 2017, Vol. 37
2017, Vol. 37



随着我国工农业和城镇化的快速发展,大气、水和土壤受到的重金属污染日益严重.大气中重金属污染物主要附着在气溶胶或颗粒物表面,可以远距离传输并通过干湿沉降的形式进入地表生态系统(林静等,2016).河流中的重金属污染物与水中悬浮物或沉积物发生吸附、络合或共沉淀作用,最终在河流沉积物中富集(张伯镇等,2015).可见释放到环境中的大部分污染物质最终都要进入土壤,土壤重金属的修复成为环境修复的核心问题.目前我国土壤重金属污染严重,受Cd、Pb、As、Cu、Zn等重金属污染的耕地约2000万hm2,每年因重金属污染的粮食(如“Cd米”)达1000多万t,造成直接经济损失超过200亿元(樊霆等,2013),因此土壤重金属污染的监测、评估和修复技术措施的研究和应用显得尤为迫切.
植物修复技术(Phytoremediation)具有成本低廉、美化环境等优点,表现出环境修复的巨大潜力.以杨柳科为代表的速生木本植物具有生长迅速、生物量大、根系发达、生长周期长的优点,能耐受一定程度的重金属污染(Marmiroli et al.,2011),并能积累较大总量的重金属,近年来其环境修复的潜力受到了较多的关注.另一方面,丛枝菌根是生态系统中常见的互利共生现象,在植物的营养循环和逆境耐受中起着重要作用,丛枝菌根真菌与植物共生的联合方式在重金属污染土壤的修复中显示出极大的应用潜力(王发园和林先贵,2007).许多研究表明无论在自然生长状况还是人工接种的条件下,许多丛枝菌种都能与杨树良好的互利共生,能够提高杨树的存活、生长和抗逆性(Beauchamp et al.,2006;Gehring et al.,2006),但丛枝菌根真菌对重金属污染物积累的效应随着菌种、杨树种类(品种或基因型)的不同而表现出一定的差异(Lingua et al.,2008;Chen et al.,2015).然而,很少有研究关注丛枝菌根对复合污染条件下不同性别杨树重金属积累能力和分配特征的影响.
美洲黑杨(Populus deltoides)具有适应性强、生长快、材质优良、抗逆性强等优点,一些优良美洲黑杨品系(既有雄株品系又有雌株品系)已经广泛种植于黄淮平原和长江中下游地区,在当地的林业生产和生态建设中发挥着巨大作用.已有的研究表明,美洲黑杨根内的寄生真菌多样性和群落结构会因土壤类型和土壤层次的不同而表现出差异(Karliński et al.,2010; Gottel et al.,2011),菌根侵染状况会受到干旱、水淹、重金属等环境胁迫的影响(Karliński et al.,2010; Yang et al.,2016),同时也会受到外生菌根竞争效应的作用(Lodge,1989).另一方面,美洲黑杨是典型的雌雄异株植物,前期研究表明,重金属对雌雄杨树生长状况和生理适应特征的影响程度依赖于重金属的种类及其浓度效应,低浓度条件下雌株对重金属的积累能力较强,但雄株对高浓度重金属胁迫具有较强的耐受性(Chen et al.,2013;Chen et al.,2014 ;Chen et al.,2015).这些研究均针对单一重金属污染开展研究,没有关注复合污染条件下雌雄杨树重金属积累和分配的差异.因此,本研究以雌雄美洲黑杨为对象,关注复合污染条件下雌雄杨树体内重金属积累与分配的差异,关注接种丛枝菌根真菌对雌雄杨树重金属吸收和分配的影响,研究结果可以为复合污染土壤的环境修复,以及利用丛枝菌根真菌-速生木本植物的联合方式修复重金属污染土壤提供理论依据.
2 材料与方法(Materials and methods) 2.1 实验材料和试验设计2014年4月,于江苏省镇江市采集一年生健康的美洲黑杨雌雄枝条,这些枝条为P. deltoides Bartr. cv. ‘Lux’(I-69/55)× P. deltoides Bartr. cv. ‘Harvard’(I-63/51)的F1代,母树树龄约30 a,根据开花结实性状辨别雌雄性别.所有枝条经0.5% NaClO(w/v)浸泡消毒30 min,去离子水冲洗数次后于Hoagland营养液中水培.选择长势一致的雌雄枝条用于菌种侵染处理.
