京科雷磁)测渗出液的初电导值(C1)。胶塞封口后将试管置于沸水中煮沸20 min,再放入摇床中振荡24 h,测终电导值(C2)。用公式(7)计算相对电导率(E):
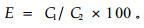
|
(7) |
相对电导率的变化或电阻抗参数re与抗寒性的关系可用Logistic方程(Repo et al., 1989)表示:
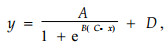
|
(8) |
式中: x(冷冻处理温度)、y(相对电导率或re)由A(处理相对电导率的最高值与最低值之差或最高re与最低re的差)、B(温度拐点处曲线的斜率)、C(拐点温度)和D(处理相对电导率最低值或re最低值)4个参数决定。应用SPSS 13.0软件通过计算方程的拐点温度(C值)表示组织半致死温度(LT50),估算抗寒性。
2 结果与分析 2.1 未经冷冻处理的茎和针叶的电阻抗白皮松未冷冻茎和针叶的电阻抗图谱的形状和电阻抗参数在抗寒锻炼期间都在发生变化。如图 2A所示,茎的电阻抗图谱在9月呈现2个半径相差不多的重叠的弧,从10月开始,两弧半径相差增加,近似于1个弧。从图 2B可以看出,叶的电阻抗图谱始终只有1个弧,在低频时数值不稳定。茎与针叶的图谱在高频时,不同日期测定的电阻(实部)与电抗(虚部)相等。

|
图 2 不同时期未冷冻白皮松茎(A)与针叶(B)的电阻抗图谱 Figure 2 Impedance spectra of non-frost-exposed stems and needles 图谱由3个种源的数据(从80 Hz到1 MHz共42个频率,从右至左)平均而得。 The spectra are the pooled data of three provenances and composed of 42 different frequencies ranging from 80 Hz to 1 MHz (from right to left, respectively). |
茎的胞外电阻率re从9月到10月呈下降趋势,从10月到1月呈缓慢上升趋势(图 3A)。叶的胞外电阻率re从9月到10月呈下降趋势,从10月到12月呈上升趋势,从12月到1月基本保持不变(图 3B)。茎的胞内电阻率ri从9月到11月呈上升趋势,从11月开始基本保持不变(图 3C)。叶的胞内电阻率ri抗寒锻炼期间呈上升趋势(图 3D)。叶的细胞膜时间恒量τm与叶的胞外电阻率re的变化趋势相同,呈先减小再增加后减小的变化趋势(图 3E)。叶的β系数在抗寒锻炼初期各种源差异显著,从9月到11月逐渐减小,11月至1月各种源无差异,且基本保持不变(图 3F)。

|
图 3 抗寒锻炼期间各种源未冷冻茎与针叶电阻抗参数的变化 Figure 3 The variation of impedance parameters of non-frost-exposed stems and needles during frost hardening |
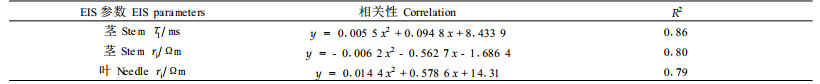
在对白皮松茎和针叶抗寒性和未冷冻电阻抗参数的关系研究中,发现茎的弛豫时间τ1、茎的胞内电阻率ri和叶的胞内电阻率ri与经冷冻处理用电导法求得的抗寒性直接相关(表 3)。
|
|
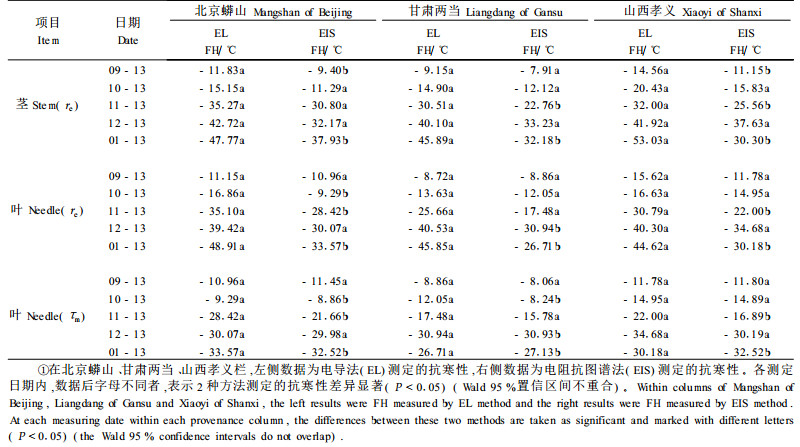
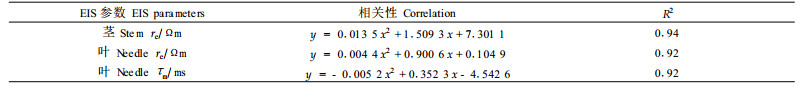
样本经过冷冻处理后,茎和针叶的胞外电阻率re、针叶的细胞膜时间恒量τm随着冻害的增强而下降。因此样本经过冷冻处理后可通过re和τm确定抗寒性。表 4为通过EIS法测定茎和针叶的re与针叶的τm求得的抗寒性和EL法求得的茎和针叶的抗寒性的比较。可以看出,在抗寒性较低时,用EIS法测出的抗寒性与用EL法测出的抗寒性相差不多,而在抗寒性较强时,用EIS法测出的抗寒性比用EL法测出的抗寒性低。EIS法与EL法测定白皮松茎和针叶的抗寒性均呈线性相关,R2=0.91(EL法与re测定白皮松茎和针叶的抗寒性)和R2=0.93(EL法与τm测定白皮松针叶的抗寒性)。此外,用EIS法得出的一些电阻抗参数与用EL法测出的抗寒性显著相关(表 5)。
|
|
|
|
