 2016, Vol. 36
2016, Vol. 36



河口区处于河流与海洋的交汇过渡地带,具有独特的环境系统功能与构成、复杂的物质交换和循环体系.从河流输入到河口的污染物,一部分无机态营养物质直接被浮游植物吸收利用,一部分颗粒态有机物则沉降到底泥中不断累积,并在特定的条件下,以底泥释放的形式重新进入水体.据Cowan和Boynton(1996)在美国Mobile湾的研究发现,由底泥释放的营养盐通量可提供浮游植物需氮量的36%、需磷量的25%.可见,底泥营养盐通量不仅是水体营养元素循环系统中的重要一环,而且对河口生态环境也具有一定的影响.国内外学者针对不同地区的底泥营养盐通量及其影响因素已展开了广泛研究,例如,Gardner等(2009)利用15N同位素示踪技术研究了美国Florida海湾水体-底泥界面氮元素的动力学过程,对底泥氨氮及氮气的释放通量进行了测定,并分析了二者与底泥耗氧量之间的关系;Rigaud等(2013)估算了法国Berre泻湖的底泥营养盐、微量元素的通量及底泥耗氧量,并推算出由底泥释放的营养盐对水体藻类营养盐需求量的贡献;邓华健(2004)通过实验室培养试验测定了渤海湾的底泥营养盐通量,指出该通量存在明显的季节与空间差异,并利用数值模型研究发现底泥通量对渤海湾水体的营养盐浓度有很大影响;董慧等(2012)对青岛市李村河口的底部营养盐通量进行了测定,并分析了溶解氧、温度、盐度、生物扰动等环境因素的影响.
珠江河口地区是中国重要的经济中心区域,人口密布,产业高度集中.随着近几十年来人口的迅速增加,大量污染物通过河流及沿岸排放进入珠江口海域,致使珠江口营养盐浓度超标(Huang et al,2003; Yin et al,2004),水体富营养化状况愈加严重(黄小平等,2001).与此同时,大量颗粒态有机物进入珠江口后,沉降进入底泥中并不断累积,成为河口区潜在的二次污染源.研究发现,珠江口底层水体铵盐浓度与底泥释放的铵盐通量有显著的相关性(潘建明等,2002),底泥是水体中氮的重要蓄积库和磷的可能输入源(黄小平,2007).据估计,底泥铵盐的平均释放通量相当于伶仃洋河流总氮输入量的9%和颗粒态有机氮输入量的24%(潘建明等,2002).由此可见,底泥营养盐通量对珠江口的营养盐收支具有不容忽视的贡献.
目前关于珠江口底泥营养盐通量的研究已取得一定的成果(潘建明等,2002; 张德荣等,2006; Zhang et al,2013; Zhang et al,2014),但大多数研究受限于观测资料,面临观测数据不足和难以获取等难题,无法充分反映底泥营养盐通量的时空变化特征,且难以对其影响机制及环境效应展开深入研究.底泥通量模型(Di Toro,2001)的应用则在很大程度上弥补了观测的不足,使研究者能够以有限的观测数据结合数值模拟研究底泥营养盐的释放过程.Brady等(2013)和Testa等(2013)利用多年历史观测数据对Chesapeake湾底泥通量模型进行了校正及验证,成功模拟出研究区域内底泥氮、磷、硅等营养盐释放的变化特征;Wilson等(2013)利用Narragansett湾中型围隔实验的多年观测资料构建了两种不同的底泥模型(双层成岩模型和多层成岩模型),并对两个模型进行了参数优化,进而对比分析两者在底泥耗氧量及营养盐通量的模拟效果,研究结果表明,两个模型均能取得与观测较为一致的结果,其中多层成岩模型的效果相对更好.上述研究已表明,底泥通量模型具有良好的应用前景,能较好地模拟出底泥关键过程的动态变化.在珠江河口,张恒和李适宇(2010)初步建立了水质-底泥耦合模型并将其应用于水体缺氧及初级产生力的模拟分析中;然而,该工作并未对底泥营养盐通量进行验证和分析.基于此,本文以珠江河口为研究对象,在水动力-水质模型的基础上建立底泥通量参数化模块,动态反映水体-底泥中营养盐的时空变化,揭示底泥营养盐通量的特征及其对水体营养盐输入的贡献.