砂培基质由粗沙、蛭石和珍珠岩组成,以体积比3∶1∶1的比例充分混匀后于0.11 MPa,121 ℃灭菌2 h.采用塑料花盆进行栽培,每盆盛装灭菌基质12 kg.丛枝菌根真菌接种物Glomus intraradices(BGC AH01)来自于“丛枝菌根真菌种质资源库”,购于北京市农林科学院植物营养与资源研究所.菌种分离自重金属污染区先锋草本植物(双穗雀稗和狗牙根)的根际土壤,通过单孢培养扩繁到高梁的根系,接种物是含有孢子、侵染根段、外生菌丝和培养基质的混合繁殖物.水培幼苗根长约10 cm时移栽幼苗,每盆移栽一株,移栽时将20 g接种物(每g接种物中约含50个孢子)接种于雌雄美洲黑杨根际,未接种处理则将灭菌后的接种物以相同方式处理.为了避免养分的缺乏,每周定期向每株美洲黑杨幼苗浇100~200 mL Hoagland营养液.
本研究为性别(雌、雄)、Pb(未污染、200 mg·kg-1)、Cd(未污染、10 mg·kg-1)、菌根真菌(未接种、接种)等4因素2水平的完全随机试验设计,共16个处理.根据我国GB15618-1995《土壤环境质量标准》,本研究所设置的污染梯度为中等程度的污染水平,在我们野外土壤环境调查时,污染严重地区能够达到本研究的污染水平.接种丛枝菌根真菌1个月以后,进行重金属污染处理,每个性别含以下处理:①对照,未接种+未进行重金属污染处理;②未接种+Pb污染;③未接种+Cd污染;④未接种+Pb/Cd复合污染;⑤接种丛枝菌根真菌+未进行重金属污染处理;⑥接种+Pb污染;⑦接种+Cd污染;⑧接种+Pb/Cd复合污染.对于污染处理,分别将200 μmol·L-1 CdCl2·2.5H2O或2 mmol·L-1的Pb(NO3)2均匀添加到相应处理中,污染物分10次添加,每次添加含重金属溶液约500 mL,间隔2 d添加1次,最终达到10 mg Cd每kg基质或200 mg Pb每kg基质的污染水平.每一处理5次重复,每重复包含5株幼苗.实验在仅遮蔽雨水的大棚中进行,气温过高时利用75%遮阳网遮蔽降温,每天对所有供试植株进行统一浇水(蒸馏水)、除草防虫和防病等管理.实验处理期间,白天温度为18~33 ℃,夜间温度为12~17 ℃,相对湿度为45%~87%.实验处理持续一个生长季,于2014年6月15开始,2014年9月30日结束.
2.2 苗木收获和分析测试实验处理结束时,每个处理随机选择3株幼苗测定菌根侵染率,每1株幼苗选取至少100根鲜活根段(直径<1 mm)测定菌根真菌侵染率,测定方法参照McGonigle等(1990),侵染率表示为含有侵染特征(丛枝结构、囊泡和菌丝)的百分率.
每个处理随机选择5株幼苗,分别收获幼苗的叶片、茎、粗根(直径>1 mm)和细根(直径<1 mm),将叶片和茎于自来水冲洗后用去离子水清洗,将粗根和细根于20 mmol·L-1的EDTA-Na中浸泡30 min,再用去离子水清洗.将所有植物器官烘干至衡重后称重,计算总生物量.将植物样品干样粉碎过筛(100目),细粉用于Pb和Cd含量测定.经HNO3-HClO4(VHNO3∶VHClO4=5∶1)消解后,采用火焰原子吸收法测定(岛津AA-7000火焰原子吸收光谱仪)各器官的Pb和Cd含量(用占干物质的质量分数计),各器官Pb和Cd的积累量为重金属含量与对应干重的乘积.
采用迁移率(translocation factor,TF)、滞留率(retention rate,RR)、富集系数(bioconcentration factor,BCF)衡量雌雄美洲黑杨对Pb和Cd的转移、分配和富集能力.地上部重金属含量、地下部重金属含量、土壤中重金属含量分别用MCA(metal concentration in above-ground organs)、MCB(metal concentration in below-ground organs)、MCS(metal concentration in soils)表示.计算公式如下:
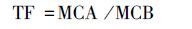
|
(1) |
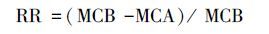
|
(2) |
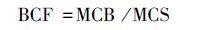
|
(3) |
采用SPSS 16.0统计软件对生物量、菌根真菌侵染率、各器官重金属含量及积累量、重金属富集与分配参数进行方差分析,用Tukey多重比较法检验处理间的差异显著性(p<0.05).用配对样本T检验(Paired Sample T test)检验某一参数因接种丛枝菌根真菌引起的差异显著性.采用多元方差分析(MANOVA)检验性别、铅、镉、菌根及其交互作用对所有参数影响的显著性.