茎的电阻抗图谱在9月呈现2个半径相差不多的重叠的弧,从10月开始,两弧的半径大小差异增加,近似于1个弧。这与Repo等(2000b)对欧洲赤松茎的研究结果有所区别,在抗寒锻炼期间欧洲赤松茎的电阻抗图谱始终为双弧;但与Zhang等(2002)关于欧洲赤松针叶的图谱变化趋势类似,即在抗寒锻炼期间始终为单弧。不过,本研究中无论茎或针叶,电阻(实部)和电抗(虚部)在高频时相等(图 2),与Repo等(2000b)和Zhang等(2002)的结果不同。这些不同之处也许是树种不同造成的。
9月初至10月初,即在抗寒锻炼准备阶段和抗寒锻炼初始阶段,茎的re、叶的re和τm都在减小,可以假设一旦到达某最低点,即临界点,白皮松已经具备了抗寒锻炼的条件,准备开始过冬。在抗寒性大幅度提高时,即10月初至12月初,茎的re、叶的re和τm也随之提高,可能是由于材料抗寒性逐渐提高,细胞合成某种蛋白以帮助植物抵抗低温。
在抗寒锻炼期,茎的弛豫时间τ1、茎的胞内电阻率ri和叶的胞内电阻率ri逐渐增加,与抗寒性变化趋势相同,并与抗寒性具有相关性。在抗寒锻炼期,细胞内可溶性糖含量增加,糖不仅与提高抗寒性有关,而且是低流动性电解质,糖含量的增加提高了细胞液浓度,这就导致了细胞质电阻的提高,这或许能够解释EIS参数在抗寒锻炼期的变化趋势。早期对欧洲赤松(Repo et al., 2000a; Zhang et al., 2003)、紫花苜蓿(Medicago stativa)、鸟爪三叶草(Lotus corniculatus) (Stout, 1988a; 1988b)和蒿柳(Salix viminalis) (Repo et al., 1997)的研究也发现,胞内电阻率ri随抗寒锻炼进程而提高。茎的弛豫时间τ1的变化也许与其细胞木质化及细胞种类有关(Repo et al., 1997)。表明,在样本不经冷冻处理情况下,参数τ1、ri可以用来预测抗寒性。
3.2 冷冻处理的电阻抗参数与抗寒性的相关性对白皮松茎和针叶电阻抗参数与抗寒性的关系研究中,发现胞外电阻率re、细胞膜时间恒量τm与抗寒性直接相关。根据EIS参数的变化趋势,样本经过冷冻处理后,胞外电阻率re是确定茎抗寒性最合适的一个参数。由于原生质膜受冻害后胞内离子外渗到细胞间隙,使胞外电阻下降,所以胞外电阻随冻害增强而下降(Repo et al., 1994; 1997; 2000a; 2000b)。Mancuso等(2004)用EIS法和EL法测定4种红千层属(Callistemon)和2种银桦属(Grevillea )植物的抗寒性的结果非常类似。但在样本抗寒性较强时,EIS法会比EL法低估抗寒性。这与Repo等(1994)的试验结果是相同的。可能是由于样本抗寒性增强后,EL法的适应性较差,离子渗出率也会随季节变化(张钢, 2005)。本文的研究发现,细胞膜时间恒量τm也可用来估测针叶抗寒性,相比胞外电阻率re,τm在用于测定针叶抗寒性时与EL法较相近。
4 结论1) 茎和针叶的EIS随着抗寒性的增强发生变化;2)样本不经冷冻处理,可通过茎的弛豫时间τ1、胞内电阻率ri和针叶的胞内电阻率ri测定抗寒性;3)样本经冷冻处理后,可通过茎与针叶的胞外电阻率re、叶的细胞膜时间恒量τm测定抗寒性。在实际生产中,需要开发一种快速、简捷的测定植物抗寒性的方法,EIS法是一种可行的方法。然而,在国内对这种方法的研究还处于起步阶段,因此,还要对不同树种、不同种源作进一步的研究,为更好地理解和广泛地应用EIS法测定植物抗寒性提供理论依据与技术手段。
张钢. 2005. 国外木本植物抗寒性测定方法综述. 世界林业研究, 18(5): 14-20. DOI:10.3969/j.issn.1001-4241.2005.05.004 |
Cox M A, Zhang Min, Willison J H M. 1993. Apple bruise assessment through electrical impedance measurements. Journal Horticultural Science, 68: 393-398. DOI:10.1080/00221589.1993.11516366 |
Macdonald J R. 1987. Impedance Spectroscopy: emphasizing solid materials and systems. New York: John Wiley & Sons, Inc..