2 数值模型(Numerical models) 2.1 一维河网与三维河口耦合水动力模型珠江口水系复杂,受珠三角河网径流和南海潮汐共同影响.珠江口与河网水系在水流及动力上相互贯通,交互影响,因此,此处采用已验证的一维河网与三维河口耦合水动力模型(胡嘉镗等,2008)对珠江口的水动力过程进行模拟,以充分反映河口区与河网水系之间的交互作用.其中,河网水动力模型基于一维圣维南方程组,采用三级联合解法进行求解,计算范围上至西江高要站、北江石角站、东江博罗站、流溪河老鸦岗站和潭江石咀站(图 1),下至8个入海口门(虎门、蕉门、洪奇沥、横门、磨刀门、鸡啼门、虎跳门、崖门),共概化299条河道及1726个计算断面.三维河口模型选用ECOMSED水动力模块(Blumberg,2002),模拟变量包括水位、三维流速、温度、盐度、湍流动能等,采用模态分裂技术进行求解,计算范围西起镇海湾,东至红海湾,南至外海75 m等深线附近,水平方向划分正交曲线网格速、温度、盐度、湍流动能等,采用模态分裂技术进行求解,计算范围西起镇海湾,东至红海湾,南至外海75 m等深线附近,水平方向划分正交曲线网格183×186(如图 1所示),垂直方向划分17个sigma层.一维河网模型和三维河口模型以8个口门作为连接断面,通过互赋边界实现耦合计算(胡嘉镗等,2008).
 |
| 图 1 一维河网与三维河口耦合水动力模型范围与计算网格 Fig. 1 Model domain and computational grids for the coupled 1D and 3D hydrodynamic model |
底泥-水体的物质交换过程与底泥和水体两个系统密不可分,为了准确模拟珠江口底泥-水体复杂的生物、化学作用,选用从美国环保署WASP(Water Analysis Simulation Program)模式发展而来的RCA(Row-Column AESOP)三维高级别水质模型(Fitzpatrick,2004),该模型涉及浮游植物、碳、氮、磷、硅、溶解氧等多个循环交互系统,共包括20个状态变量(表 1).水质模型各循环系统的作用关系如图 2a所示.基于质量守恒原理并考虑物质的生化过程及源汇项,推导出各水质因子的控制方程,其一般形式可表达为:
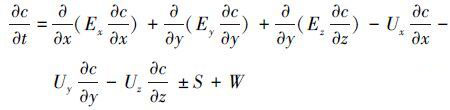
|
(1) |
式中,c为水质因子浓度(mg·L-1);t为时间(s);x、y、z分别表示纵向、横向和垂向坐标;E为扩散系数(m2·s-1);U为对流速度(m·s-1);S为水质因子动力学反应源汇项(mg·L-1·s-1);W为外源输入(mg·L-1·s-1).
 |
| 图 2 水质与底泥耦合模型(RCA-SFM)示意图(a.水质模型RCA,b.底泥通量模型SFM) Fig. 2 Schematic of the coupled water quality and sediment model(RCA-SFM) |
| 表 1 RCA水质模型状态变量 Table 1 State variables in the water quality model RCA |
除了上述复杂的富营养化模块之外,RCA模型另一个突出的特点是耦合了底泥通量模型(Sediment Flux Model,SFM,Di Toro,2001),用以描述底泥的生物化学过程及其与水体之间的动力学关联.SFM可以根据颗粒态有机物(POM,包括POC、PON和POP)沉积通量及底泥中原有的有机物含量,动态模拟出经早期成岩反应后底泥中的POM和营养盐通量及底泥耗氧量(SOD)的时空变化(如图 2b所示).SFM模型将底泥划分为好氧层和厌氧层,好氧层厚度取决于底泥中的溶解氧浓度,通常只有几毫米厚,厌氧层厚度则等于底泥层厚度与好氧层厚度之差;由于底泥好氧层的厚度非常小,好氧层内经早期成岩反应产生的物质通量远远小于厌氧层,因此,SFM不考虑好氧层内发生的早期成岩作用.底泥中的POM分为G1、G2、G3三个等级,各等级所代表的反应半周期不同(Westrich et al,1984),其动力学方程可用下式表示:
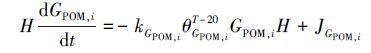
|
(2) |
式中,H为底泥厚度(m); GPOM,i 为各级别POM在底泥中的浓度(g·m-3); kGPOM,i 为各级别POM的反应速率(d-1);T为底泥温度(℃); θGPOM,i 为各级别POM的温度修正系数; JGPOM,i 为从水体至底泥的各级别POM沉降通量(g·m-2·d-1).
