 2008, Vol. 44
2008, Vol. 44文章信息
- 王祥福, 郭泉水, 巴哈尔古丽, 刘正宇, 任明波.
- Wang Xiangfu, Guo Quanshui, Bahar Guli, Liu Zhengyu, Ren Mingbo.
- 崖柏群落优势乔木种群生态位
- Niche of Dominant Arbor Populations in Thuja sutchuenensis Community
- 林业科学, 2008, 44(4): 6-13.
- Scientia Silvae Sinicae, 2008, 44(4): 6-13.
-
文章历史
- 收稿日期:2007-03-12
-
作者相关文章


2. 中国林业科学研究院新疆分院 乌鲁木齐 830000;
3. 重庆市药物种植研究所 重庆 408435
2. Xinjiang Branch of Chinese Academy of Forestry Urumqi 830000;
3. Institute of Chongqing Medical Plant Cultivation Chongqing 408435
崖柏(Thuja sutchuenensis)为柏科(Cupressaceae)崖柏属(Thuja)常绿乔木。它的木材化石出现于侏罗纪中期,在白垩纪曾经出现鼎盛时期,到第三纪该属物种大量消失。1892年4月,法国传教士法戈斯(R.P.Farges)在我国重庆市城口县石灰岩山地首次采获标本,编号为Farges 1158。7年后,该标本作为新种的模式标本,收藏于法国巴黎自然博物馆。此后100余年间,曾有人多次前往产地调查,但均未见其踪迹,为此很多专著都将其定为野外灭绝物种(Fu et al.,1992;1999)。1999年,刘正宇先生在原标本采集地重新发现了崖柏,并采集到了带球果的标本。现在该标本收藏于中国科学院植物研究所标本馆内,并先后得到中国科学院、英国皇家植物园和美国哈佛大学权威专家的确认。2003年IUCN将其评定为极危物种。
生态位是种群生态研究的核心问题(Grinell,1917;柳江等,2002),就其本质而言,生态位是物种在特定尺度下特定环境中的功能单位,包括物种对环境的要求和环境对物种的影响两方面及其相互作用规律,是物种属性的特征表现(Aplet et al.,1994;Hurlbert,1978;Leibold et al.,1995)。在众多的生态学理论中,生态位理论一直处于十分醒目的地位,并一度成为生态学研究的热点和焦点(李德志等,2006a)。通过生态位的研究可以探讨种间关系、群落结构、群落演替、生物多样性和物种进化等方面的科学问题(任青山,1998)。近年来,濒危植物的种群生态位研究倍受重视,并已取得了一些成果(毕润成等,2003;刘金福等,1999;王琳等,2001;张峰等,2004),但关于崖柏种群生态位的研究尚未见报道。目前对崖柏的研究主要集中在分类学描述(Xiang et al.,2002)、木材结构(唐熙等,2005)、种群大小级结构及种群动态(刘建锋等,2004)、种群分布格局(刘建锋等,2005)、球果特征及出种量(王祥福等,2007)、优势乔木树种种间关系(郭泉水等,2007)等几个方面。本研究旨在通过崖柏群落优势乔木种群生态位分析,揭示崖柏和崖柏群落中乔木优势种群对可利用资源的分享状况以及种间关系,为进一步保护和扩繁这一极度濒危物种提供科学依据。
