



多环芳烃主要是以附着于颗粒物质的形式存在于大气中,通过呼吸、饮食、皮肤接触等途径进入人体,不仅有致癌、致突变作用和遗传毒性,而且可以通过胎盘与DNA结合形成加合物,导致胎儿DNA损伤,影响胎儿生长发育。母体孕期暴露于高浓度的多环芳烃与低出生体重、胎儿生长受限、早产、出生缺陷等不良出生结局有关[1-6]。部分致癌性多环芳烃(PAHs)通过胎盘,可以引发胎儿肺、肝脏、淋巴组织和神经系统肿瘤[7-8]。然而其生物学作用机制还有待进一步研究。各种遗传学损伤都会通过基因表达来指导细胞功能/个体表型的表现,因此基因表达是各种遗传损伤和细胞功能/个体表型之间的桥梁。BaP是主要的致癌性多环芳烃之一,本研究将分析宫内BaP暴露对胎儿几个癌症相关基因表达的改变,从而分析其可能的作用机制和潜在的健康影响。
1 材料与方法 1.1 主要仪器与试剂Waters 2960高效液相色谱仪及色谱工作站(美国)、Waters 474荧光检测器(美国)、Supelco固相萃取装置(美国)、3 mL 500 mg C18固相萃取小柱(美国Supelco公司)、QIANGEN Rotor-Gene Q实时荧光定量PCR仪(德国)。
QIAGEN RNeasy Mini Kit试剂盒(德国)、TaKaRa SYBR Green Realtime PCR Master Mix试剂盒(日本)、BaP标准品(美国Supelco公司)、甲醇(色谱纯,德国Merck公司)等。
1.2 样品的采集及保存2010年3月—4月在太原市某二级甲等综合性医院随机收集20个新生儿的脐带血各10 mL,其中2 mL脐带血冻存于-80℃低温冰箱中,用于RNA的提取。8 mL脐带血当天分离血清,-70℃避光保存,用于血清BaP的检测。
1.3 血清多环芳烃测定用高效液相色谱—荧光检测法测定血清中BaP的含量。1 mL血清经固相萃取浓缩至100 μL甲醇中,采用梯度淋洗法进行检测。荧光检测器Ex = 280 nm,Em = 389 nm,进样量为10 μL,流速为0.8 mL/min,流动相为乙睛:水(0 min: 60% : 40%,20 min: 60% : 40%,35 min: 100% : 0,45min: 60% : 40%)。
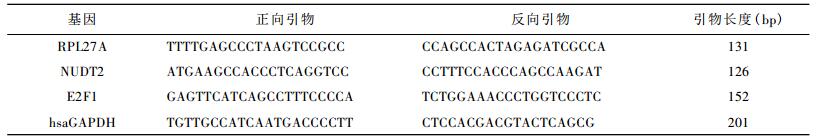
1.4 Real-time PCR基因表达检测用QIAGEN RNeasy Mini Kit试剂盒进行总RNA提取。根据GeneBank数据库中各基因的序列信息,使用引物设计软件primer expression 2.0设计RPL27A、NUDT2、E2F1和内参基因hsaGAPDH的PCR引物,引物由上海英骏生物技术有限公司合成(表 1)。
采用SYBR染料法检测基因的相对表达量,所用试剂盒为TaKaRa SYBR Green Realtime PCR Master Mix。
PCR扩增,反应过程及循环条件:预变性(95℃,30 s),变性(95℃,5 s),退火(60℃,34 s),延伸(72℃,30 s),40个循环。
采用2-⊿⊿Ct法计算mRNA的表达量:
⊿Ct=Ct目的基因-Ct内参基因
⊿⊿Ct=⊿Ct暴露组-⊿Ct对照组
相对表达量= 2(-⊿⊿Ct)
Ct:指荧光信号达到设定的阈值时所经历的循环数。
1.5 统计分析用Excel建立数据库,用SAS 9.1进行统计分析。
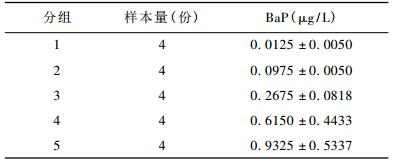
2 结果 2.1 脐带血BaP水平20份脐带血血清BaP平均浓度为0.38 ± 0.31 μg/L。按照血清BaP浓度,将20份样本分成5个不同水平的暴露组,各暴露组的BaP水平见表 2。
2.2 不同暴露组基因表达水平与BaP暴露水平的相关关系
以第1组为对照组,计算其他各暴露组的RPL27A、NUDT2和E2F1的mRNA相对表达量,分析不同暴露组BaP平均水平与各基因相对表达量的相关关系(见图 1-3)。由图 1-3可见,随着BaP暴露水平的增高,RPL27A和NUDT2呈现表达上调的趋势,而E2F1呈现表达下调的趋势(P<0.0001)。

|
| 图 1 不同暴露组BaP平均水平与RPL27A相对表达量的相关关系 |

|
| 图 2 不同暴露组BaP平均水平与NUDT2相对表达量的相关关系 |

|
| 图 3 不同暴露组BaP平均水平与E2F1相对表达量的相关关系 |
3 讨论
本研究初步探讨了宫内BaP暴露对RPL27A、NUDT2和E2F1 3个癌症相关基因表达的影响。
RPL27A,定位于染色体11p15,该基因编码的蛋白是核糖体60S的构成蛋白之一,属于核糖体蛋白的L15P家族,位于细胞质内。在RPL27A基因第一个内含子的阅读框有翻译起始密码子ATG,该基因亦保存有转录调控因子、Box-A和GABP序列[9]。基因结构决定基因功能,RPL27A具有转录活性,参与真核细胞翻译的起始、延长和终止,可调控细胞周期。有研究报道该基因在肺癌和结肠癌组织出现表达异常[10]。
随BaP暴露水平增加,RPL27A mRNA表达水平增加,说明RPL27A可能是BaP引发肿瘤的作用机制或靶点之一。
NUDT2位于染色体9p13,该基因编码的蛋白属于MutT核苷焦磷酸酶家族的成员之一,是NUDIX水解酶超家族的一个分支[11]。该基因产物参与特定核苷焦磷酸酶的MutT序列的修饰,而MutT酶可通过水解致突变核苷结合底物如8-oxo-dGTP而防止DNA复制错误[12]。NUDT2是候选抑癌基因。NUDT2蛋白可水解二腺苷5’,5’- p1,p4-四磷酸盐(Ap4A)为AMP和ATP,以维持细胞内Ap4A的动态平衡。Ap4A与许多细胞功能有关,如参与调控细胞增殖、DNA复制和修复、凋亡等。在细胞受到热、氧化或DNA损伤刺激时,细胞内的Ap4A水平会增加[13-14],从而诱导细胞凋亡。NUDT2是雌激素抑制基因,在乳腺癌细胞中通过HER2 pathway而被诱导表达。其高表达与乳腺癌的不良临床症状有关,可促进乳腺癌细胞增殖,是乳腺癌的预后因子之一[15]。