NH4+-N、NO3--N、PO43--P等营养盐进入到底泥后,会在好氧层和厌氧层进一步发生降解,如NH4+-N在好氧层中被硝化、NO3--N在缺氧环境及厌氧层中通过反硝化被去除.同时,厌氧层内POM降解产生的无机营养盐也在底泥中参与各项生化反应.好氧层和厌氧层的物质经过生化反应后,其浓度的变化可用方程式表示为:
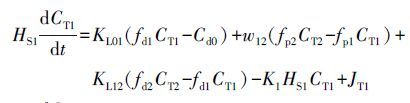
|
(3) |
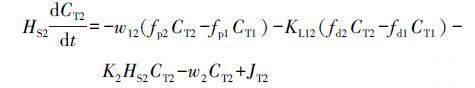
|
(4) |
式中,CT1为好氧层中的物质总浓度(mg·m-3),CT2为厌氧层中的物质总浓度(mg·m-3),fd1为好氧层中溶解态物质比例,fp1为好氧层中颗粒态物质比例,fd2为厌氧层中溶解态物质比例,fp1为厌氧层中颗粒态物质比例,Cd0为底泥间隙水中各类营养盐的浓度(mg·m-3),JT1为好氧层因生化反应产生的物质的内源项(mg·m-2·d-1),JT2为厌氧层内因早期成岩作用和生化反应产生的物质的内源项(mg·m-2·d-1),HS1为好氧层的厚度(m),HS2为厌氧层的厚度(m),KL01为好氧层与上覆水之间的物质传输系数(m·d-1),KL12为厌氧层与好氧层之间的物质传输系数(m·d-1),w12为好氧层与厌氧层之间的颗粒混合速度(m·d-1),w2为厌氧层中物质的沉积速度(m·d-1),K1为好氧层中物质的反应速率(d-1),K2为厌氧层中物质的反应速率(d-1).
底泥营养盐通量的产生与其在上覆水体及底泥好氧层中的浓度有关,同时受底泥-水界面的传质系数影响,依据Fick第一定律,可用以下方程表示:
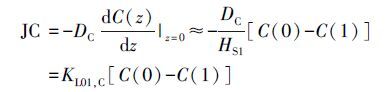
|
(5) |
式中,JC表示某种营养盐通量(mg·m-2·d-1);DC为该物质在好氧层中的扩散系数(m2·d-1); C(0) 、 C(1) 分别为上覆水和好氧层中的营养盐浓度(mg·m-3);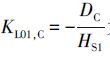
底泥耗氧量(SOD)表示底泥中生化反应所消耗的氧气,计算式为:
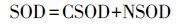
|
(6) |
式中,CSOD是底泥中H2S及CH4氧化的耗氧量(mg·m-2·d-1);NSOD表示底泥中NH4+-N硝化反应产生的耗氧量(mg·m-2·d-1).
RCA和SFM模型在珠江口、切萨比克湾等区域已有应用实例(Brady et al 2013;Testa et al 2013;张恒等,2010),更为详细的模型介绍可参考上述文献.
3 模型设置(Model settings) 3.1 水动力模型设置为了研究夏季珠江口底泥营养盐通量的变化特征,并考虑到构建模型期间边界条件和验证数据的可获得性,一维河网与三维河口耦合水动力模型选择了2个观测资料相对丰富、能够满足建模需求的模拟时段,分别为1999年3—7月和2005年11月—2006年8月,在每个时段的前2个月进行模型积分预热,使模型达到相对稳定的状态.一维河网模型上游边界条件采用高要、石角、博罗水文站的实测流量及老鸦岗和石咀水文站的逐时水位观测数据,三维河口模型外海水位边界采用全球潮汐预测模型(OTIS)在南海北部的优化子集数据,外海温盐边界参考大万山站的历史监测数据进行设定,风速、风向等海表面驱动条件采用美国NCEP每6 h一次的气象数据.三维模型的外模时间步长为5 s(与一维模型的时间步长一致),内模时间步长为30 s.
3.2 水质与底泥耦合模型设置三维水质与底泥耦合模型(RCA-SFM)的模拟时段与水动力模型一致,每个时段的前2个月作为模型积分预热时段,并重复运行3次,以消除初始条件的影响并使模型达到相对稳定的状态.1999年3—7月的河流水质边界主要参考珠江水利委员会在八大口门的常规水质监测数据及前人的研究成果(张恒等,2010);2005年11月—2006年8月的河流水质边界则主要参考南海分局在八大口门的逐月水质监测数据(包括DO、NH4+-N、NO2--N+NO3--N、PO43--P),有机碳、有机氮及有机磷的设置分别参考珠江口的相关调查研究成果(张德荣,2010;王帅,2009;杨婉玲等,2010).外海水质边界的设定主要参考珠江口的相关研究成果(张恒等,2010)及南海分局的水质监测数据.SFM底泥通量模型中POP、POC、PON及营养盐浓度的设置主要参考目前珠江口已有的应用成果(张恒等,2010).水质模型的计算时间步长为60 s.