1 研究地概况研究在大巴山山系的重庆市城口县和开县山区进行(108°30′—109°15′ E,31°30′—31°50′ N)。研究区海拔900~2 200 m,年均气温为6.0~10.0 ℃,1月平均气温-0.8~4.5 ℃,7月平均气温15.2~20.2 ℃,极端最低气温-10.0 ℃,极端最高气温30.0 ℃,全年无霜期150~200 d,年日照时数1 000~1 200 h,年降水量1 200~1 400 mm,≥0 ℃年积温2 580~3 880 ℃,≥10 ℃年积温1 290~2 970 ℃,积雪期3个月左右。地带性植被类型为中亚热带常绿阔叶林(吴征镒,1983)。崖柏集中分布地段的土壤类型为石灰岩母质上发育的山地褐土或棕褐土。
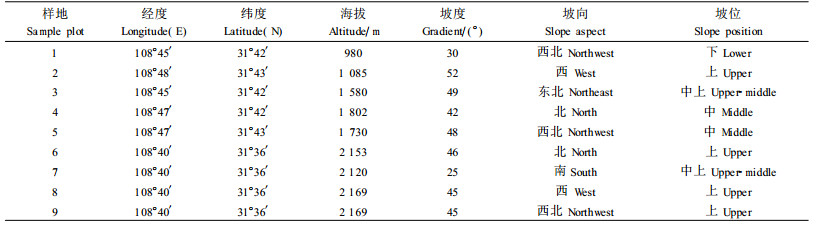
2 研究方法 2.1 样地设置和调查崖柏天然分布的海拔下限为980 m,上限为2 169 m,沿着海拔梯度选择典型地段,设置9块20 m×20 m的调查样地(表 1)。在每块样地内均匀设置4个10 m×10 m的乔木样方、4个5 m×5 m的灌木样方和4个2 m×2 m的草本样方。在乔木样方内,调查乔木的种类、株数、胸径、树高及幼苗幼树的高度和基径;在灌木和草本样方内,调查灌木和草本植物的种类、多度和盖度。
|
|
重要值=相对密度+相对频度+相对优势度,相对显著度以该树种的胸高断面积之和与所有树种总胸高断面积的比值表示(宋永昌,2001)。
2.3 优势乔木种群的生态位宽度将每个样方作为一维资源状态,以每个样方中不同乔木树种的个体数作为计算水平生态位宽度和垂直生态位宽度(臧润国等,2005)的表征特征。水平生态位宽度的测度采用Shannon-Wiener多样性指标(侯琳等,2006)和Levins生态位宽度指数(吴刚等,1999),计算公式如下:
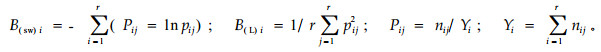
|
式中:B(sw)i为树种i采用Shannon-Wiener多样性指标测定的生态位宽度值;B(L)i为树种i采用Levins生态位宽度指数测定的生态位宽度;Pij为树种i利用第j种资源占其所利用全部资源的比例;r为资源位数(样方数),nij为树种i利用第j种资源位的个体数(用乔木树种的个体数表示),Yi为树种i所利用全部资源位的个体总数。B(sw)i标准化形式为B(sw)i′=1/lnr×B(sw)i,B(L)i和B(sw)i′具有域值[0,1]。
以1 m为1高度级(资源位),统计各高度级内出现的每个乔木树种的个体数,以此为基础,计算不同乔木树种的垂直生态位宽度。垂直生态位宽度与水平生态位宽度计算方法一样,但计算公式中各符号的含义不完全相同。其中资源位数(r)为高度级数;Pij为某乔木树种i在第j资源位(高度级)中的个体数比例。
2.4 优势乔木种群的生态位重叠计算内容包括水平生态位重叠和垂直生态位重叠。应用以下公式计算
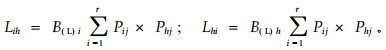
|
式中:Lih和Lhi分别是i种对h种和h种对i种的生态位重叠,Lih和Lhi的值域为[0,1];Pij和Phj分别是种i和种h对第j个资源的利用率,用植物种的个体数表示;B(L)i和B(L)h分别是种i和种h的Levins生态位宽度(黄英姿,1994)。
