 2019, Vol. 39
2019, Vol. 39



2. 重庆大学环境科学系, 重庆 400044
2. Department of Environmental Science, Chongqing University, Chongqing 400044
近年来, 由于工农业废水的排放, 导致我国地表水体氮素含量不断升高, “水华”、“赤潮”现象频发, 对农业、渔业、旅游业等造成了巨大影响.目前, 硝化-反硝化生物脱氮工艺普遍面临有机碳源不足、反硝化不充分、总氮达标难度大的困境, 而额外投加碳源不仅大大增加了废水脱氮成本, 且会进一步引起温室气体甲烷的大量排放(Solomon et al., 2007; 沈李东等, 2011), 加剧温室效应.鉴于此, 低碳耗脱氮技术开发或者经济碳源替代成为可能的解决途径.
硝态氮型厌氧甲烷氧化(NOx-Dependent Anaerobic Methane Oxidation, NOx-DAMO)是一种新兴的生物脱氮过程, 其以硝酸盐或亚硝酸盐为电子受体, 以甲烷为电子供体, 可以同步脱除氮素污染并缓解温室效应.NOx-DAMO过程的反应式如下所示:
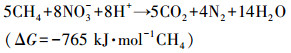
|
(1) |
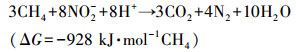
|
(2) |
2006年, 科研人员首次富集了该类微生物, 并通过同位素标记法证实ANME(anaerobic methanotrophic)古菌和NC10门细菌均参与了反应(Raghoebarsing et al., 2006).随后, Ettwing等(2008)证实了归属于NC10门的细菌M.oxyfera (Candidatus Methylomirabilis oxyfera)可独立完成厌氧甲烷氧化过程, 该生物过程被称为亚硝酸盐型甲烷厌氧氧化(n-DAMO).同时, 研究人员利用分子生物学技术揭示了n-DAMO细菌在淡水湿地(Han et al., 2013; Hu et al., 2014a)、水库消落带(Wang et al., 2016)、南海底泥(Chen et al., 2014)及污水处理厂污泥(Luesken et al., 2011; Kampman et al., 2014)等自然及人工生态系统中广泛存在, 并以各种接种物成功富集n-DAMO功能微生物, 并发现M.oxyfera菌相对丰度可达15%~80%.然而, M.oxyfera细菌生长速率缓慢, 脱氮速率有限, 成为限制n-DAMO过程进一步工程应用的瓶颈.针对以上问题, 研究人员通过调整温度(Hu et al., 2009; He et al., 2015a; 赵荣等, 2017)、pH值(Zhu et al., 2012)、基质浓度(Lieberman et al., 2004; He et al., 2016)、微量元素(Lu et al., 2016)等培养条件对其进行优化.同时, 反应器构型改造(Hu et al., 2014b)、电辅助(Wu et al., 2016a)等也被用于强化n-DAMO过程.
由于n-DAMO微生物增殖缓慢, 富集量少, 因此, 研究者都在尽量保存生物量, 极少研究排泥对n-DMAO活性的影响.但培养体系中存在少量其他竞争微生物和抑制性产物, 均会对n-DAMO微生物的生长产生干扰, 故可以通过排泥的方式对n-DAMO微生物进行加速富集并提高活性.基于此, 本研究以实验室稳定运行的序列式活性污泥法反应器(SBR)中n-DAMO功能微生物为研究对象, 考察排泥对n-DAMO微生物及脱氮过程的影响, 并利用荧光定量PCR技术、高通量测序技术分析功能微生物与其余微生物群落结构变化, 进一步解析内在的微生物学机制, 以期进一步丰富n-DAMO过程的强化策略.
2 材料与方法(Materials and methods) 2.1 反应器构型实验所采用的反应装置如图 1所示(柴风光等, 2018; Li et al., 2018), 反应器材质为双层玻璃, 有效容积为2 L, 反应器内液相体积占1.5 L, 气相体积占0.5 L, 外隔层为水浴区.反应器内置搅拌子, 使用磁力搅拌器进行搅拌.