RCA和SFM模型涉及的参数众多,如何合理确定参数取值是模型应用中的一个难点.本文主要是在参考前人研究成果(张恒等,2010; Fitzpatrick,2004)的基础上,确定珠江口水质与底泥耦合模型参数的取值区间及推荐值,通过扰动主要参数的方式进行敏感性分析,挑选对模拟结果影响相对较大的参数(分析结果表明,敏感性较高的参数主要包括水体中的浮游植物生长速率、硝化速率,以及底泥中的硝化速率、反硝化速率),再针对其中敏感性较高的参数设置不同的取值组合,分析各参数组合驱动下的模拟与观测资料的比对效果,从中选择计算误差相对最小的一套参数作为最终参数取值.整体而言,模型能较好地反映出珠江口水质和底泥物质分布特征(详见后续分析结果),表明所选用的参数(部分取值区间及最终取值见表 2)具有良好的适用性.
| 表 2 水质-底泥耦合模型(RCA-SFM)参数 Table 2 Parameters for the coupled water quality and sediment model(RCA-SFM) |
用于模型验证的观测数据包括:①1999年7月珠江口水污染研究项目的水质及底泥监测资料(李炎等,2002; Chen et al 2004),采样位置见图 3;②2006年7—8月珠江口南海分局夏季航次的水质监测数据,采样位置见图 3.以上验证数据的分布基本上涵盖了研究区域整体范围,并且包含有大面站、连续站两类数据,满足模型验证的需求.
 |
| 图 3 珠江口调查站点位置及分区示意图 Fig. 3 Locations of the sampling stations and subareas in the Pearl River Estuary |
水质模型验证指标包括无机氮(DIN)、磷酸盐(PO43--P)、溶解性硅(DSi)和溶解氧(DO).图 4给出了1999年7月和2006年7月大面站表层DIN和PO43--P的模拟值与观测值,比对结果说明水质模型能较好地反映出珠江口营养盐的空间分布特征(浓度从口门向外海逐渐降低);各水质指标的模拟值与观测值的相关性系数基本上均大于0.6,相对平均误差均小于38%,其中,DO和DIN的相对平均误差分别小于17%和30%(表 3).同时对1999年7月的3个连续站点进行了水质验证,其中,C1连续站(靠近内伶仃岛)的结果见图 5.由图可见,连续站水质因子的模拟值与观测值吻合良好,两者的量值相近且在垂向剖面的时间变化趋势上基本一致,其相对平均误差基本在30%以内(表 3),表明水质模型能较准确地刻画出水质要素的时间变化过程.另外需要指出的是,一维河网与三维河口耦合水动力模型的验证结果在文献(胡嘉镗等,2008)中已有详细介绍,此处不再赘述,仅在表 3列出盐度的验证统计指标以供参考.由表 3可见,盐度的模拟值与观测值的相关性系数大于0.9,相对平均误差均小于18%.