随BaP暴露水平增加,NUDT2 mRNA表达水平增加,可能是由于BaP引起的DNA损伤诱导了具有DNA损伤修复调控作用的抑癌基因NUDT2的表达,说明NUDT2也可能是BaP引发肿瘤的机制之一。
E2F1基因定位于20q11.2,该基因编码的蛋白属于转录因子E2F家族。E2F家族在细胞周期调控中发挥重要作用,是抑癌基因,也是小DNA肿瘤病毒转化蛋白的靶对象。E2F蛋白包含几个保守结构域,包括DNA结合结构域、与转录因子蛋白(DP)相互作用的二聚体结构域、富含酸性氨基酸的反式激活结构域和肿瘤抑制相关结构域。E2F1可调控细胞增殖,参与p53依赖性和非依赖性凋亡(p53-dependent/independent apoptosis) [16]。E2F1的高表达可以激活ATM \ ATR \ CHK1和CHK2等激酶的活性,促使p53 N端丝氨酸残基磷酸化而被激活,从而诱导DNA损伤修复、细胞周期阻滞或细胞凋亡[17-20]。E2F1也可以直接反式激活p73启动子,而诱导细胞周期阻滞和凋亡[21-22]。E2F1和c-Myc还可通过诱导转录和调控信号转导通路来调控细胞生长和死亡[23]。
然而,E2F1也可以抑制凋亡、促进癌细胞存活而发挥癌基因的作用[24]。有研究发现E2F1、NF-κB和EGR1在前列腺癌中出现过表达(overexpression),对细胞增殖、存活和转移起重要作用[25-30]。Zheng等[31]的研究表明,E2F1可与NF -κB(nuclear factor-κB)相互作用,诱导Egr1基因的转录表达,而EGR1蛋白增多可以诱导一系列生长因子的生成,包括EGF (epidermal growth factor)、PDGF (platelet-derived growth factor)和IGF-Ⅱ (insulin-like growth factor Ⅱ)等,这些生长因子可激活重要的细胞生存信号通路(survival pathway)-PI3K/AKT信号通路(Phosphoisitide-3-kinase/AKT pathway),从而促进抵抗药物诱导的前列腺癌细胞的凋亡。
细胞凋亡或凋亡抑制是DNA损伤修复失败后的两种重要的竞争性的结局,细胞凋亡是自主性的细胞清除过程,是为了更好地适应生存环境而主动争取的一种死亡过程。细胞凋亡抑制是癌症发生的重要机制之一。E2F1做为凋亡调控因子,究竟发挥癌基因还是抑癌基因的作用,可能主要取决于其下游基因的存在/缺失或状态。
在本研究中,随着致癌物质BaP暴露水平的增加,E2F1表达下调,可能会有两种情况,第一种情况,如果E2F1诱导凋亡,发挥抑癌基因的作用,那么,随着宫内BaP暴露水平的增加,E2F1表达下调,说明宫内BaP暴露可使E2F1诱导凋亡的作用减弱,则在一定程度上可能会增加以后患癌的风险; 另一种情况,如果E2F1抑制凋亡,发挥癌基因的作用,那么,随着宫内BaP暴露水平的增加,E2F1表达下调,则可能是由于致癌物BaP引起机体反馈性的损伤修复机制,如非E2F1诱导的p53依赖的损伤修复,由于p53的过度表达,可以反馈性地抑制E2F1的表达,这种损伤修复的反应也说明了宫内BaP暴露可能会给胎儿带来不良影响,E2F1可能是BaP引发不良影响的作用机制或靶点之一。
低剂量、持续的环境污染暴露对健康的影响已越来越引起人们的重视,苯并(a)芘等多环芳烃来源广泛,污染普遍存在,其对人群的健康影响是不容忽视的。本研究仅初步探讨了苯并(a)芘对最敏感人群—生命初期阶段可能的分子水平的影响,为今后进一步深入研究起抛砖引玉的作用。
| [1] | Perera FP, Rauh V, Tsai WY, et al. Effects of transplacental exposure to environmental pollutants on birth outcomes in a multiethnic pollution[J]. Environ Health Perspect, 2003, 111(2): 201–205. |
| [2] | Perera FR, Rauh V, Whyatt RM, et al. Molecular evidence of an interaction between prenatal environmental exposures and birth outcomes in a multiethnic population[J]. Environ Health Perspect, 2004, 112(5): 626–630. doi: 10.1289/ehp.6617 |
| [3] | Choi H, Rauh V, Garfinkel R, et al. Prenatal exposure to airborne polycyclic aromatic hydrocarbons and risk of intrauterine growth restriction[J]. Environ Health Perspect, 2008, 116(5): 658–665. doi: 10.1289/ehp.10958 |
| [4] | Choi H, Jedrychowaki W, Spengler J, et al. International studies of prenatal exposure to polycyclic aromatic hydrocarbons and fetal growth[J]. Environ Health Perspect, 2006, 114(11): 1744–1750. |