 |
| 图 1 SBR示意图 Fig. 1 Diagram of sequence batch reactor |
本研究以人工景观湖底泥(60 mL)、城市污水处理厂污泥浓缩池厌氧污泥(400 mL)、稻田土沉积物(取表层泥以下10 cm处污泥450 mL)混合物为接种物.接种污泥沉淀并弃去上清液, 再混合均匀过10目筛, 在连续曝氮气条件下用纯水冲洗污泥3~5次.最后取380 mL装入SBR反应器中, 并加入1.5 L培养液(柴风光等, 2018; Li et al., 2018), 成分如表 1所示(Ettwig et al., 2009).矿物基质、酸性微量元素、碱性微量元素分开配制, 每升基质中添加0.5 mL酸性微量元素(包含0.1 mol · L-1 HCl)、0.2 mL碱性微量元素(包含0.1 mol · L-1 NaOH).
| 表 1 微量元素与矿物基质表 Table 1 Trace elements and mineral matrix |
设置两组相同的SBR反应器, 分别在严格厌氧条件下运行, 其中, 实验组记为1号, 对照组记为2号.维持反应器中亚硝酸盐浓度为30 mg · L-1, 水力停留时间为90 d, 反应温度为30 ℃, pH通过1 mol · L-1的HCl维持在7.0~7.5之间.1号反应器在实验第42 d排泥500 mL, 排泥比为33.3%;在第172 d排泥250 mL, 排泥比为16.7%.实验期间两只反应器每天曝气10 min(体积分数占比:5%N2、90%CH4、5%CO2)(Lu et al., 2019), 曝气速率为150 mL · min-1, 反应器内压力维持在1.01×105~1.11×105 Pa.
2.4 分析方法 2.4.1 化学分析水样经0.45 μm滤膜过滤后, 通过N-(1-萘基)-乙二胺光度法测定亚硝酸盐浓度.脱氮速率通过线性拟合进行计算, 具体方法为反应前4 h每一小时取样测定亚硝酸盐浓度, 以时间为横轴、以亚硝氮为纵轴拟合直线, 斜率即为最大脱氮速率.通过气相色谱法(SC-3000B, 重庆川仪自动化股份有限公司)测定氮气、甲烷浓度, 并利用理想气体方程和亨利公式计算反应器内总氮气量和总甲烷量.
2.4.2 高通量测序分析取第132 d和第214 d对照组与实验组反应器中污泥作为样本, 利用DNA提取试剂盒抽提样本基因组DNA, 选用细菌通用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)进行PCR扩增.PCR产物经纯化与定量后上机(Illumina Miseq PE250/PE300平台上海派森诺生物科技有限公司).
2.4.3 实时荧光定量PCR分析以提取的基因组DNA为模板, 选用qP1F (GGGCTTGACATCCCA CGAACCTG)和qP1R (CGCCTTCCTCCAGCTTG ACGC)(Ettwig et al., 2009)引物进行定量扩增.扩增程序为:95 ℃预变性3 min, 95 ℃变性15 s, 60 ℃退火30 s, 72 ℃延伸30 s, 进行40个循环, 72 ℃再延伸5 min.每个样品均有3个生物学重复, 基因拷贝数以污泥干重计.
3 结果与讨论(Results and discussion) 3.1 排泥对n-DAMO反应器脱氮效能影响两个SBR反应器的亚硝酸盐浓度如图 2所示.对每天前4 h的亚硝酸盐浓度-时间曲线进行拟合, 发现符合零级动力学方程(表 2, R2> 0.94), 故用前4 h的平均脱氮速率代指最大脱氮速率.第1次排泥前, 1号反应器亚硝酸盐去除量为2.10~5.06 mg · L-1, 2号反应器亚硝酸盐去除量为3.43~4.41 mg · L-1.第1次排泥后, 1号反应器亚硝酸盐去除量先降低为1.69 mg · L-1, 然后恢复到3.24 mg · L-1, 最后达到10.37 mg · L-1; 2号反应器亚硝酸盐去除量最大达到5.02 mg · L-1.第2次排泥后, 1号反应器亚硝酸盐去除量先降低为4.56 mg · L-1, 然后恢复到9.58 mg · L-1, 最后达到12.18 mg · L-1; 2号反应器亚硝酸盐去除量最大达到4.24 mg · L-1.结果表明, 排泥之后1号反应器亚硝酸盐去除量较2号反应器明显提高, 证明排泥可以强化反应器脱氮效能.