3 结果与分析(Results and analysis) 3.1 各种因子及其交互作用对所有参数影响的显著性检验多元方差分析表明(表 1),所有参数综合地受到性别、Pb、Cd、菌根及其交互效应的极显著影响(p<0.01).
| 表 1 多元方差分析检验性别、铅、镉、菌根及其交互效应对所有参数的综合影响 Table 1 Multivariate analysis of variance(MANOVA)in identifying the single and interactive effects of sex,Pb,Cd and AM on all parameters |
如图 1a所示,与对照相比,所有处理并没有显著影响雌雄美洲黑杨的生物量.对照条件下,接种菌根真菌并没有显著影响雌雄美洲黑杨的生物量,在各种污染条件下,接种菌根真菌均不同程度增加了雌雄美洲黑杨的生物量,但未达显著水平.相应条件下,雌雄美洲黑杨的生物量没有显著的性别差异.
 |
| 图 1 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨生物量(a)和侵染率(b)的影响(注:图柱上标不含相同字母表示处理间差异显著(p<0.05)) Fig. 1 Effects of AMF inoculation on biomass(a)and mycorrhizal colonization rate(b)in females and males of Populus deltoides when exposed to HMs stress |
如图 1b所示,未接种丛枝菌根真菌条件下,没有检测出美洲黑杨根系含有菌根真菌侵染的特征.接种丛枝菌根真菌的所有处理侵染率均大于30%,对应条件下雌雄间的侵染率没有显著的性别差异.与接种丛枝菌根真菌的对照相比,Cd污染条件下雄株的侵染率显著增加.复合污染条件下雌株的侵染率显著低于Cd污染条件下的侵染率.
3.3 接种丛枝菌根真菌对雌雄美洲黑杨各器官Pb含量的影响如表 2所示,未接种丛枝菌根真菌条件下,与对照相比,Pb污染和复合污染均显著增加了美洲黑杨雄株叶片、粗根和细根Pb含量.与Pb污染相比,复合污染条件下雄株的叶Pb和粗根Pb含量分别增加232%和19%.另一方面,Pb污染和复合污染均显著增加了雌株叶片和细根Pb含量,复合污染还显著增加了雌株粗根Pb含量.与Pb污染相比,复合污染条件下雌株的叶Pb含量增加了45%,细根Pb含量却下降了26%.
| 表 2 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨各器官Pb含量的影响 Table 2 Effects of AMF inoculation on Pb content in organs of females and males of Populus deltoides when exposed to HMs stress |
Pb污染条件下,接种丛枝菌根真菌均显著增加了2种性别植株叶片、粗根和细根Pb含量,雄株叶片、粗根和细根Pb含量分别增加了75%、40%和104%,雌株这些器官Pb含量分别增加48%、159%和88%.复合污染条件下,接种丛枝菌根真菌也显著增加了雌株叶片、粗根和细根的Pb含量,这些器官Pb含量分别增加19%、172%和232%;然而,接种丛枝菌根真菌仅显著增加了雄株的细根Pb含量,增加幅度为142%,却显著降低了叶片和粗根Pb含量,下降幅度分别为39%和17%.
Pb污染条件下,与未接种雌株相比,对应雄株具有较高的粗根Pb含量;与接种雌株相比,对应雄株具有较低的粗根Pb含量,较高的细根Pb含量.复合污染条件下,与未接种雌株相比,对应雄株具有较高的叶Pb、粗根Pb和细根Pb含量;与接种雌株相比,对应雄株具有较低的粗根Pb和细根Pb含量.
3.4 接种丛枝菌根真菌对雌雄美洲黑杨幼苗各器官Pb积累量及总Pb积累量的影响如表 3所示,未接种丛枝菌根真菌条件下,与对照相比,Pb污染和复合污染均显著增加了雄株叶片、细根Pb积累量和总Pb积累量,Pb污染还显著增加了雄株粗根Pb积累量.与Pb污染相比,复合污染条件下的雄株叶Pb积累量增加158%.与对照相比,Pb污染和复合污染均显著增加了雌株叶片、细根Pb积累量和总Pb积累量,Pb污染还显著增加了雌株茎Pb积累量.与Pb污染相比,复合污染条件下雌株茎Pb积累量和细根Pb积累量分别下降29%和45%.
| 表 3 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨各器官Pb积累量的影响 Table 3 Effects of AMF inoculation on Pb accumulation amount in organs of females and males of Populus deltoides when exposed to HMs stress |
Pb污染条件下,接种丛枝菌根真菌显著增加了雄株的叶片、粗根、细根Pb积累量及总Pb积累量,增加幅度分别为112%、123%、177%、118%;复合污染条件下,接种丛枝菌根真菌显著增加了雄株细根Pb积累量和总Pb积累量,增加幅度为197%和43%.Pb污染和复合污染条件下,接种丛枝菌根真菌均显著促进了雌株叶片、粗根、细根Pb积累量及总Pb积累量,Pb污染下增加幅度分别为67%、251%、70%和66%,复合污染条件下增加幅度分别为43%、418%、251%和134%.