 |
| 图 4 大面站表层无机氮、磷酸盐浓度的模拟值与观测值比较 Fig. 4 Comparisons of simulated and observed concentrations of surface DIN and PO43--P at survey stations |
 |
| 图 5 1999年7月23—24日C1连续站无机氮、磷酸盐、溶解性硅及溶解氧浓度的模拟值与观测值比较 Fig. 5 Comparisons of simulated and observed concentrations of DIN,PO43--P,DSi,and DO at time-series station C1 in July,23—24,1999 |
| 表 3 RCA水质模型验证统计指标 Table 3 Validation metrics of the water quality model RCA |
底泥通量模型主要反映底泥与上覆水体中营养盐、溶解氧等物质的交换过程及底泥中各物质的动态变化,为评估底泥通量模型的合理性,分别对底层水和底泥的营养盐浓度、底泥耗氧量及营养盐通量进行验证.其中,用于底层水质验证的观测数据取自文献报道结果(潘建明等,2002),但由于该文中未明确交代底层采样深度,因此统一采用距底部1 m处的模拟值与之进行比对,结果见图 6.如图所示,除个别站点外,各站点的底层水质因子模拟值与观测值均较为接近,相对平均误差均小于37%,而个别站点出现较大偏差,可能与模拟与观测的采样深度不完全一致有关.另外,底泥中的验证数据来源于珠江口水污染研究项目(李炎等,2002; Chen et al 2004),包括了1999年7月底泥孔隙水的NH4+-N浓度及底泥耗氧量(SOD)观测资料,验证结果见图 6.由图可见,除15号站点外,底泥孔隙水中NH4+-N浓度的模拟值与观测值均吻合良好,相对平均误差为19%,而15号站点出现较大的偏差,可能是该区域底泥初始条件设置与实际情况存在偏差所致;SOD的模拟值与观测值亦较为接近,相对平均误差为20%,而在13号站点的潮周期标准偏差相对较大,其主要原因在于该站点位处淡水与海水交互地带,水动力条件随潮汐振荡变化明显,从图 6中亦可发现,该站点的底层DO、DIN等水质浓度亦出现明显的潮周期变化.从整体来看,底泥通量模型能较好地模拟出底泥中营养盐的浓度变化及相关生物化学过程.
 |
| 图 6 1999年7月底层水质浓度和底泥孔隙水中NH4+-N浓度、SOD的模拟值与观测值比较 Fig. 6 Comparisons of simulated and observed concentrations of water quality components at the bottom and NH4+-N concentrations and SOD in the benthic sediments in July 1999 |
关于珠江口底泥营养盐通量,目前已有的观测资料比较有限,难以定量评估底泥通量模型的模拟效果.下面主要将模型计算的底泥营养盐通量与文献报道结果进行量值与空间分布趋势上的比对,结果列于表 4和表 5.其中,表 4给出了1999年7月底泥NH4+-N通量的模型计算值及相关研究的估算结果(李炎等,2002; Chen et al 2004).需要指出的是,基于Fick第一定律估算得到的通量值因不考虑NH4+-N从底泥向沉积物-水界面传输过程中发生的生物化学反应,其结果较之实际通量会相对偏小,而通过假定推算出来的最大通量值则会明显偏高;由表 4可见,SFM模型计算的通量值处于文献估算通量与最大通量值之间,且相对更接近于估算通量,说明模型通量计算结果合理.另外,由表 5给出的对比结果可知,模型通量计算值的分布与相关的研究结果较为接近,反映出夏季时珠江口NH4+-N主要从底泥向水体输送、NO23主要从水体向底泥输送及PO43--P既可能由水体向底泥输入,也可能由底泥向水体输送的趋势,与前人的研究结论一致(黄小平,2007; Zhang et al 2014);在珠江口近海,其分布与前人的研究结果也非常接近(Zhang et al 2013;张德荣等,2006),夏季时NH4+-N和PO43--P主要从底泥向水体输送,NO23主要从水体向底泥输送.
| 表 4 1999年7月底泥NH4+-N通量的模拟值与文献报道结果的对比 Table 4 Comparisons of sediment NH4+-N fluxes from our model results and from literature report in July 1999mmol·m-2·d-1 |
| 表 5 底泥营养盐通量的模拟值与前人研究结果的比较 Table 5 Comparisons of sediment nutrient fluxes from our model results and from previous studies |
基于上述已验证的水质-底泥耦合模型的模拟结果,分析珠江口夏季底泥营养盐通量的分布特征.图 7给出了1999年7月和2006年7月底泥营养盐通量的月均分布.由图可见,在珠江口海域,夏季时NH4+-N主要是由底泥向水体输送,底泥通量是水体NH4+-N的重要来源.从空间分布上看,底泥NH4+-N通量从河口内向外海方向递减,越靠近外海其通量越小,表 5中的区域统计结果显示,伶仃洋的底泥NH4+-N通量变化范围为0.24~8.88 mmol·m-2·d-1,平均值为4.59 mmol·m-2·d-1,珠江口近海的NH4+-N通量变化范围为-0.01~4.14 mmol·m-2·d-1,平均值为0.59 mmol·m-2·d-1;受沿岸排海污染源的影响,最大值出现在伶仃洋的东部沿岸及深圳湾内沿岸,其次主要集中在口门附近,这是由于经上游河流输入到口门的POC、PON、POP等有机物在口门附近发生沉降进入底泥,经早期成岩作用产生的NH4+-N向水体输送所致.同时,比较两个年份的通量分布可发现,2006年夏季底泥NH4+-N通量普遍增大,这有可能是因为底泥中长期积累下来的有机物经过降解产生NH4+-N,提高了间隙水中的NH4+-N浓度,从而使其释放通量增大.经统计,与1999年夏季相比,2006年夏季底泥间隙水中的NH4+-N浓度平均升高了21%.