 |
| 图 2 亚硝酸盐浓度变化曲线 Fig. 2 Concentration variation of nitrite in two reactors |
| 表 2 反应器排泥前后关键时期脱氮速率 Table 2 Nitrogen removal rate of reactor before and after withdrawing sludge |
两个反应器的脱氮速率如图 3所示.在整个运行过程中, 2号反应器的脱氮速率基本稳定在19 mg · L-1 · d-1.在运行初始阶段, 1号反应器脱氮速率与2号相差不大, 为17.00 mg · L-1 · d-1.第1次排泥后1号反应器脱氮速率逐渐上升至62.21 mg · L-1 · d-1, 为2号反应器的2.23倍.在第2次排泥后, 1号反应器的脱氮速率继续升高, 并在反应末期达到最高值73.10 mg · L-1 · d-1, 是2号反应器的3.41倍.结果表明, 排泥可以提高反应器的脱氮效率.
 |
| 图 3 脱氮速率变化曲线 (黑色虚线箭头表示排泥) Fig. 3 Removal rate of nitrogen in No.1, 2 reactor |
为了进一步探究反应过程, 对两个反应器中的物料进行衡算, 结果如表 3所示.以n-DAMO反应化学计量学为基础, 实测的N2产气速率、甲烷消耗速率均接近于理论值, 表明反应器中发生的是n-DAMO反应, 所以通过排泥可以提高反应器的n-DAMO效能.
| 表 3 CH4消耗、N2产生速率的理论与实际值对比 Table 3 Theory of production rate of N2, consumption of CH4 compared with the actual value |
目前所报道的n-DAMO细菌单独培养脱氮速率及其丰度如表 4所示, 其中, 普遍速率为30~40 mg · L-1 · d-1, 本实验1号反应器在运行到1291 d时脱氮速率达到73.10 mg · L-1 · d-1, 与研究报道的最高速率75.9 mg · L-1 · d-1相差不大, 说明排泥对提高n-DAMO细菌脱氮速率有重要作用.
| 表 4 n-DAMO过程脱氮速率 Table 4 Nitrogen removal rate of n-DAMO process |
两个反应器中不同阶段的微生物群落结构如图 4所示.通过排泥的操作, 1号反应器内微生物群落结构明显优化, n-DAMO细菌隶属的NC10门所占比例均大幅提高, 从38.3%上升到67.7%.变形菌门、绿弯菌门、绿菌门、芽单胞菌门相对丰度全部降低, 表明排泥可以扩大n-DAMO细菌的生长优势.而2号反应器中NC10相对丰度基本维持不变.其他研究表明, 在经过富集且运行良好的n-DAMO反应器中, n-DAMO细菌丰度一般在50%~80%.本研究初始阶段n-DAMO细菌丰度只有38.3%, 较其他研究偏低, 但经过排泥后n-DAMO细菌丰度提高到67.7%, 与其他研究相当.
 |
| 图 4 门水平(a)和属水平(b)微生物群落结构差异比较 (G.C和H1分别为1号反应器实验132 d、214 d测序结果; G.C2和H2分别为2号反应器实验132 d、214 d测序结果) Fig. 4 Comparison of microbial community structure differences at phylum level(a) and genus level(b) |
从属水平上看, 隶属于NC10门的M.oxyfera菌是两个反应器的共同优势微生物.除此之外, 随着排泥的进行大部分在1号反应器中占优的微生物种属, 相对丰度均产生了明显的下降(图 4b).其中, Unclassified_GCA004相对丰度从8.64%下降到2.39%, Unclassified_Rhodocyclaceae相对丰度从4.38%下降到1.43%, Unclassified_Fimbriimonadaceae相对丰度从1.38%下降到0.06%, Methylosinu相对丰度从3.70%下降到0.75%.研究表明, Unclassified_GCA004在微氧条件下有甲烷氧化耦合反硝化的功能(Liu et al., 2013), 通常在红树林沉积物(Wu et al., 2016b)和淡水湖泊沉积物(Wan et al., 2017)中检测到, 在这两种生境下NO3--N和NO2--N浓度分别为0~0.329 mg · L-1、0.063~0.16 mg · L-1(Fernandes et al., 2010); 而在本实验中NO2--N浓度约为30~40 mg · L-1, 且不存在硝酸盐, 适宜n-DAMO细菌的生长(15~50 mg · L-1), 不适宜Unclassified_GCA004的生长, 所以通过排泥可以强化n-DAMO细菌的优势.Yoshida等(2012)在0.3 mg · g-1的稻田土中发现Unclassified_Rhodocyclaceae具有反硝化的能力, 基质利用速率约为0.09 mmol · g-1 · h-1, 而n-DAMO细菌的基质利用速率为0.28 mmol · g-1 · h-1(Allegue et al., 2018), 较Unclassified_Rhodocyclaceae更快, 所以反应器的环境更适宜n-DAMO细菌, 且排泥后n-DAMO细菌基质利用速率更高, 更容易成为优势物种.Methylosinus是一种以甲烷为碳源的甲烷氧化细菌, 研究报道, Methylosinus的甲烷消耗速率为0.95 mg · L-1 · h-1(Ordaz et al., 2014), 而M.oxyfera菌甲烷消耗速率为1.36 mg · L-1 · h-1(Allegue et al., 2018), 说明n-DAMO细菌利用甲烷的能力更强, 排泥使所有微生物同比减少的情况下, n-DAMO细菌凭借高的基质利用速率占有优势.综上所述, 经过排泥后, M.oxyfera菌以其更适合反应器环境与更高的基质利用速率, 使得非目标微生物相对丰度都有所降低, 而M.oxyfera菌丰度增高, 最终达到强化n-DAMO过程的目的.