|
MacDougall R C, Thompson R G, Piene H. 1987. Stem electrical capacitance and resistance measurements as related to total foliar biomass of balsam fir trees. Canadian Journal of Forest Research, 17: 1071-1074. DOI:10.1139/x87-164 |
Mancuso S, Rinaldelli E. 1996. Response of young mycorrhizal and non-mycorrhizal plants of olive tree (Olea europaea L.) to saline conditions Ⅱ. Dynamics of electrical impedance parameters of shoots and leaves. Advances in Horticultural Science, 10: 135-145. |
Mancuso S. 1998. Seasonal dynamics of electrical impedance parameters in shoots and leaves relate to rooting ability of olive (Olea europaea) cuttings. Tree Physiology, 19: 95-101. |
Mancuso S, Nicese F P, Masi E, et al. 2004. Comparing fractal analysis, electrical impedance and electrolyte leakage for the assessment of cold tolerance in Callistemon and Grevillea spp. Journal of Horticultural Science & Biotechnology, 79: 627-632. |
Ozier-Lafontaine H, Bajazet T. 2005. Analysis of root growth by impedance spectroscopy (EIS). Plant Soil, 277: 299-313. DOI:10.1007/s11104-005-7531-3 |
Repo T, Lappi J. 1989. Estimation of standard error of impedance-estimated frost resistance. Scandinavian Journal of Forest Research, 4: 67-74. DOI:10.1080/02827588909382547 |
Repo T, Pelkonen P, Tuovinen T. 1990. Frost hardiness assessment: New developments//Proceedings of the XIX IUFRO World Congress, division 2, 101-106.
|
Repo T, Zhang Min. 1993. Modelling woody plant tissues using a distributed electrical circuit. Journal of Experimental Botany, 44: 977-982. DOI:10.1093/jxb/44.5.977 |
Repo T, Zhang Min, Ryypp A, et al. 1994. Effects of freeze-thaw injury on parameters of distributed electrical circuits of stems and needles of Scots pine seedlings at different stages of acclimation. Journal of Experimental Botany, 45: 823-833. DOI:10.1093/jxb/45.6.823 |
Repo T, Leinonen M, Paakkonen T. 1997. Electrical impedance analysis of shoots of Scots pine: intracellular resistance correlates with frost hardiness//Proceedings of the Finnish-Japanese Workshop on Molecular and Physiological Aspects of Cold and Chilling Tolerance of Northern Crops. Jokioinen, Finland, 27-30.
|
Repo T, Zhang Gang, Ryypp A, et al. 2000a. The electrical impedance spectroscopy of Scots pine (Pinus sylvestris L.) shoots in relation to cold acclimation. Journal of Experimental Botany, 51: 2095-2107. DOI:10.1093/jexbot/51.353.2095 |
Repo T, Zhang Gang, Ryypp A, et al. 2000b. The relation between growth cessation and frost hardening in Scots pines of different origins. Trees, 14: 456-464. DOI:10.1007/s004680000059 |
Repo T, Laukkanen J, Silvennoinen R. 2005. Measurement of the tree root growth using electrical impedance spectroscopy. Silva Fennica, 39(2): 159-166. |
Stout D G. 1988a. Effect of cold acclimation on bulk tissue electrical impedance Ⅰ. Measurements with birdsfoot trefoil at subfreezing temperatures. Plant Physiology, 86: 275-282. DOI:10.1104/pp.86.1.275 |
Stout D G. 1988b. Effect of cold acclimation on bulk tissue electrical impedance Ⅱ. Measurements with alfalfa and birdsfoot trefoil at nonfreezing temperatures. Plant Physiology, 86: 283-287. DOI:10.1104/pp.86.1.283 |
Zhang Gang, Ryypp A, Repo T. 2002. The electrical impedance spectroscopy of Scots pine needles during cold acclimation. Physiologia Plantarum, 115: 385-392. DOI:10.1034/j.1399-3054.2002.1150308.x |
Zhang Gang, Ryypp A, Vapaavuori E, et al. 2003. Quantification of additive response and stationarity of frost hardiness by photoperiod and temperature in Scots pine. Canadian Journal of Forest Research, 33: 1772-1784. DOI:10.1139/x03-100 |
Zhang Min, Repo T, Willison J H M, et al. 1995. Electrical impedance analysis in plant tissues: on the biological meaning of Cole-Cole α in Scots pine needles. European Biophysics Journal, 24: 99-106. |