 2016, Vol. 36
2016, Vol. 36



2. 福建省污染控制与资源循环重点实验室(福建师范大学), 福州 350007;
3. 中国环境科学研究院, 北京 100012
2. Fujian Key Laboratory of Pollution Control and Resource Recycling, Fuzhou 350007;
3. Chinese Research Academy of Environmental Sciences, Beijing 100012
浮游植物是水体“初级生产力”的一个表现,其丰度、群落结构与富营养化紧密相关,已被广泛用于富营养化的预警和评价(Tett,2004).有学者提出,在研究浮游植物时,将其浮游阶段与其休眠阶段结合起来作为一个整体研究,在研究季节性水华时,除了考虑营养盐波动、摄食压力等条件外,还需考虑浮游植物休眠期的作用(吴天灵,2009).目前,国内学者集中研究了水华暴发时期藻类生物量的变化,而对水华消退之后,水体底部沉积物中微藻变化情况报道较少(万能等,2010;赵汉取等,2009).付永虎等(2009)通过调查南海典型海域沉积物中浮游植物休眠体,鉴定出浮游植物休眠体83种,隶属于47个属,其中,甲藻孢囊51种,硅藻休眠体(包括休眠抱子和休眠细胞)29种.
研究表明,有些浮游植物在秋、冬季生长环境不利时,将下沉到沉积物表面,呈休眠状态,并且在沉积物中保持长时间的活性;春季,少部分微藻因腐烂和分解损失外,其他微藻有重新进入上覆水体的能力,到夏季增殖而大量暴发(Reynolds,1984;Bostr m et al.,1989;Brunberg et al.,2002).研究显示,冬季上覆水和沉积物中存在的微藻对夏季水华暴发强度有显著影响,如果冬季上覆水中没有藻类,夏季水华藻类生物量将降低64%,而冬季沉积物中没有微藻,夏季水华藻类生物量将降低50%(Jolanda et al.,2005).用“N”示踪法研究发现,在沉积物中过冬的微藻休眠体是夏季水华的暴发藻类之一(Preston,1980).浮游植物休眠体被认为是沉积物中特殊的生物类群,水体的营养盐含量、水温、盐度、沉积物的类型、颗粒大小和沉积速率、人为因素等对其种类组成和分布特征都有一定的影响(Monhamed et al.,2011;王朝晖等,2014;Sildever et al.,2015;Moore et al.,2015).姜霞等(2010)对巢湖沉积物微藻生物量季节性变化特征的研究表明,水体富营养程度较高的西部湖区,其沉积物表层(0~2 cm)微藻生物量显著高于富营养化程度相对低的东部湖区,除了呈现空间差异,沉积物微藻生物量也表现出时间差异;阎荣等(2004)对太湖的研究调查也表明,沉积物中微藻生物量在温度较低的4月高于同年温度较高的6月.微藻休眠体种类组成和分布特征反映了一段时间内藻类的种类组成及群落结构等信息(王朝晖等,2014),其萌发为水体提供了大量的藻类营养细胞,揭示未来水华发生的潜势(Persson,2001;Ishikawa et al.,2014),因此,休眠体被认为是水华和赤潮发生的“种源”(张琪等,2012;Anderson et al.,2005),可以把沉积物微藻休眠体当作水体环境变化的生物指示指标和信号载体.
随着经济和社会发展的需要,福建省大多数河流建设了梯级水电站,大坝的建设使水流变缓,水体易发生富营养化(周亮进等,2008).外源营养盐的持续输入,加上沉积物内源营养盐释放,以及在合适的气象水文条件下,水中氮和磷浓度增加,易暴发藻类水华,而产毒藻的蓝藻水华威胁着人们的饮水安全(肖兴复等,2005).九龙江北溪流域建设了近400个梯级水库,曾于2009年1月暴发过甲藻门佩氏拟多甲藻(Peridiniopsis penardii)水华.沉积物微藻种类、丰度及分布对浮游植物的群落结构有重要影响.因此,本研究以九龙江北溪江东水库为研究对象,于2011年研究沉积物中微藻的时空分布规律及其影响因素,这对于江东水库富营养化的预警和防控,保障饮水安全,实现水资源的可持续利用具有重要的理论意义和现实价值.