 |
| 图 5 1号反应器132 d、214 d微生物群落结构差异性分析 Fig. 5 Difference analysis of microbial community structure of sludge in No.1 reactor on 132 d and 214 d |
反应器中n-DAMO细菌数量如图 6所示.经过排泥, 1号反应器内功能微生物数量大幅提升.qPCR结果表明, 1号反应器132 d n-DAMO细菌的量为1.404×108 copies · g-1, 到214 d增长为4.854×108 copies · g-1, 提高了2.46倍; 2号反应器132 d n-DAMO细菌的量为2.490×108 copies · g-1, 到214 d增长为3.291×108 copies · g-1, 仅增长了32%.在He等(2015b)的研究中, n-DAMO细菌量为(5.0×108±0.4×108) copies · g-1, 与本研究基本一致; 而在Zhou等(2015)的研究中, n-DAMO细菌量为(0.85×108~1.0×108) copies · g-1, 本研究是其1~5倍.实验结果表明, 排泥确实可以直接提高n-DAMO细菌的数量.
 |
| 图 6 反应器中n-DAMO菌绝对数量比较 Fig. 6 Comparison of absolute abundance of n-DAMO bacrteria in the reactors |
本文以NO2-为电子受体成功富集n-DAMO功能微生物, 通过排泥对n-DAMO过程进行强化.132 d时1号反应器中n-DAMO细菌数量仅为2号反应器的56.39%, 但其脱氮速率是2号反应器的2.23倍, 表明1号反应器中n-DAMO细菌的活性是2号反应器的3.95倍, 214 d时1号反应器中n-DAMO微生物数量是2号反应器的1.47倍, 脱氮速率达到了2号反应器的3.41倍, 活性提高了2.31倍.1号反应器n-DAMO细菌数量和活性都高于2号反应器, 可能是由于排泥使得n-DAMO细菌丰度提高.1号反应器中n-DAMO细菌丰度所占比例是2号反应器的1.77倍, 达到了67.7%, 与目前已报道的最高丰度相当, 较高的丰度有利于n-DAMO细菌发挥优势作用.
4 结论(Conclusions)1) 排泥能够提高SBR中n-DAMO过程的脱氮速率, 脱氮速率从初期的17.00 mg · L-1 · d-1最高达到73.10 mg · L-1 · d-1, 可满足工程应用需求, 具备开发潜力.
2) 排泥可以选择性富集n-DAMO菌M.oxyfera, 强化后M.oxyfera相对丰度由38.3%上升到67.7%.n-DAMO菌数量由1.404×108 copies · g-1增长到4.854×108 copies · g-1.
3) 排泥后非目标微生物Unclassified_GCA004、Unclassified_Rhodocyclaceae、Unclassified_Fimbriimonadaceae和Methylosinus相对丰度均明显降低, M.oxyfera具有更高的基质利用速度与生长速度可能是其内在原因.
Allegue T, Arias A, Fernandez-Gonzalez N, et al. 2018. Enrichment of nitrite-dependent anaerobic methane oxidizing bacteria in a membrane bioreactor[J]. Chemical Engineering Journal, 347: 721–730.
DOI:10.1016/j.cej.2018.04.134
|
柴风光, 卢培利, 李微薇, 等. 2018. 利用硝酸盐和亚硝酸盐同步富集厌氧甲烷氧化微生物的比较实验[J]. 微生物学通报, 2018, 45(4): 762–770.
|
Chen J, Zhou Z C, Gu J D. 2014. Occurrence and diversity of nitrite-dependent anaerobic methane oxidation bacteria in the sediments of the South China Sea revealed by amplification of both 16S rRNA and pmoA genes[J]. Applied Microbiology and Biotechnology, 98(12): 5685–5696.