3 结果与分析 3.1 崖柏群落植物组成按照各样地所处的海拔,将样地分别归并为海拔1 085 m以下(1号和2号样地)、海拔1 085~2 000 m(3、4、5号样地)及海拔2 000 m以上(6、7、8、9号样地)3个地段,同时将样地调查资料分地段进行统计。结果表明:在海拔1 085 m以下的崖柏群落乔木层中,主要树种包括崖柏、大叶青冈(Cyclobalanopsis jenseniana)、小叶青冈(C. gracilis)和川陕鹅耳枥(Carpinus fargesiana),林分郁闭度为0.30~0.85,平均树高为5.57~7.75 m,平均胸径为7.82~8.14 cm;在海拔1 085~2 000 m的崖柏群落乔木层中,主要树种包括崖柏、大叶青冈和华千金榆(Carpinus cordata),林分郁闭度为0.6~0.9,平均树高为4.68~5.97 m,平均胸径为7.82~8.59 cm;在海拔2 000 m以上的崖柏群落乔木层中,主要树种包括崖柏、高山栎(Quercus semicarpifolia)、川陕鹅耳枥等,林分郁闭度为0.40~0.55,平均树高为4.59~6.76 m,平均胸径为9.05~11.69 cm。
在海拔1 085 m以下的崖柏群落灌木层和草本层中,主要灌木包括短枝六道木(Abelia engleriana)、月月青(Itea ilicifolia)、细枝柃(Eurya loquaiana)及含羞草叶黄檀(Dalbergia mimosoides),主要草本植物包括丝叶苔草(Carex capilliformis);在海拔1 085~2 000 m处,主要灌木包括粉红杜鹃(Rhododendron oreodoxa)、短枝六道木、具柄冬青(Ilex pedunculosa)、金叶柃(Eurya aurea)、蠔猪刺(Berberis julianae)、城口黄栌(Cotinus coggygria)及小叶栒子(Cotoneaster microphylla),主要草本植物为丝叶苔草(Carex capilliformis)及蔓龙胆(Crawfurdia japonica);在海拔2 000 m以上,主要灌木包括粉红杜鹃、中华吊钟花(Enkianthus chinensis)、红柄木犀(Osmanthus armatus)、细叶黄杨(Buxus bodinieri)、蠔猪刺及木姜子(Litsea pungens),主要草本植物包括杉蔓石松(Lycopodium annotinum)、红毛尾药菊(Synotis erythropappa)及扁竹根(Iris japonica)。
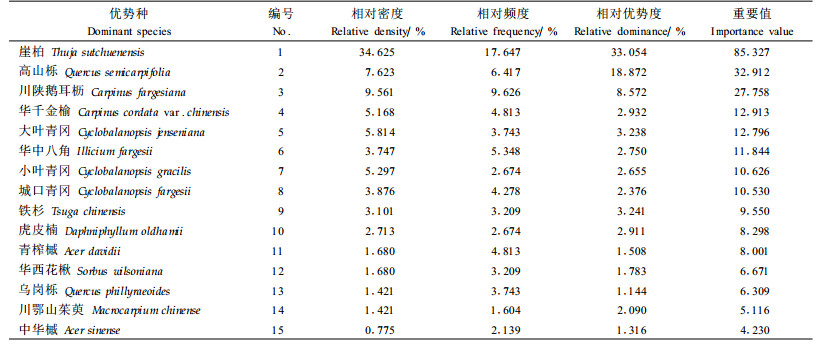
3.2 主要乔木树种重要值在本次调查中,崖柏群落的乔木层中共出现41个树种,其中前15位树种的重要值之和占84%。可见,这15种乔木树种在崖柏群落中具有重要的建群作用,为崖柏群落的优势树种(表 2)。
|
|
采用Shannon-Wiener多样性指标和Levins生态位宽度指数2种方法计算优势乔木树种的水平生态位宽度值,目的在于通过2种计算方法检验生态位宽度值的变化趋势是否一致, 而垂直生态位宽度仅采用Levins生态位宽度指数计算。结果见表 3。
|
|
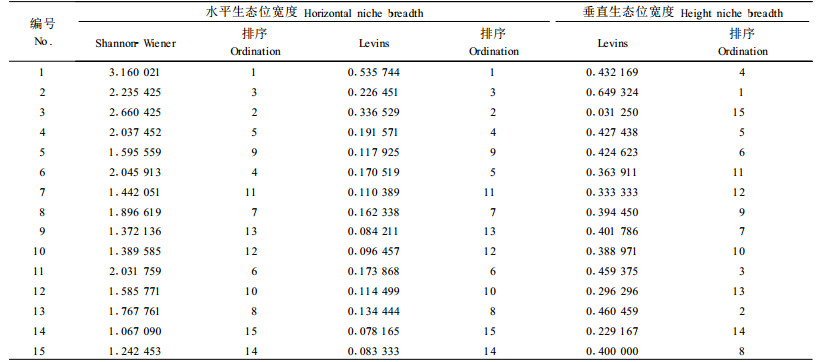