Pb污染条件下,与未接种雌株相比,对应雄株具有较低的茎和细根Pb积累量;与接种雌株相比,对应雄株具有较高的细根Pb积累量.复合污染条件下,与未接种雌株相比,对应雄株具有较高的叶Pb积累量;然而,与接种雌株相比,对应雄株具有较低的粗根、细根Pb积累量及总Pb积累量.
3.5 接种丛枝菌根真菌对雌雄黑杨幼苗各器官Cd含量的影响对照及Pb污染条件下,2种性别美洲黑杨各器官中均未检测出Cd.如表 4所示,未接种丛枝菌根真菌条件下,与Cd污染相比,复合污染条件下雄株叶片、茎、粗根和细根Cd含量分别增加224%、89%、99%和68%,雌株茎、粗根和细根Cd含量却分别降低53%、55%和27%.
| 表 4 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨各器官Cd含量的影响 Table 4 Effects of AMF inoculation on Cd content in organs of females and males of Populus deltoides when exposed to HMs stress |
Cd污染条件下,接种丛枝菌根真菌引起雄株叶片、茎和细根Cd含量分别增加71%、54%和279%;接种丛枝菌根真菌引起雌株细根Cd含量增加了25%,叶片和茎Cd含量却分别下降了57%和42%.复合污染条件下,接种丛枝菌根真菌引起雄株叶片、茎和粗根Cd含量降低了69%、43%和41%,细根Cd含量却增加了31%;接种丛枝菌根真菌引起雌株叶片Cd含量下降32%,茎、粗根和细根Cd含量却显著增加了39%、322%和62%.
Cd污染条件下,未接种雌株具有比对应雄株更高的茎、粗根和细根Cd含量,然而,接种雌株具有比对应雄株更低的叶和细根Cd含量.复合污染条件下,未接种雄株所有器官的Cd含量均显著高于对应雌株,然而,接种雄株仅叶片和细根Cd含量显著高于对应雌株,而雄株粗根的Cd含量却低于雌株.
3.6 接种丛枝菌根真菌对雌雄美洲黑杨幼苗各器官Cd积累量及总Cd积累量的影响如表 5所示,未接种丛枝菌根真菌条件下,与Cd污染相比,复合污染条件下雄株叶片Cd积累量和总Cd积累量分别增加297%和112%,而雌株各器官Cd积累量及总积累量没有显著变化.
| 表 5 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨各器官Cd积累量的影响 Table 5 Effects of AMF inoculation on Cd accumulation amount in organs of females and males of Populus deltoides when exposed to HMs stress |
Cd污染条件下,接种丛枝菌根真菌引起雄株叶片、茎、粗根、细根Cd积累量及总积累量分别增加85%、53%、81%、236%和160%,但接种丛枝菌根真菌仅引起雌株细根的Cd积累量增加59%,对其其他器官Cd积累量及总Cd积累量无显著影响.复合污染条件下,接种丛枝菌根真菌引起雄株叶片Cd积累量下降57%,细根Cd积累量却增加60%,对其他器官Cd积累量及总积累量无显著影响;接种丛枝菌根真菌引起雌株茎、粗根、细根Cd积累量及总Cd积累量分别增加52%、691%、71%和70%.
Cd污染条件下,与接种丛枝菌根真菌的雌株相比,对应雄株具有较高的叶片、细根Cd积累量和总Cd积累量.复合污染条件下,与未接种丛枝菌根真菌的雌株相比,对应雄株具有较高的叶Cd积累量和总Cd积累量.
3.7 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨Pb富集特征的影响如表 6所示,与对照相比,未接种丛枝菌根真菌条件下,Pb污染条件下雄株Pb迁移率下降44%,复合污染条件下却增加32%;Pb污染和复合污染条件下雌株Pb迁移率均显著下降,下降幅度分别是57%和35%.值得注意的是,与Pb污染相比,复合污染条件下雄株和雌株的Pb迁移率均显著增加,增加幅度分别为135%和52%.Pb污染条件下,接种丛枝菌根真菌一定程度下降低了2种性别的Pb迁移率,但均未达显著水平.复合污染条件下,接种丛枝菌根真菌引起雄株和雌株Pb迁移率分别下降65%和59%.Pb污染条件下,不管接种丛枝菌根真菌与否,性别间的Pb迁移率均无显著差异.复合污染条件下,与未接种雌株相比,对应雄株具有更高的Pb迁移率.
| 表 6 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨Pb/Cd富集特征的影响 Table 6 Effects of AMF inoculation on allocation pattern of Pb and Cd in females and males of Populus deltoides when exposed to HMs stress |
与对照相比,Pb污染和复合污染条件下,不管接种丛枝菌根真菌与否,2种性别的Pb富集系数均显著下降.与Pb污染相比,复合污染条件下的雌株或雄株Pb富集系数没有显著变化.Pb污染和复合污染条件下,接种丛枝菌根真菌均显著增加了2种性别的Pb富集系数.Pb污染和复合污染的对应条件下,2种性别植株的Pb富集系数均没有显著差异.