DOI:10.1007/s00253-014-5733-4
|
Ettwig K F, Shima S, Pas-Schoonen K T V D, et al. 2008. Denitrifying bacteria anaerobically oxidize methane in the absence of Archaea[J]. Environmental Microbiology, 10(11): 3164–3173.
DOI:10.1111/emi.2008.10.issue-11
|
Ettwig K F, Van A T, Kt P S, et al. 2009. Enrichment and molecular detection of denitrifying methanotrophic bacteria of the NC10 phylum[J]. Applied and Environmental Microbiology, 75(11): 3656–3662.
DOI:10.1128/AEM.00067-09
|
Fernandes S O, Lokabharathi P A, Bonin P C, et al. 2010. Denitrification:an important pathway for nitrous oxide production in tropical mangrove sediments (Goa, India)[J]. Journal of Environmental Quality, 39(4): 1507–1516.
DOI:10.2134/jeq2009.0477
|
Fu L, Ding J, Lu Y Z, et al. 2017. Nitrogen source effects on the denitrifying anaerobic methane oxidation culture and anaerobic ammonium oxidation bacteria enrichment process[J]. Applied Microbiology and Biotechnology, 101(9): 3895–3906.
DOI:10.1007/s00253-017-8163-2
|
Han P, Gu J D. 2013. A newly designed degenerate PCR primer based on pmoA, gene for detection of nitrite-dependent anaerobic methane-oxidizing bacteria from different ecological niches[J]. Applied Microbiology and Biotechnology, 97(23): 10155–10162.
DOI:10.1007/s00253-013-5260-8
|
He Z, Geng S, Shen L, et al. 2015a. The short-and long-term effects of environmental conditions on anaerobic methane oxidation coupled to nitrite reduction[J]. Water Research, 68(47): 554–562.
|
He Z, Cai C, Shen L, et al. 2015b. Effect of inoculum sources on the enrichment of nitrite-dependent anaerobic methane-oxidizing bacteria[J]. Applied Microbiology and Biotechnology, 99(2): 939–946.
|
He Z, Geng S, Wang L, et al. 2016. Improvement of mineral nutrient concentrations and pH control for the nitrite-dependent anaerobic methane oxidation process[J]. Separation and Purification Technology, 162(3): 148–153.
|
Hu B L, Shen L D, Huang Q, et al. 2014a. Evidence for nitrite-dependent anaerobic methane oxidation as a previously overlooked microbial methane sink in wetlands[J]. Proceedings of the National Academy of Sciences of the United States of America, 111(12): 4495–4500.
DOI:10.1073/pnas.1318393111
|
Hu B, He Z, Geng S, et al. 2014b. Cultivation of nitrite-dependent anaerobic methane-oxidizing bacteria:impact of reactor configuration[J]. Applied Microbiology and Biotechnology, 98(18): 7983–7991.
DOI:10.1007/s00253-014-5835-z
|
Hu S, Zeng R J, Burow L C, et al. 2009. Enrichment of denitrifying anaerobic methane oxidizing microorganisms[J]. Environmental Microbiology Reports, 1(5): 377–384.
DOI:10.1111/j.1758-2229.2009.00083.x
|
Kampman C, Hendrickx T L G, Luesken F A, et al. 2012. Enrichment of denitrifying methanotrophic bacteria for application after direct low-temperature anaerobic sewage treatment[J]. Journal of Hazardous Materials, 227: 164–171.
|
Kampman C, Temmink H, Hendrickx T L G, et al. 2014. Enrichment of denitrifying methanotrophic bacteria from municipal wastewater sludge in a membrane bioreactor at 20℃[J]. Journal of Hazardous Materials, 274: 428–435.
DOI:10.1016/j.jhazmat.2014.04.031
|
Li W, Lu P, Chai F, et al. 2018. Long-term nitrate removal through methane-dependent denitrification microorganisms in sequencing batch reactors fed with only nitrate and methane[J]. Amb Express, 8(1): 108.
DOI:10.1186/s13568-018-0637-9
|
Lieberman R L, Rosenzweig A C. 2004. Biological methane oxidation:Regulation, biochemistry, and active site structure of particulate methane monooxygenase[J]. Critical Reviews in Biochemistry and Molecular Biology, 39(3): 147–164.