| [5] | Dejmek J, Solansky I, Benes I, et al. The impact of polycyclic aromatic hydrocarbons and fine particles on pregnancy outcome[J]. Environ Health Perspect, 2000, 108(12): 1159–1164. doi: 10.1289/ehp.001081159 |
| [6] | Tang D, Li TY, Liu JJ, et al. PAH-DNA adducts in cord blood and fetal and child development in a Chinese cohort[J]. Environ Health Perspect, 2006, 114(8): 1297–1300. doi: 10.1289/ehp.8939 |
| [7] | Youndai EV, Holloway AC, Fobter WG. Environmental and occupational factors affecting fertility and IVF success[J]. Hum Reprod Update, 2005, 11(1): 43–57. |
| [8] | 侯海燕, 王丹. 多环芳烃对胎儿和出生结局的影响[J]. 国际妇产科学杂志, 2008, 35(3): 171–174. |
| [9] | Kusuda J, Hirai M, Tanuma R, et al. Genomic structure and chromosome location of RPL27A/Rpl27a, the genes encoding human and mouse ribosomal protein L27A[J]. Cytogenet Cell Genet, 1999, 85(3-4): 248–251. doi: 10.1159/000015303 |
| [10] | De Felice B, Ciarmiello LF, Wilson RR. Identification of a cDNA clone encoding DIP1-binding protein in Drosophila melanogaster[J]. Mol Biol Rep, 2004, 31(3): 165–169. doi: 10.1023/B:MOLE.0000043554.10462.17 |
| [11] | 林军, 胡沂淮, 田兵, 等. 双MutT/Nudix结构域蛋白的进化:相互独立的基因重复—融合事件形成类似的结构域组成[J]. 遗传学报, 2009, 36(10): 603–610. |
| [12] | Mildvan AS, Weber DJ, Abeygunawardana C. Solution structure and mechanism of the MutT pyrophosphohydrolase[J]. Adv Enzymol Relat Areas Mol Biol, 1999, 73: 183–209. |
| [13] | Vasilenko N, Moshynskyy I, Zakhartchouk A. SARS coronavirus protein 7a interacts with human Ap4A-hydrolase[J]. Virol J, 2010, 7: 31. doi: 10.1186/1743-422X-7-31 |
| [14] | Kisselev LL, Justesen J, Wolfson AD, et al. Diadenosine oligophosphates (Ap (n) A), a novel class of signaling molecules[J]. FEBS Lett, 1998, 427(2): 157–163. doi: 10.1016/S0014-5793(98)00420-7 |
| [15] | Oka K, Suzuki T, Onodera Y, et al. Nudix-type motif 2 in human breast carcinoma: a potent prognostic factor associated with cell proliferation[J]. Int J Cancer, 2011, 128(8): 1770–82. doi: 10.1002/ijc.25505 |
| [16] | Gurtner A, Fuschi P, Martelli F, et al. Transcription factor NF-Y induces apoptosis in cells expressing wild-type p53 through E2F1 upregulation and p53 activation[J]. Cancer Res, 2010, 70(23): 9711–9720. doi: 10.1158/0008-5472.CAN-10-0721 |
| [17] | Roqoff HA, Pickering MT, Debatis ME, et al. E2F1 induces phosphorylation of p53 that is coincident with p53 accumulation and apoptosis[J]. Mol Cell Biol, 2002, 22(15): 5308–5318. doi: 10.1128/MCB.22.15.5308-5318.2002 |