3.8 接种丛枝菌根真菌对受重金属胁迫的雌雄美洲黑杨Cd富集特征的影响如表 6所示,未接种丛枝菌根真菌条件下,与Cd污染相比,复合污染条件下雄株Cd迁移率增加78%,雌株的Cd迁移率没有显著变化.Cd污染和复合污染条件下,接种丛枝菌根真菌均显著降低了2个性别的Cd迁移率,雄株分别下降37%和69%,雌株则分别下降64%和53%.各种对应条件下,雄株的Cd迁移率均显著大于雌株.
未接种丛枝菌根真菌条件下,与Cd污染相比,复合污染条件下雄株Cd富集系数显著增加86%,而雌株却下降20%.2种污染条件下,接种丛枝菌根真菌均不同程度地增加了2种性别的Cd富集系数.Cd污染条件下,与未接种雌株相比,对应雄株具有较低的Cd富集系数,接种丛枝菌根真菌条件下性别间的差异相反;复合污染条件下,与未接种雌株相比,雄株具有较高的Cd富集系数,接种条件下两性别的Cd富集系数无显著差异.
4 讨论(Discussion)重金属复合污染不同于单一污染的环境效应,对于重金属的吸收、迁移和富集特征常常表现出协同促进或拮抗的作用.例如,曹福亮等(2012)发现,与单一污染相比,Pb/Cd复合污染降低了银杏各器官的Cd含量,却不同程度增加了各器官的Pb含量;张永超等(2011)发现,复合污染条件下,Cd的添加对白榆叶、茎Pb含量具有促进作用,Pb的添加对白榆所有器官Cd含量具有拮抗作用;Wang等(2016)发现,低浓度Cd和Zn污染条件下,2种重金属表现出拮抗的作用,而在高浓度条件下则表现出协同吸收的作用;Yu等(2015)发现,与单一污染相比,复合污染条件下龙葵根系的Pb和Cd含量都表现出协同促进作用.这些研究表明,复合污染引起协同吸收或者拮抗作用的发生会因植物种类和重金属浓度的不同而表现出差异,复合污染的环境效应显得非常复杂.本研究中,复合污染引起了美洲黑杨雄株叶片和根系Pb和Cd含量均不同程度的增加,叶片的增加幅度最大;然而,复合污染虽然不同程度地增加了雌株叶片的Pb或Cd含量,却不同程度地降低了粗根和细根的Pb或Cd含量.可见美洲黑杨雄株在Pb/Cd复合污染条件下,对Pb/Cd 2种离子的吸收和积累发生了协同促进作用,这可能与复合污染条件下,不同重金属离子对培养基质颗粒表面存在的竞争吸附有关,即一种离子的存在会对共存的其他离子的吸附产生抑制作用,从而增加土壤中重金属的生物有效浓度(林青和徐绍辉,2008).然而,对Pb/Cd的协同吸收作用并没有发生在雌株中,推测复合污染可能引起雌株根系受到较大伤害,根系受损导致的活力下降会导致根系对重金属离子或其它矿质元素的吸收能力下降.另一方面,与单一污染相比,复合污染条件下2种性别的Pb或Cd的迁移率显著增加,而Pb或Cd在它们根系的滞留率显著降低,与孙建伶和罗立强(2014)及曹福亮等(2012)的研究结果相似.复合污染条件下,较多的Pb或Cd向美洲黑杨叶片的转移将因叶片脱落而归还到土壤中,给复合污染土壤修复造成更大的困难.