由表 3可知,不同乔木种群的生态位宽度存在一定的差异,2种计算方法得到的结果基本一致。按照水平生态位宽度的大小排列,处在前3位的是崖柏、高山栎及川陕鹅耳枥;排在后2位的是川鄂山茱萸及中华槭;其他树种如华千金榆、大叶青冈、华中八角、小叶青冈、城口青冈、铁杉、虎皮楠、青榨槭、华西花楸、乌岗栎的生态位宽度居于中间位置。
生态位宽度是度量植物种群对资源环境利用状况的尺度(李生等,2003;Weider,1993;张继义等,2003)。水平生态位宽度反映不同树种在水平空间上对资源的利用和占据能力。影响生态位宽度的因素很多,其中树种的生态习性是其中的一个重要原因。实地调查发现,崖柏群落具有柏类林共有的生态习性(蒋有绪等,1998),主要特征是抗风、耐干旱、耐土壤贫瘠或生境贫瘠,无论处在何等的热量条件(如不同的纬度,不同的海拔等)下,它都喜生于阳光充足、通风、干燥的生境中。崖柏自然分布地段多为向阳的陡坡、山脊或石质山坡。与崖柏相比,生态位宽度较大的高山栎也基本具备这些特性。高山栎天然分布区气候干湿季明显,冬季干旱、雨量少、日照强、日温差大。在气候相对温凉、土壤肥沃深厚的地段高山栎长势良好,在干旱贫瘠的地段也能生存。由于树种之间竞争的作用,天然分布区以阳坡、山脊、陡坡、岩石露头多、相对干旱贫瘠的地方为主。栎属、青冈属、栲属及水青冈属中的多数树种,都是我国常绿阔叶林和常绿落叶阔叶混交林树种组成中的基本成分,是建群种或优势种,但崖柏种群能够生存的环境,对它们而言可能不是最为适宜的,从而导致生态位宽度变窄。
植被的垂直分布也是影响崖柏群落优势乔木种群生态位宽度的重要因素。已有研究表明,川陕鹅耳枥天然分布的海拔范围为1 000~2 560 m(郑万钧,1997),与此次调查样地的海拔范围较接近;青榨槭天然分布的海拔范围是500~2 100 m,所以其生态位宽度较别的树种大;大叶青冈、小叶青冈和虎皮楠等树种天然分布的海拔多在1 800 m以下,铁杉多分布在海拔1 800 m以上(吴征镒,1983)。植物长期适应的结果决定了树种只能在一定的海拔范围内生存,超出这一范围其适应能力就会受到限制,这也是影响树种生态位宽度的因素之一。
各优势种群的生态位宽度与其重要值具有明显的正相关,其相关系数达到0.988 72(Shannon-Wiener多样性指标)和0.825 279(Levins生态位宽度指数)。崖柏的重要值和生态位宽度都显著地高于其他物种,表明其在群落中处于优势地位;川陕鹅耳枥、高山栎和华千金榆的重要值和生态位宽度较大,表明它们在创造以崖柏为优势种的群落环境中起着重要作用。
3.3.2 垂直生态位宽度垂直生态位宽度(高度生态位)表明不同物种利用随着高度变化的以光因子为主导的生态资源的能力,垂直生态位宽度大,表明它们的适应幅度较大,不同高度的个体在各资源位内分布较为均匀。
由表 3可以看出,优势乔木种群的垂直生态位宽度排序为:高山栎>乌岗栎>青榨槭>崖柏>华千金榆>大叶青冈>铁杉>中华槭>城口青冈>虎皮楠>华中八角>小叶青冈>川鄂山茱萸>华西花楸>川陕鹅耳枥。
在崖柏群落中,崖柏种群的垂直生态位排在第4位。与前3个树种比较,11 m以上的崖柏结实母树以及2 m以下的崖柏幼苗、幼树较少,是崖柏垂直生态位宽度值低于高山栎、乌岗栎、青榨槭等的主要原因。