| [18] | Powers JT, Hong S, Mayhew CN, et al. E2F1 uses the ATM signaling pathway to induce p53 and Chk2 phosphorylation and apoptosis[J]. Mol Cancer Res, 2004, 2(4): 203–214. |
| [19] | Shieh SY, Ahn J, Tamai K, et al. The human homologs of checkpoint kinases Chk1 and Cds1 (Chk2) phophorylate p53 at multiple DNA damage-inducible sites[J]. Genes Dev, 2000, 14(3): 289–300. |
| [20] | Zhou BB, Elledge SJ. The DNA damage response: putting checkpoints in perspective[J]. Nature, 2000, 408(6811): 433–439. doi: 10.1038/35044005 |
| [21] | Irwin M, Marin MC, Phillips AC, et al. Role for the p53 homologue p73 in E2F1-induced apoptosis[J]. Nature, 2000, 407: 645–648. doi: 10.1038/35036614 |
| [22] | Stiewe Tand, Puzer BM. Role of the p53-homologue p73 in E2F1-induced apoptosis[J]. Nat. Genet, 2000, 26(4): 464–469. doi: 10.1038/82617 |
| [23] | Matsumura L, Tanaka H, Kanakura Y. E2F1 and c-myc in cell growth and death[J]. Cell Cycle, 2003, 2(4): 333–338. |
| [24] | Ginsberg D. E2F1 pathways to apoptosis[J]. FEBS Lett, 2002, 529(1): 122–125. doi: 10.1016/S0014-5793(02)03270-2 |
| [25] | Abdulkadir SA, Qu Z, Garabedian E, et al. Impaired prostate tumorigenesis in Egr1-deficient mice[J]. Nat Med, 2001, 7(1): 101–107. doi: 10.1038/83231 |
| [26] | Baron V, De Gregorio G, Krones-Herzig A, et al. Inhibition of Egr-1 expression reverses transformation of prostate cancer cells in vitro and in vivo[J]. Oncogene, 2003, 22(27): 4194–4204. doi: 10.1038/sj.onc.1206560 |
| [27] | Virolle T, Krones-Herzig A, Barron V, et al. Egr1 promotes growth and survival of prostate cancer cells [J]. Identification of novel Egr1 target genes[J]. J Biol Chem, 2003, 278 (14) : 11802-11810. |
| [28] | Agus DB, Cordon-Cardo C, Fox W, et al. Prostate cancer cell cycle regulators: response to androgen withdrawal and development of androgen independence[J]. J Natl Cancer Inst, 1999, 91(21): 1869–1876. doi: 10.1093/jnci/91.21.1869 |
| [29] | Suh J, Payvandi F, Edelstein LC, et al. Mechanisms of constitutive NF-Κb activation in human prostate cancer cells[J]. Prostate, 2002, 52(3): 183–200. doi: 10.1002/pros.v52:3 |
| [30] | Davis JN, Wojno KJ, Daignault S, et al. Elevated E2F1 inhibits transcription of the androgen receptor in metastatic hormoneresistant prostate cancer[J]. Cancer Res, 2006, 66(24): 11897–11906. doi: 10.1158/0008-5472.CAN-06-2497 |
| [31] | Chao guZheng, Zijia Ren, Hetian Wang, et al. E2F1 induces tumor cell survival via nuclear factor-κB dependent induction of EGR1 transcription in prostate cancer cells[J]. Cancer Res, 2009, 69(6): 2324–2331. doi: 10.1158/0008-5472.CAN-08-4113 |