利用植物与耐性丛枝菌根联合修复受重金属污染土壤是一种重要的修复方式.本研究中,我们发现接种丛枝菌根真菌会影响重金属在植物器官的积累特征,Pb污染和Pb/Cd复合污染条件下,接种丛枝菌根真菌显著增加了雌雄美洲黑杨叶片、粗根和细根的Pb含量(复合污染条件下雄株叶片和粗根除外);接种丛枝菌根真菌均显著增加了雌雄美洲黑杨细根的Cd含量,对其它器官Cd含量的影响没有明显的规律,因性别、器官、污染条件不同而表现出差异.接种丛枝菌根真菌引起植物重金属积累的增加,一方面可能是因为丛枝菌根活化了根际圈的重金属(张旭红等,2012),能够提高土壤重金属的生物有效性(Liu et al.,2015),另一方面丛枝菌根增加了根系的吸收面积(刘晓娜等,2011),增加了对重金属的吸收.其它学者也发现了类似的结果,例如,李霞等(2014)发现,接种G. intraradices能增加翅荚木地上和地下部的Pb积累量;Chen等(2015)发现,接种G. mosseae不同程度地促进了雌雄青杨对Pb的积累,对雌株的促进效应更显著;Ali等(2015)发现,接种G. intraradices显著增加了各Cd污染梯度条件下亚麻的Cd含量.菌根不但会影响植物根系对重金属的吸收,还会影响重金属在植物器官中的转移和分配.例如,黄晶等(2012)发现,接种G. intraradices降低了紫花苜蓿地上部Cd含量,却显著增加了地下部Cd含量,从而减弱了重金属向地上部的转运能力.廖妤婕(2014)发现G. mosseae虽然减少了Pb和Cd的吸收量,但能够增强桉树根部对重金属的滞留作用、细胞壁固持作用和可溶组分区隔化作用(液泡),促进重金属向活性更低的形态转化.菌根增强植物根系对重金属的滞留作用与菌根内含大量半胱氨酸的蛋白配体有关,这类配体能与重金属形成复合物而降低重金属的迁移能力(王发园和林先贵,2007);也可能与重金属大量沉积于菌丝细胞壁和液泡有关(Xu et al.,2016),重金属在菌丝细胞的积累可以缓冲重金属对植物根系的负面影响.本研究中,不管是单一污染还是复合污染条件下,接种丛枝菌根真菌均能显著增加雌雄美洲黑杨Pb和Cd的富集系数,并能降低Pb和Cd向雌雄美洲黑杨地上部分的迁移率,增加Pb和Cd在雌雄美洲黑杨根系的滞留率,与以上研究结果相似.菌根的这种效应对于利用多年生木本植物修复土壤显得尤其重要,一方面菌根能增加雌雄美洲黑杨的重金属积累量,另一方面能将更多的重金属积累在根系中,既有利于污染土壤的环境修复,也能减少因落叶等原因导致重金属向土壤的归还.
重金属积累总量是衡量植物积累重金属能力的重要指标,前期研究表明,杨树不同物种(He et al.,2013)、基因型(Baldantoni et al.,2014)、种群(Lei et al.,2007)积累重金属的能力存在差异,这些研究为重金属污染的土壤修复以及抗性性状的筛选提供了一定的参考.本研究中,我们发现,雌雄美洲黑杨之间的重金属积累能力存在显著差异,并且这种差异会因污染条件的不同发生变化,例如,Pb污染条件下,接种丛枝菌根真菌的雌雄之间Pb积累量没有显著的性别差异;但是,Cd污染条件下,与接种丛枝菌根真菌的雌株相比,对应雄株积累的总Cd积累量较高;复合污染条件下,接种丛枝菌根真菌的雌株Pb和Cd的总积累量均较高.本研究中,采用砂培基质是利用其良好的通透性而有利于菌根真菌的侵染,从而认识菌根真菌对于植物重金属吸收和积累的效应.然而,砂培基质与土壤存在一定差别,在相同的污染水平,砂培基质较低的有机质含量会增加重金属的生物有效性(Li et al.,2013),所以本研究的结论可能更适用于重金属生物有效性较高的环境.
5 结论(Conclusions)1) Pb/Cd复合污染对于雌雄美洲黑杨Pb和Cd的吸收具有独特的效应,复合污染的效应因美洲黑杨性别差异而表现不同,Pb/Cd复合污染条件下雄株各器官中Pb和Cd表现出协同促进作用,而雌株叶片的Pb和Cd表现出协同促进作用,根系中的Pb和Cd却表现出拮抗作用;
2) 接种丛枝菌根真菌能显著提高雌雄美洲黑杨的Pb和Cd富集系数,能不同程度提高2种性别对Pb和Cd的积累能力,降低Pb和Cd向雌雄美洲黑杨地上部分的迁移率;
3) 雌雄美洲黑杨积累重金属能力的差异因污染条件的不同而不同.因此,本研究表明接种耐性菌根真菌能提高美洲黑杨的重金属积累与修复能力,对于不同重金属污染条件下,应根据实际情况选择合适的性别品系进行造林和生态修复.