底泥NO23通量的表现则与NH4+-N的相反,1999年及2006年夏季均表现为从水体向底泥的强烈输送(图 7),底泥对水体NO23呈现“汇”的作用.从空间分布上看,口门及其附近海域的NO23通量值最大,局部可达到-10 mmol·m-2·d-1以上;进入河口区后底泥通量绝对值会迅速减小,这种情况在伶仃洋尤为明显,例如2006年7月的NO23通量值从虎门附近海域的-7.14 mmol·m-2·d-1迅速变化至内伶仃岛西侧的-1.43 mmol·m-2·d-1,到河口下游已接近为0.这主要是因为输入口门的水体NO23浓度明显高于底泥中的NO23浓度,导致NO23从水体向底泥的强烈输送,表现出较大的底泥通量,同时在向海输送的过程中,水中NO23还受到海水稀释及藻类生长吸收的影响,在上述3个方面的共同作用下促使NO23浓度快速降低,水体与底泥的NO23浓度差也快速减小,从而使其通量值迅速下降.经统计,伶仃洋的底泥NO23通量变化范围为-10.06~-0.14 mmol·m-2·d-1(表 5),平均值为-4.87 mmol·m-2·d-1;珠江口近海的NO23通量较小,变化范围为-1.45~0.68 mmol·m-2·d-1,平均值为-0.17 mmol·m-2·d-1,远小于伶仃洋的平均通量,说明口门输入对底泥NO23通量有较大的影响,大量的NO23输送进入珠江河口,底泥成为NO23的汇.
珠江口夏季底泥PO43--P通量的输送方向比较不规律,PO43--P可能从底泥向水体输送,也可能是从水体向底泥输送,整体上呈现从底泥向水体输送、从河口向外海递减的分布规律.在伶仃洋东部及深圳湾内沿岸,受沿岸排放点源的影响,底泥主要表现为PO43--P的汇;经统计,伶仃洋的底泥PO43--P通量变化范围为-0.37~0.41 mmol·m-2·d-1(表 5),平均值为0.07 mmol·m-2·d-1,珠江口近海的通量则在-0.12~0.09 mmol·m-2·d-1之间,平均为0.02 mmol·m-2·d-1.另外,对比两个年份的底泥PO43--P通量分布可发现,2006 年夏季PO43--P的通量值相对于1999年夏季有较明显的升高,其原因与底泥NH4+-N通量的类似,主要是底泥中的PO43--P浓度平均升高了54%,从而促进底泥PO43--P的释放.
 |
| 图 7 1999年及2006年7月珠江口月均底泥营养盐通量模拟结果 Fig. 7 Monthly-mean simulated sediment nutrient fluxes in the Pearl River Estuary in July of 1999 and 2006 |
将珠江口划分为AREA1和AREA2两个区域(图 3),统计分析底泥通量对营养盐输入的贡献,其中,AREA1代表伶仃洋海域,其外源营养盐主要来自珠江口东四口门的输入,AREA2则包括了珠江口8个口门,考虑的是底泥通量对整个珠江口营养盐输入的影响.统计结果见表 6.
由表 6统计结果可知,底泥NH4+-N通量对珠江口无机氮(DIN)输入有一定的贡献,在伶仃洋海域(AREA1)其通量相当于东四口门陆源DIN输入量的8%,而在整个珠江口范围内(AREA2)其通量可达至相当于14%以上的陆源DIN输入量;相对于陆源总氮(TN)输入量而言,两个夏季模拟时段的底泥NH4+-N通量的贡献程度为6%~12%,与潘建明等(2002)估计的9%较为接近.相对于底泥NH4+-N通量所起的“源”作用,底泥NO23通量则是去除水体中NO23的一个途径,底泥是水体NO23的汇.统计结果表明,通过底泥NO23通量的输送可去除水体9%~16%的陆源DIN输入量或7%~12%的陆源TN输入量.可见,底泥NO23通量与NH4+-N通量一样,对伶仃洋乃至整个珠江口内的营养盐循环具有重要影响.
| 表 6 底泥通量对营养盐输入的贡献 Table 6 Contribution of sediment fluxes to nutrient inputs |
底泥PO43--P通量的统计结果显示,其相对贡献在不同年份变化明显,1999年和2006年夏季在伶仃洋的底泥通量分别相当于-2%、15%的陆源总磷(TP)输入量,在整个珠江口内则分别相当于2%、22%;底泥PO43--P通量在两个模拟时段对无机磷(DIP)输入的贡献亦有类似的变化规律.不难看出,2006年夏季底泥通量的贡献较之1999年夏季有明显提高,例如,1999年夏季珠江口内的PO43--P底泥通量仅相当于陆源DIP输入的4%,在2006年夏季却增至31%,这个增长趋势在伶仃洋海域内同样明显.