3.4 生态位重叠 3.4.1 水平生态位重叠生态位重叠是指群落中的资源被2个或多个种群同时共同分享,它是种群的一种生态特性,它既能反映不同种群同时利用同一资源的情况,又能反映这些生态位重叠的种群在某些生态因子联系上的相似性(吴淑芳,2003;臧润国等,2005)。对崖柏群落中15个优势乔木种群的水平生态位重叠值进行计算,结果见表 4。
|
|
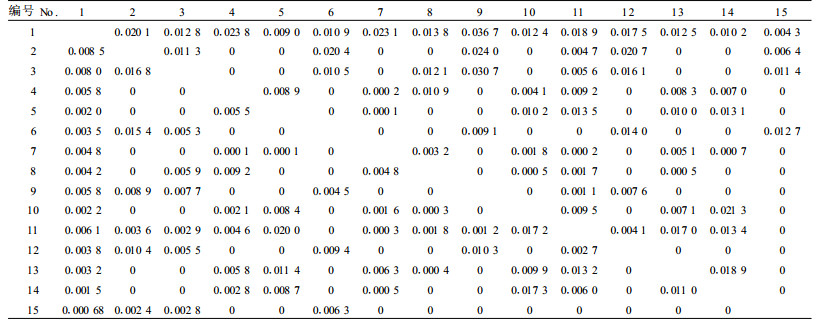
由表 4可以看出,崖柏与其他乔木种群均发生生态位重叠,其中,崖柏对其他树种的生态位重叠值(Lih值)较大,集中在0.004 33~0.036 73,而其他树种对崖柏的生态位重叠值(Lhi值)相对较小,集中在0.000 68~0.008 52。崖柏对其他树种的生态位重叠较大的现象表明:崖柏和这些种群存在着一定的资源利用性竞争,而且崖柏具有较强的竞争能力,产生这种现象的原因与崖柏具有较大的生态位宽度有关。一般认为,生态位宽度较大的物种由于对资源的利用能力较强、分布较广而与其他种群间的生态位重叠较大(胡正华等,2004)。对一个种对而言,前一个种对后一个种的重叠值较高,但后种对前种的值则不一定高,如崖柏对铁杉的生态位重叠值为0.036 73,而铁杉对崖柏的生态位重叠值为0.005 77,这是因为此生态位重叠值是由Levins生态位宽度值指标而定的,一般规律是生态位宽度值大的种类对生态位宽度较窄的种类可能有较高的重叠值,反之则低(臧润国等,2005)。其他树种对崖柏的生态位重叠值相对较小,反映出其他树种对崖柏所能利用的环境资源的利用比较少。
对照表 3,将崖柏对其他树种的生态位重叠值按大小排序为:铁杉>华千金榆>小叶青冈>高山栎>青榨槭>华西花楸>城口青冈>川陕鹅耳枥>乌岗栎>虎皮楠>华中八角>川鄂山茱萸>大叶青冈>中华槭。崖柏对其他树种生态位重叠值的大小排序反映了崖柏与这些树种对生态条件要求的相近程度,生态位重叠值越大,说明它们与崖柏对生态条件的要求越相似。以生态位重叠值最大的铁杉为例,铁杉自然分布的生境范围非常广阔,它可以从亚高山至中山,跨越相当于温带至暖温带和亚热带的气候;但其分布生境中有一个重要的特征,即全年温差较小,气候温和、湿润乃至比较潮湿,地形以狭窄间谷或山地云雾积聚的坡地为主,这与崖柏生存地域的气候条件和地形条件类似。对于其他树种对崖柏的生态位重叠较小的原因,同样也可以从树种对生态环境的需求得到解释。比如崖柏与大叶青冈,二者的主要区别在于大叶青冈属于喜温暖的常绿阔叶树种,而崖柏属于耐温凉的树种(蒋有绪等,1998),所以它们的生态位重叠值较小。
一般认为,生境要求比较相近的种群间生态位重叠较大;但也有研究表明,同属的种群间由于生物学生态学特征相似,在一定程度上存在对环境资源需求的分化,从而会导致生态位重叠程度降低,以便使它们共存于某一小生境中(陈波等,1995)。本研究中同属的大叶青冈、小叶青冈和城口青冈相互间的生态位重叠值为0.000 06~0.004 75;川陕鹅耳枥和华千金榆、高山栎和乌岗栎以及青榨槭和中华槭相互间的重叠值为0就属于这种情况。
将崖柏群落中优势乔木树种的生态位重叠值与生态位宽度大小进行比较,可以发现生态位宽度与生态位重叠之间的确存在“生态位宽度值较大的物种与其他种群间的生态位重叠值也大”这种现象,如崖柏和高山栎;但也存在一些个例,如崖柏与铁杉,这种现象与史作民等(1999)提出“具有较大生态位宽度的物种间生态位重叠值不一定高”的结论相一致。主要原因在于树种的生物生态学特性不一定相同,它们对资源的要求不完全一样。