| [${referVo.labelOrder}] | Ali N, Masood S, Mukhtar T, et al. 2015. Differential effects of cadmium and chromium on growth, photosynthetic activity, and metal uptake of Linum usitatissimum in association with Glomus intraradices[J]. Environmental Monitoring and Assessment, 187(6) : 1–11. |
| [${referVo.labelOrder}] | Baldantoni D, Cicatelli A, Bellino A, et al. 2014. Different behaviours in phytoremediation capacity of two heavy metal tolerant poplar clones in relation to iron and other trace elements[J]. Journal of Environmental Management, 146 : 94–99. DOI:10.1016/j.jenvman.2014.07.045 |
| [${referVo.labelOrder}] | Beauchamp V B, Stromberg J C, Stutz J C. 2006. Arbuscular mycorrhizal fungi associated with Populus-Salix stands in a semiarid riparian ecosystem[J]. New Phytologist, 170(2) : 369–380. DOI:10.1111/nph.2006.170.issue-2 |
| [${referVo.labelOrder}] | 曹福亮, 郁万文, 朱宇林. 2012. 银杏幼苗修复 Pb 和 Cd 重金属污染土壤特性[J]. 林业科学, 2012, 48(4) : 8–13. |
| [${referVo.labelOrder}] | Chen L, Hu X, Yang W, et al. 2015. The effects of arbuscular mycorrhizal fungi on sex-specific responses to Pb pollution in Populus cathayana[J]. Ecotoxicology and Environmental Safety, 113 : 460–468. DOI:10.1016/j.ecoenv.2014.12.033 |
| [${referVo.labelOrder}] | Chen L, Wang L, Chen F, et al. 2013. The effects of exogenous putrescine on sex-specific responses of Populus cathayana to copper stress[J]. Ecotoxicology and Environmental Safety, 97 : 94–102. DOI:10.1016/j.ecoenv.2013.07.009 |
| [${referVo.labelOrder}] | Chen L, Zhang L, Tu L, et al. 2014. Sex-related differences in physiological and ultrastructural responses of Populus cathayana to Ni toxicity[J]. Acta PhySiologiae Plantarum, 36(7) : 1937–1946. DOI:10.1007/s11738-014-1570-4 |
| [${referVo.labelOrder}] | 樊霆, 叶文玲, 陈海燕, 等. 2013. 农田土壤重金属污染状况及修复技术研究[J]. 生态环境学报, 2013, 22(10) : 1727–1736. |
| [${referVo.labelOrder}] | Gehring C A, Mueller R C, Whitham T G. 2006. Environmental and genetic effects on the formation of ectomycorrhizal and arbuscular mycorrhizal associations in cottonwoods[J]. Oecologia, 149(1) : 158–164. DOI:10.1007/s00442-006-0437-9 |
| [${referVo.labelOrder}] | Gottel N R, Castro H F, Kerley M, et al. 2011. Distinct microbial communities within the endosphere and rhizosphere of Populus deltoides roots across contrasting soil types[J]. Applied and Environmental Microbiology, 77(17) : 5934–5944. DOI:10.1128/AEM.05255-11 |
| [${referVo.labelOrder}] | He J, Ma C, Ma Y, et al. 2013. Cadmium tolerance in six poplar species[J]. Environmental Science and Pollution Research, 20(1) : 163–174. DOI:10.1007/s11356-012-1008-8 |
| [${referVo.labelOrder}] | 黄晶, 凌婉婷, 孙艳娣, 等. 2012. 丛枝菌根真菌对紫花苜蓿吸收土壤中镉和锌的影响[J]. 农业环境科学学报, 2012, 31(1) : 99–105. |
| [${referVo.labelOrder}] | Karliński L, Rudawska M, Kieliszewska Rokicka B, et al. 2010. Relationship between genotype and soil environment during colonization of poplar roots by mycorrhizal and endophytic fungi[J]. Mycorrhiza, 20(5) : 315–324. DOI:10.1007/s00572-009-0284-8 |
| [${referVo.labelOrder}] | Lei Y B, Korpelainen H, Li C Y. 2007. Physiological and biochemical responses to high Mn concentrations in two contrasting Populus cathayana populations[J]. Chemosphere, 68(4) : 686–694. DOI:10.1016/j.chemosphere.2007.01.066 |
| [${referVo.labelOrder}] | Li T, Liang C, Han X, et al. 2013. Mobilization of cadmium by dissolved organic matter in the rhizosphere of hyperaccumulator Sedum alfredii[J]. Chemosphere, 91(7) : 970–976. DOI:10.1016/j.chemosphere.2013.01.100 |