2 材料与方法(Material and methods) 2.1 研究区域概况九龙江(东经116°50′~118°02′,北纬24°12′~25°44′)是福建省第二大河,位于福建省西南部,流经龙岩、漳州、厦门三市,其地理位置及研究区域位置如图 1所示.九龙江流域由3条主要支流河流构成:北溪、西溪、南溪,河流全长285 km,流域面积14741 km2.其中,九龙江北溪是所经流域的重要饮用水源,它为流经之地(龙岩、华安、漳州、厦门)近400万人提供生活必须的饮用水.北溪流全长274 km,流域面积9640 km2.据九龙江北溪浦南水文站统计,九龙江北溪平均径流量为83亿m3·s-1,径流深度为963 mm,因此,九龙江北溪属于丰水带河流.地理位置处于北回归线附近,为亚热带季风气候,多年平均气温19.9~21.1 ℃,多年平均降水量1400~1800 mm.且所处地区为信风带,受到春夏锋面雨及夏秋台风带来的降水的影响,因此,径流量的年内分布也是不均匀的,丰水期为5—7月,其中,最大流量出现在6月,最小流量出现在1月.本研究选取九龙江北溪——江东库区作为研究对象.
 |
| 图 1 九龙江江东水库地理位置图 Fig. 1 Location of Jiangdong reservoir in Jiulong River |
根据九龙江江东水库段河道型库区,本研究采样断面包括河流区、河流-湖泊缓冲区、湖泊区,以及特殊点位(如支流入口处),于2011年4、6、9、12月采用蚌式采泥器、GPS定位采集九龙江江东库区8个断面的表层沉积物样品,其经、纬位置如表 1所示.将采集的表层沉积物放在干净的聚乙烯盆中混合均匀,迅速装袋,排尽袋中空气,密封避光保存,运回实验室,4 ℃避光冷藏以备后期分析.全过程保持黑暗、低温.在每次测试取样前,需将样品再次混合均匀,以消除取样误差.沉积物采集的同时,用便携式溶解氧仪同步记录上覆水水温和溶解氧.
| 表 1 九龙江江东库区采样断面经纬度 Table 1 The geological position of the main sampling points of Jiangdong reservoir in Jiulong River |
沉积物采样时同步采集1 L上覆水体,加15 mL鲁格氏碘液固定,运回实验室,静置48 h后,将样品浓缩至30 mL,加入40%甲醛溶液保存,其用量为样品量的4%,完成预处理.后期取已完成预处理的样品0.1 mL置于0.1 mL(20 mm×20 mm)的计数框内,在生物显微镜下,以10×40倍进行观察计数,而后采用视野法按照公式(1)计算上覆水体中浮游植物的细胞丰度.
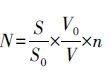
|
(1) |
式中,N为1 L上覆水中细胞丰度(cells·L-1),S为计数框面积(mm2),S0为计数视野面积(mm2),V0为浓缩后体积(mL),V为计数体积(mL),n为计数所得平均每个视野藻类数目(cells).
2.3.2 表层沉积物基本性质测定方法九龙江江东库区表层沉积物粒径使用 Mastersize 2000型激光粒度分析仪进行测试;九龙江江东库区表层沉积物各形态磷含量的测定采用欧洲发展框架委员会推荐的SMT法,该方法可以提取5种形态磷,即总磷、有机磷、无机磷、铁铝结合态磷及钙结合态磷(Ruban et al.,2001);碳、氮含量使用元素分析仪(德国Elementar Vario EL III)进行测试;沉积物含水率的测定采用烘干法(全国农业技术推广服务中心,2006).
2.3.3 表层沉积物微藻丰度与分布分析沉积物藻类分析采用悬浮法,具体操作如下:称取混合均匀的表层沉积物1.00 g,置于干燥洁净的小烧杯中,加入100 mL灭菌蒸馏水,充分混匀,移取30 mL混合液至事先做好标识的试剂瓶内.加入鲁哥氏碘液,用量为样品量的1.5%,并加入40%甲醛溶液保存,其用量为样品量的4%.按照章宗涉等(1991)的淡水浮游植物图谱对各种藻类进行判定.
取处理后的样品0.1 mL置于0.1 mL的计数框内,在10×40倍的倒置显微镜(Nikon TS100)下进行观察计数,而后采用视野法,按照公式(2)计算沉积物的藻类数量,最后将其换算成细胞丰度(cells·g-1(以底泥干重计)).
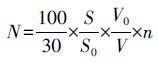
|
(2) |
式中,N为1 g底泥中细胞丰度(cells·g-1(以底泥湿重计)),S为计数框面积(mm2),S0为计数视野面积(mm2),V0为浓缩后体积(mL),V为计数体积(mL),n为计数所得平均每个视野藻类数目(cells).