3.4.2 垂直生态位重叠对崖柏群落15个优势种群的垂直生态位重叠进行计算的结果见表 5。根据表 5可以将优势种群间垂直生态位重叠划分为0、0.001~0.009、0.01~0.09及0.1~0.9这4个层次。重叠值介于0.1~0.9的有9个,重叠值为0的有4个,重叠值介于0.01~0.09的有32个,重叠值介于0.001~0.009的有165个。崖柏对其他树种和其他树种对崖柏的生态位重叠值大小差异不大,集中在0.004~0.084。
|
|

其他树种对崖柏的垂直生态位重叠值按大小排序为:乌岗栎>城口青冈>高山栎>铁杉>小叶青冈>华千金榆>大叶青冈>青榨槭>华中八角>中华槭>华西花楸>虎皮楠>川陕鹅耳枥>川鄂山茱萸;崖柏对其他树种的垂直生态位重叠值按大小排序为:小叶青冈>城口青冈>乌岗栎>铁杉>川陕鹅耳枥>华千金榆>大叶青冈>华中八角>华西花楸>高山栎>青榨槭>中华槭>虎皮楠>川鄂山茱萸。
这些结果表明,在崖柏群落中,随着高度变化许多乔木树种与崖柏存在以光因子为主导的生态资源利用方面的竞争,但竞争程度不同。这一结果对于指导人工干预其他物种,救护崖柏种群具有理论指导意义。
4 结论与讨论1) 崖柏群落中优势种群水平生态位宽度值(Levins生态位宽度)较大的包括崖柏、川陕鹅耳栎、高山栎和华千金榆,较小的包括虎皮楠、铁杉、中华槭和川鄂山茱萸。各种群的水平生态位宽度与重要值呈明显的正相关。垂直生态位宽度值较大的包括高山栎、乌岗栎、青榨槭和崖柏,较小的包括小叶青冈、川鄂山茱萸、华西花楸和川陕鹅耳栎。
2) 崖柏种群对其他种群的水平生态位重叠值高于其他种群对崖柏种群的生态位重叠值,说明崖柏种群具有较强的竞争能力;崖柏群落优势乔木种群间水平生态位重叠值较小,反映出现存崖柏群落结构具有一定的稳定性。
3) 乌岗栎、城口青冈、高山栎和铁杉对崖柏的垂直生态位重叠值较大;崖柏对小叶青冈、城口青冈、乌岗栎和铁杉的垂直生态位重叠值较大。这一结果表明,在崖柏群落的优势乔木种群之间存在着以光因子为主导的生态资源竞争。以不同树种与崖柏之间的垂直生态位重叠为参考,采取必要的人工干预措施,对于拯救这一极度濒危物种具有重要意义。
4) 崖柏在现存群落中具有较强的竞争力,但已有研究表明崖柏结实存在严重的大小年现象,且每个球果的出种量仅为3粒,无种子的球果占7.1%(王祥福等,2007),种子活力仅为8%(郭泉水等,2007)。综上所述,建立保护区和开展崖柏无性繁殖研究是扩繁该物种的有效手段。另外开展崖柏引种研究,扩大其分布范围也是非常必要的。
5) 目前,虽然人们理解生态位的概念比较容易,但如何测度则相当困难。对生态位的定量研究,大都是根据所研究种群能利用的空间或资源来划分物种能利用的资源状态等级,然后利用不同的计测公式计算生态位的宽度和重叠。基于单一资源轴的生态位宽度计测研究目前在国内还属多数,但随着多维生态位计测在方法上的不断突破,今后,多维生态位研究将会受到更多关注,而多元统计分析方法将在其中扮演重要角色(李德志等,2006b)。目前用于计算各个种群的生态位宽度和生态位重叠的指标除广泛应用不同取样样地的个体数目外,胸高断面积、生物量或重要值也有应用,但多种指标的综合比较和应用较少,今后这些问题有待进一步研究。本研究所调查的对象仅为以崖柏为优势的群落,开展崖柏零散分布区及该地区以其他物种为优势群落的生态位研究,将会为保护和发展这一极度濒危物种提供更好的理论指导。