| [${referVo.labelOrder}] | 李霞, 彭霞薇, 伍松林, 等. 2014. 丛枝菌根对翅荚木生长及吸收累积重金属的影响[J]. 环境科学, 2014, 35(8) : 3142–3148. |
| [${referVo.labelOrder}] | 廖妤婕. 2014. 丛枝菌根真菌作用下桉树对土壤中 Pb, Zn 和 Cd 的耐受机理研究[D]. 南宁:广西大学 |
| [${referVo.labelOrder}] | 林静, 张健, 杨万勤, 等. 2016. 岷江下游五通桥段小型集水区大气降水中 pH 值对重金属含量的影响[J]. 环境科学学报, 2016, 36(4) : 1419–1427. |
| [${referVo.labelOrder}] | 林青, 徐绍辉. 2008. 土壤中重金属离子竞争吸附的研究进展[J]. 土壤 (Soils), 2008, 40(5) : 706–711. |
| [${referVo.labelOrder}] | Lingua G, Franchin C, Todeschini V, et al. 2008. Arbuscular mycorrhizal fungi differentially affect the response to high zinc concentrations of two registered poplar clones[J]. Environmental Pollution, 153(1) : 137–147. DOI:10.1016/j.envpol.2007.07.012 |
| [${referVo.labelOrder}] | Liu H, Yuan M, Tan S, et al. 2015. Enhancement of arbuscular mycorrhizal fungus (Glomus versiforme) on the growth and Cd uptake by Cd-hyperaccumulator Solanum nigrum[J]. Applied Soil Ecology, 89 : 44–49. DOI:10.1016/j.apsoil.2015.01.006 |
| [${referVo.labelOrder}] | 刘晓娜, 赵中秋, 陈志霞, 等. 2011. 螯合剂、菌根联合植物修复重金属污染土壤研究进展[J]. 环境科学与技术, 2011, 34(12) : 127–133. |
| [${referVo.labelOrder}] | Lodge D. 1989. The influence of soil moisture and flooding on formation of VA-endo-and ectomycorrhizae in Populus and Salix[J]. Plant and Soil, 117(2) : 243–253. DOI:10.1007/BF02220718 |
| [${referVo.labelOrder}] | Marmiroli M, Pietrini F, Maestri E, et al. 2011. Growth, physiological and molecular traits in Salicaceae trees investigated for phytoremediation of heavy metals and organics[J]. Tree Physiology, 31(12) : 1319–1334. DOI:10.1093/treephys/tpr090 |
| [${referVo.labelOrder}] | McGonigle T P, Miller M H, Evans D G, et al. 1990. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi[J]. New Phytologist, 115(3) : 495–501. DOI:10.1111/nph.1990.115.issue-3 |
| [${referVo.labelOrder}] | 孙建伶, 罗立强. 2014. 土壤铅, 镉单一和复合污染及与钙, 锌交互作用对韭菜生长和铅镉积累特性的影响[J]. 农业环境科学学报, 2014, 33(6) : 1100–1105. |
| [${referVo.labelOrder}] | 王发园, 林先贵. 2007. 丛枝菌根在植物修复重金属污染土壤中的作用[J]. 生态学报, 2007, 27(2) : 793–801. |
| [${referVo.labelOrder}] | Wang S, Zhao Y, Guo J, et al. 2016. Effects of Cd, Cu and Zn on Ricinus communis L.Growth in single element or co-contaminated soils:Pot experiments[J]. Ecological Engineering, 90 : 347–351. DOI:10.1016/j.ecoleng.2015.11.044 |
| [${referVo.labelOrder}] | Xu Z, Ban Y, Yang R, et al. 2016. Impact of Funneliformis mosseae on the growth, Pb uptake and localization of Sophora viciifolia[J]. Canadian Journal of Microbiology, 62 : 361–373. DOI:10.1139/cjm-2015-0732 |
| [${referVo.labelOrder}] | Yang H, Koide R T, Zhang Q. 2016. Short-term waterlogging increases arbuscular mycorrhizal fungal species richness and shifts community composition[J]. Plant and Soil, 404(1) : 373–384. |
| [${referVo.labelOrder}] | Yu C, Peng X, Yan H, et al. 2015. Phytoremediation ability of Solanum nigrum L. to Cd-contaminated soils with high levels of Cu, Zn, and Pb[J]. Water, Air, & Soil Pollution, 226(5) : 1–10. |
| [${referVo.labelOrder}] | 张伯镇, 雷沛, 潘延安, 等. 2015. 重庆主城区次级河流表层沉积物重金属污染特征及风险评价[J]. 环境科学学报, 2015, 35(7) : 2185–2192. |
| [${referVo.labelOrder}] | 张旭红, 林爱军, 张莘, 等. 2012. 丛枝菌根真菌对旱稻根际 Pb 形态分布的影响[J]. 中国农学通报, 2012, 28(6) : 24–29. |
| [${referVo.labelOrder}] | 张永超, 陈存根, 钟发明, 等. 2011. 重金属铅, 镉在白榆中分布规律和累积特性研究[J]. 西北林学院学报, 2011, 26(5) : 6–11. |