5 结论(Conclusions)1) 经观测数据验证表明,本文所建立的珠江口三维水质与底泥耦合模型成功地描述了珠江口的水质-底泥动态变化过程,各水质要素的模拟结果在变化趋势及量值上均与观测结果吻合良好,计算误差控制在合理范围内,底泥中营养盐、溶解氧浓度及通量的模拟结果与实测值也较为接近,模型能够较好地模拟出底泥中营养盐浓度及通量的变化特征,可为后续关于底泥早期成岩作用、底泥与水体物质交换过程等问题的研究提供技术支持.
2) 耦合模型的模拟结果表明,底泥NH4+-N和PO43--P的通量主要是从底泥向水体输送,底泥对水体氮磷元素起“源”的作用,从整个珠江口来看,两个模拟年份夏季的通量相当于陆源DIN、DIP输入量的14%~16%、4%~31%,相当于陆源TN、TP输入量的11%~12%、2%~22%;底泥NO23通量则主要从水体向底泥输送,底泥是水体NO23重要的汇,从整个珠江口来看,两个模拟时段的底泥通量相当于去除了水体中12%~16%的陆源DIN输入量、9%~12%的陆源TN输入量.底泥营养盐通量对珠江口的营养盐循环有明显的影响.在空间分布上,底泥NH4+-N、NO23、PO3--P通量的量值均表现为从河口向外海方向递减,在伶仃洋NH4+-N、NO23、PO43--P底泥通量平均分别为4.59、-4.87、0.07 mmol·m-2·d-1,在珠江口近海NH4+-N、NO23、PO43--P底泥通量平均分别为0.59、-0.17、0.02 mmol·m-2·d-1.
3) 目前有关珠江口底泥营养盐通量的观测资料非常有限,给本文的模型验证工作带来了一定的限制,因此需待加强相关监测调查,以获取时空间覆盖面更为广泛的数据,进一步率定及改进模型,从而提高其模拟精度.另一方面,受篇幅所限,本文未对模型参数的敏感性展开分析,同时也未考虑由参数取值变化所引起的计算结果的不确定性,在后续的研究工作中,需要全面分析参数敏感性及综合考虑边界条件、污染负荷输入、参数设置等带来的模型不确定性的影响.
| [1] | Blumberg A F.2002.A Primer for ECOMSED Version 1.3 User Manual[R].Mahwah, New Jersey:HydroQual,Inc. |
| [2] | Brady D C, Testa J M, Di Toro D M, et al. 2013. Sediment flux modeling: calibration and application for coastal systems[J]. Estuarine,Coastal and Shelf Science , 117 : 107–124. DOI:10.1016/j.ecss.2012.11.003 |
| [3] | Chen J C, Heinke G W, Zhou M J. 2004. The Pearl River estuary pollution project(PREPP)[J]. Continental Shelf Research , 24 (16) : 1739–1744. DOI:10.1016/j.csr.2004.06.004 |
| [4] | Cowan J L, Boynton W R. 1996. Sediment-water oxygen and nutrient exchanges along the longitudinal axis of Chesapeake Bay: seasonal patterns,controlling factors and ecological significance[J]. Estuaries , 19 (3) : 562–580. DOI:10.2307/1352518 |
| [5] | 邓华健.2004.渤海湾沉积物-水界面营养盐交换通量的研究[D].天津: 天津大学 http://cdmd.cnki.com.cn/article/cdmd-10056-2006052039.htm |
| [6] | Di Toro D M. 2001. Sediment Flux Modeling[M]. New York: Wiley-Interscience . |
| [7] | 董慧, 郑西来, 张健.2012. 污染河口区沉积物-水界面营养盐交换通量的实验研究[J]. 海洋环境科学 , 2012, 31 (3) : 423–428. |
| [8] | Fitzpatrick J J.2004.User's Guide for RCA Release 3.0[R].Magwag,New Jersey:HydroQual Inc. |
| [9] | Gardner W S, McCarthy M J. 2009. Nitrogen dynamics at the sediment-water interface in shallow,sub-tropical Florida Bay: why denitrification efficiency may decrease with increased eutrophication[J]. Biogeochemistry , 95 (2/3) : 185–198. |
| [10] | 胡嘉镗, 李适宇.2008. 珠江三角洲一维盐度与三维斜压耦合模型[J]. 水利学报 , 2008, 39 (11) : 1174–1182. |
| [11] | 黄小平. 2007. 中国南海珠江口污染防治与生态保护[M]. 广州: 广东经济出版社 . |
| [12] | 黄小平, 田磊, 彭勃, 等.2001. 珠江口海域环境污染研究进展[J]. 热带海洋学报 , 2001, 29 (1) : 1–7. |