毕润成, 尹文兵, 王艳妮. 2003. 山西南部脱皮榆种群生态位的研究. 西北植物学报, 23(7): 1266-1271. DOI:10.3321/j.issn:1000-4025.2003.07.039 |
陈波, 周兴民. 1995. 三种嵩草群落中若干植物种的生态位宽度与重叠分析. 植物生态学报, 19(2): 158-169. DOI:10.3321/j.issn:1005-264X.1995.02.007 |
郭泉水, 王祥福, 巴哈尔古丽, 等. 2007. 崖柏群落优势乔木树种种间关系. 生态学杂志, 26(12): 1911-1917. |
胡正华, 于明坚, 彭传正, 等. 2004. 古田山国家自然保护区黄山松林主要种群生态位研究. 生态环境, 13(4): 619-621, 629. DOI:10.3969/j.issn.1674-5906.2004.04.043 |
黄英姿. 1994. 生态位理论研究中的数学方法. 应用生态学报, 5(3): 331-337. |
侯琳, 雷瑞德, 王得祥, 等. 2006. 黄龙山林区封育天然油松群落种群生态位特征. 西北植物学报, 26(3): 585-591. DOI:10.3321/j.issn:1000-4025.2006.03.026 |
蒋有绪, 郭泉水, 马娟, 等. 1998. 中国森林群落分类及其群落学特征. 北京: 科学出版社, 中国林业出版社, 55-56.
|
李德志, 刘科轶, 臧润国, 等. 2006a. 现代生态位理论的发展及其主要流派. 林业科学, 42(8): 88-94. |
李德志, 石强, 臧润国, 等. 2006b. 物种或种群生态位宽度与生态位重叠的计测模型. 林业科学, 42(7): 95-103. |
李生, 陈存及, 曹永慧, 等. 2003. 乳源木莲天然林主要种群生态位的研究. 江西农业大学学报:自然科学版, 25(3): 374-378. |
刘建锋, 江泽平, 肖文发, 等. 2005. 极度濒危植物——崖柏种群空间格局与动态的初步研究. 江西农业大学学报:自然科学版, 27(5): 708-712. |
刘建锋, 肖文发, 郭志华, 等. 2004. 珍稀濒危植物——崖柏种群结构与动态初步研究. 江西农业大学学报:自然科学版, 26(3): 377-380. |
刘金福, 洪伟. 1999. 格氏栲群落生态学研究——格氏栲林主要种群生态位的研究. 生态学报, 19(3): 347-352. DOI:10.3321/j.issn:1000-0933.1999.03.010 |
柳江, 洪伟, 吴承祯, 等. 2002. 退化红壤区植被恢复过程中灌木层主要种群的生态位特征. 植物资源与环境学报, 11(2): 11-16. DOI:10.3969/j.issn.1674-7895.2002.02.003 |
任青山. 1998. 天然次生林主要种群生态位结构的研究. 东北林业大学学报, 26(2): 5-10. |
史作民, 程瑞梅, 刘世荣. 1999. 宝天曼落叶阔叶林种群生态位特征. 应用生态学报, 10(3): 265-269. DOI:10.3321/j.issn:1001-9332.1999.03.003 |
宋永昌. 2001. 植被生态学. 上海: 华东师范大学出版社, 45-46.