2.3.4 沉积物甲藻孢囊丰度分析方法甲藻孢囊样品的清洗和富集方法如下:称取混匀后的2 g沉积物样品,加入30 mL灭菌后的蒸馏水置于超声波振荡器中超声分散2 min.将分散后的样品均匀地倒在筛绢上,用灭菌后的蒸馏水冲洗筛上物,收集过筛后的液体;按此方法依次通过200 μm、60 μm孔径的筛绢,将含有孢囊的60 μm孔径筛绢过筛后的液体均匀地倒在20 μm孔径筛绢上,反复冲洗,收集20 μm孔径筛绢的筛上物,并用灭菌的蒸馏水定容至5 mL,即可得到甲藻孢囊富集样品(Matsuoka,2000).根据淡水甲藻孢囊图谱,包括孢囊形状、表面纹饰、细胞壁结构、颜色和萌发孔等特征进行显微镜镜检,并结合孢囊萌发后营养细胞的PCR结果,对甲藻孢囊的种属进行判定.
甲藻孢囊丰度分析采用显微镜镜检法,具体方法如下:吸取1 mL甲藻孢囊富集样品放置于1 mL(20 mm×40 mm)计数框上,进行全视野计数,而后按照公式(3)进行换算,得到该断面的表层沉积物甲藻孢囊丰度.
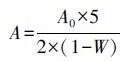
|
(3) |
式中,A为i断面沉积物中甲藻孢囊数量(cells·g-1(以底泥干重计));A0为1 mL样品中含有的甲藻孢囊数量(cells·g-1(以底泥干重计));W为i断面沉积物含水率.
3 结果与讨论(Results and analysis) 3.1 上覆水中藻类丰度和结构分析对九龙江北溪江东库区4个主要断面龙津溪支流汇入口、马洋溪支流、石龟头回水区、江东取水口(J1、J5、J6、J8)的上覆水藻类进行观察,结果如图 2所示.4月上覆水中的藻类细胞丰度为全年最高,藻类丰度为5.56×105 ~1.23×106 cells·L-1,以隐藻门、裸藻门、硅藻门、绿藻门为主,隐藻门和裸藻门分别占水体藻类的17.61%~37.83%和29.24%~42.29%,龙津溪支流汇入口断面(J1)甚至达到了106 cells·L-1.6月藻类丰度为4.19×105 ~6.87×105 cells·L-1,以蓝藻门、硅藻门、绿藻门为主,蓝藻门占水体浮游植物的32.2%~63.89%;9月藻类丰度为6.24×105 ~8.56×105 cells·L-1,以绿藻门、裸藻门、隐藻门、硅藻门为主,绿藻门和裸藻门分别占水体浮游植物的10.00%~40.00%和23.85%~49.02%;12月藻类丰度为2.28×105 ~5.38×105 cells·L-1,包括硅藻门、绿藻门、隐藻门、裸藻门、甲藻门等,藻类多样性比较好,藻细胞丰度为全年最低.
 |
| 图 2 2011年九龙江江东库区上覆水浮游植物藻类丰度分析 Fig. 2 Analysis of algae in the overlying water in different seasons of Jiangdong Reservoir in 2011 |
九龙江江东库区大部分断面表层沉积物平均粒径在4~10 μm之间,大部分为粉砂质粘壤土,大多数断面沉积物粉粒所占比例为52.4%~72.6%.4、6、12月的表层沉积物粒径在断面上区别很小;9月的表层沉积物龙津溪支流汇入口(J1)和干流河道拐弯处(J3)砂粒比例提高,占到到30%~32%,其余断面主要以粉粒为主.
3.2.2 表层沉积物碳、氮、磷含量分析九龙江江东库区主要断面龙津溪支流汇入口、马洋溪支流、石龟头回水区、江东取水口(J1、J5、J6、J8)表层沉积物碳、氮含量如图 3所示:该区域总碳含量为9140.2~17760.0 mg·kg-1,总氮含量为950.0~1600.4 mg·kg-1.从上游至下游呈现递减的趋势,J1、J5断面为支流断面,其碳和氮含量明显高于库区断面J6、J8,这主要是由于支流断面有较多的人类活动,带来较多的农业污染源,使得表层沉积物中的有机质含量、氮营养盐含量较高的现象.
 |
| 图 3 2011年九龙江江东库区表层沉积物总氮(a)和总碳(b)含量 Fig. 3 Content of TN(a)and TC(b)of surface sediment in Jiangdong Reservoir in 2011 |
九龙江江东库区主要断面总磷含量为483.7~856.2 mg·kg-1,以IP(无机磷)为主,含量为272.6~557.6 mg·kg-1,占总磷质量分数的52.80%~87.70%,其次为OP(有机磷),含量为112.0~296.6 mg·kg-1,占总磷质量分数的19.23%~42.79%(图 4).在无机磷组成中,以Fe/Al结合态磷为主,
 |
| 图 4 2011年九龙江江东库区表层沉积物各形态磷含量 Fig. 4 Content of various forms of phosphorus in surface sediment in Jiangdong Reservoir in 2011 |
因为PO43-与沉积物中Fe3+吸附形成固态二价FeOOH-PO3-4化合物,当上覆水水温高、溶解氧低,水库底层处于厌氧状况,这时Fe/Al结合态磷容易溶解,释放到上覆水中(刘培怡,2011;鲁婷等,2013).