| [13] | Huang X P, Huang L M, Yue W Z. 2003. The characteristics of nutrients and eutrophication in the Pearl River estuary,South China[J]. Marine Pollution Bulletin , 47 (1) : 30–36. |
| [14] | 李炎,董礼先.2002.珠江水域污染研究计划课题研究报告[R].香港: 香港科技大学 |
| [15] | 潘建明, 周怀阳, 扈传昱, 等.2002. 夏季珠江口沉积物中营养盐剖面分布和界面交换通量[J]. 海洋学报 , 2002, 24 (3) : 52–59. |
| [16] | Rigaud S, Radakovitch O, Couture R M, et al. 2013. Mobility and fluxes of trace elements and nutrients at the sediment-water interface of a lagoon under contrasting water column oxygenation conditions[J]. Applied Geochemistry , 31 : 35–51. DOI:10.1016/j.apgeochem.2012.12.003 |
| [17] | Testa J M, Brady D C, Di Toro D M, et al. 2013. Sediment flux modeling: Simulating nitrogen,phosphorus,and silica cycles[J]. Estuarine,Coastal and Shelf Science , 131 : 245–263. DOI:10.1016/j.ecss.2013.06.014 |
| [18] | 王帅.2009.珠江八大口门表层沉积物和水体中氮的分布特征和影响因素[D].上海:上海海洋大学 |
| [19] | Westrich J T, Berner R A. 1984. The role of sedimentary organic matter in bacterical sulfate reduction: The G model tested[J]. Limnology and Oceanography , 29 (2) : 236–249. DOI:10.4319/lo.1984.29.2.0236 |
| [20] | Wilson R F, Fennel K, Mattern J P. 2013. Simulating sediment-water exchange of nutrients and oxygen: A comparative assessment of models against mesocosm observations[J]. Continental Shelf Research , 63 : 69–84. DOI:10.1016/j.csr.2013.05.003 |
| [21] | 杨婉玲, 赖子尼, 魏泰莉, 等.2010. 2006年珠江入海口营养现状调查及生态危害评价[J]. 广东农业科学 , 2010, 37 (11) : 196–199. |
| [22] | Yin K D, Lin Z F, Ke Z Y. 2004. Temporal and spatial distribution of dissolved oxygen in the Pearl River Estuary and adjacent coastal waters[J]. Continental Shelf Research , 24 (16) : 1935–1948. DOI:10.1016/j.csr.2004.06.017 |
| [23] | 张德荣.2010.珠江口颗粒磷的形态、生物可利用性及其动力学研究[D].广州:中国科学院广州地球化学研究所 |
| [24] | 张德荣, 陈繁荣, 杨永强, 等.2006. 夏季珠江口外近海沉积物/水界面营养盐的交换通量[J]. 热带海洋学报 , 2006, 24 (6) : 53–60. |
| [25] | 张恒, 李适宇.2010. 基于改进的RCA水质模型对珠江口夏季缺氧及初级生产力的数值模拟研究[J]. 热带海洋学报 , 2010, 29 (1) : 20–31. |
| [26] | Zhang L, Wang L, Yin K D, et al. 2013. Pore water nutrient characteristics and the fluxes across the sediment in the Pearl River estuary and adjacent waters,China[J]. Estuarine,Coastal and Shelf Science , 133 : 182–192. DOI:10.1016/j.ecss.2013.08.028 |
| [27] | Zhang L, Huang X P, Wang L, et al. 2014. Spatial and seasonal variations of nutrients in sediment profiles and their sediment-water fluxes in the Pearl River Estuary,Southern China[J]. Journal of Earth Science , 25 (1) : 197–206. DOI:10.1007/s12583-014-0413-y |