|
唐熙, 李振宇, 胡玉熹. 2005. 中国特有濒危植物崖柏的木材结构研究. 武汉植物学研究, (2): 149-153. DOI:10.3969/j.issn.2095-0837.2005.02.010 |
王琳, 张金屯. 2001. 濒危植物矮牡丹的生态位研究. 生态学杂志, 20(4): 65-69. DOI:10.3321/j.issn:1000-4890.2001.04.017 |
王祥福, 郭泉水, 巴哈尔古丽, 等. 2007. 世界级极危物种——崖柏球果特征及出种量的研究. 林业科学研究, 20(5): 673-677. DOI:10.3321/j.issn:1001-1498.2007.05.014 |
吴刚, 梁秀英, 张旭东, 等. 1999. 长白山红松阔叶林主要树种高度生态位的研究. 应用生态学报, 10(3): 262-264. DOI:10.3321/j.issn:1001-9332.1999.03.002 |
吴淑芳. 2003. 海拔资源空间天然杉木与主要伴生树种的生态位及种间竞争. 江西农业大学学报:自然科学版, 25(3): 269-373. |
吴征镒. 1983. 中国植被. 北京: 科学出版社, 857-856.
|
张峰, 上官铁梁. 2004. 翅果油树群落优势种群生态位分析. 西北植物学报, 24(1): 70-74. DOI:10.3321/j.issn:1000-4025.2004.01.013 |
张继义, 赵哈林, 张铜会, 等. 2003. 科尔沁沙地植物群落恢复演替系列种群生态位动态特征. 生态学报, 23(12): 2741-2746. DOI:10.3321/j.issn:1000-0933.2003.12.030 |
臧润国, 成克武, 李俊清, 等. 2005. 天然林生物多样性保育与恢复. 北京: 中国科学技术出版社, 213-214.
|
郑万钧. 1997. 中国树木志(第二卷). 北京: 中国林业出版社, 2190-2191.
|
Aplet G H, Vitosek P M. 1994. An age-altitude matrix analysis of Hawaii an Rain forest succession. Journal of Ecology, 82(1): 137-147. DOI:10.2307/2261393 |
Fu L K, Jin J M. 1992. China plant red data book rare and endangered plant 1. Beijing: Science Press.
|
Fu L K, Yu Y F, Farjon A. 1999. Cupressaceae//Wu Z Y, Raen P H. Flora of China 4. Beijing: Science Press, 3.
|
Grinell J. 1917. The niche relationship of California Thrsher. Auk, 1: 64-82. |
Hurlbert S H. 1978. The measurement of niche overlap and some relatives. Ecology, 9(1): 67-77. |
Leibold M A. 1995. The niche concept revisited mechanistic models and community context. Ecology, 76(5): 1371-1382. DOI:10.2307/1938141 |
Weider L J. 1993. Niche breadth and life history variation in a hybrid Daphnia complex. Ecology, 74(3): 935-943. DOI:10.2307/1940817 |
Xiang Q P, Farjon A, Li Z Y, et al. 2002. Thuja sutchuenensis: a rediscovered species of the Cupressaceae. Botanical Journal of Linnean Society, 139: 305-310. DOI:10.1046/j.1095-8339.2002.00055.x |