DOI:10.1080/10409230490475507
|
Liu J, Sun F, Wang L, et al. 2013. Molecular characterization of a microbial consortium involved in methane oxidation coupled to denitrification under micro-aerobic conditions[J]. Microbial Biotechnology, 7(1): 64–76.
|
Lu P, Liu T, Ni B, et al. 2019. Growth kinetics of Candidatus 'Methanoperedens nitroreducens' enriched in a laboratory reactor[J]. Science of the Total Environment, 695: 442–450.
|
Lu Y Z, Fu L, Ding J, et al. 2016. Cr(Ⅵ) reduction coupled with anaerobic oxidation of methane in a laboratory reactor[J]. Water Research, 102: 445–452.
DOI:10.1016/j.watres.2016.06.065
|
Luesken F A, Jaime Sánchez, Alen T A, et al. 2011. Simultaneous nitrite-dependent anaerobic methane and ammonium oxidation processes[J]. Applied and Environmental Microbiology, 77(19): 6802–6807.
DOI:10.1128/AEM.05539-11
|
Ordaz A, López J C, Figueroa-González I, et al. 2014. Assessment of methane biodegradation kinetics in two-phase partitioning bioreactors by pulse respirometry[J]. Water Research, 67: 46–54.
DOI:10.1016/j.watres.2014.08.054
|
Raghoebarsing A A, Pol A, Pas-Schoonen K T V D, et al. 2006. A microbial consortium couples anaerobic methane oxidation to denitrification[J]. Nature, 440(7086): 918–921.
DOI:10.1038/nature04617
|
沈李东, 胡宝兰, 郑平. 2011. 甲烷厌氧氧化微生物的研究进展[J]. 土壤学报, 2011, 48(3): 619–628.
|
Solomon S, Qin M, Manning Z, et al. 2007. Climate Change 2007:The Physical Science Basis[J]. Cambridge University Press: 1–21.
|
Wan Y, Ruan X, Zhang Y, et al. 2017. Illumina sequencing-based analysis of sediment bacteria community in different trophic status freshwater lakes[J]. Microbiology Open, 6: e450.
DOI:10.1002/mbo3.450
|
Wang R, Han X, Lu P, et al. 2016. Co-occurrence and diversity of nitrite-dependent anaerobic methane oxidizing and anaerobic ammonium oxidizing bacteria in the water-level fluctuation zone of the three gorges reservoir, China[J]. Fresenius Environmental Bulletin, 25(12): 5456–5466.
|
Wu Q, Chang J, Yan X, et al. 2016a. Electrical stimulation enhanced denitrification of nitrite-dependent anaerobic methane-oxidizing bacteria[J]. Biochemical Engineering Journal, 106: 125–128.
DOI:10.1016/j.bej.2015.11.014
|
Wu P, Xiong X, Xu Z, et al. 2016b. Bacterial communities in the rhizospheres of three mangrove tree species from Beilun Estuary, China[J]. Plos One, 11(10): e0164082.
DOI:10.1371/journal.pone.0164082
|
Yoshida M, Ishii S, Fujii D, et al. 2012. Identification of active denitrifiers in rice paddy soil by DNA-and RNA-based analyses[J]. Microbes & Environments, 27(4): 456–461.
|
Yue X, Yu G, Liu Z, et al. 2018. Start-up of the completely autotrophic nitrogen removal over nitrite process with a submerged aerated biological filter and the effect of inorganic carbon on nitrogen removal and microbial activity[J]. Bioresource Technology, 254: 347–352.
DOI:10.1016/j.biortech.2018.01.107
|
赵荣, 朱雷, 吴箐, 等. 2017. 亚硝酸盐型甲烷厌氧氧化过程影响因素研究[J]. 环境科学学报, 2017, 37(1): 178–184.
|
Zhou L, Wang Y, Long X E, et al. 2014. High abundance and diversity of nitrite-dependent anaerobic methane-oxidizing bacteria in a paddy field profile[J]. FEMS Microbiology Letters, 360(1): 33–41.
DOI:10.1111/fml.2014.360.issue-1
|
Zhu B, Van G D, Fritz C, et al. 2012. Anaerobic oxidization of methane in a minerotrophic peatland:enrichment of nitrite-dependent methane-oxidizing bacteria[J]. Applied and Environmental Microbiology, 78(24): 8657–8665.
DOI:10.1128/AEM.02102-12
|