3.3 表层沉积物中微藻丰度分析图 5为九龙江江东库区沉积物微藻分布情况.沉积物中微藻丰度范围在7.41×105~2.17×106 cells·g-1(以干重计),主要为硅藻门和绿藻门为主.硅藻在全年的沉积物藻类中占主要优势,丰度达到3.66×105~1.58×106 cells·g-1(以干重计),占沉积物微藻的45.52%~86.42%,其优势种属为直链藻(Melosira).硅藻在沉积物中的丰度较为稳定,全年变化较少,均在105 cells·g-1(以干重计)以上,这主要与硅藻是一种广温性藻类有关,硅藻一般可以生长的温度范围为15~31 ℃,江东水库水温适宜,适合硅藻生长,因此,硅藻借此成为全年沉积物中的优势藻类.沉积物中绿藻丰度达到1.09×105~6.16×105 cells·g-1(以干重计),其优势种属为栅藻(Scenedesmus),占沉积物微藻的5.96%~35.55%.沉积物甲藻孢囊丰度在102 cells·g-1(以干重计)范围内,主要种属为拟多甲藻属,所占比例约为70%~80%,其他主要为裸甲藻属.
 |
| 图 5 2011年九龙江江东库区沉积物微藻分析 Fig. 5 Analysis of algaes in the surface sediments in different seasons of Jiangdong Reservoir in 2011 |
时间上,12月沉积物中甲藻孢囊丰度明显高于其他月份(p<0.05,n=27).冬季水温较低,不利用水体中甲藻生长,将下沉到沉积物表面,呈休眠状态形成甲藻孢囊,同时水温较低不适合甲藻孢囊萌发,因而12月积累在沉积物中的甲藻孢囊数量相比于其他月份多.空间上,沉积物微藻总体呈现上游向下游递减的趋势,高值出现在12月的马洋溪支流(J5)断面.库区断面的沉积物微藻丰度在全年相对稳定.
根据北溪流域综合规划,干流河段从上游到下游共规划上百个梯级水电站,江东水库和西陂水库是其中两个水库(图 1),两个水库均发生过不同程度的水华现象.2012—2013年采集西陂水库表层沉积物,其表层沉积物微藻平均丰度在104~105 cells·g-1(以底泥干重计)左右,主要是硅藻和绿藻,硅藻的平均丰度为105 cells·g-1(以底泥干重计),占微藻比例的81.8%~90.9%以上;其次是绿藻,平均丰度为104 cells·g-1(以底泥干重计),其他门类的藻类较少.由此可以看出,两库区沉积物中微藻种类相似,在空间上具有一定连续性.
使用SPSS18.0软件,对沉积物微藻丰度与上覆水浮游植物/表层沉积物基本性质进行相关性分析,结果如表 3所示.2011年4月、12月上覆水的水温较为接近,在16.5~19.5 ℃之间,而6月、9月上覆水水温较高,可达26 ℃左右.一般而言,水温越高的时候,水体中溶解氧含量越低.由表 3可知,上覆水水温和溶解氧呈显著负相关,相关系数为-0.899(p<0.01).沉积物中蓝藻、隐藻、甲藻孢囊丰度与上覆水水温呈现显著的负相关性(p<0.01),上覆水体的水温对于蓝藻、隐藻、甲藻孢囊的分布是一个重要的影响因子,水温上升有利于沉积物蓝藻、隐藻、甲藻孢囊的萌发;水温下降,这些藻形成休眠体度过不利的环境条件,这与12月九龙江江东库区沉积物甲藻丰度明显高于其他月份相符合.另外,沉积物中蓝藻、隐藻、甲藻孢囊丰度与溶解氧呈现显著正相关(p<0.01).夏季,高温和低溶解氧环境下,沉积物中蓝藻、隐藻、甲藻孢囊丰度相对较低.沉积物微藻丰度与上覆水体浮游植物呈显著负相关,相关系数为-0.596(p<0.05,n=27),反映了沉积物微藻丰度对上覆水体浮游植物的贡献.
| 表 3 2011年九龙江江东库区沉积物微藻丰度与沉积物性质及上覆水环境因子相关性分析 Table 3 Correlation analysis of abundance of microalgae in surface sediments and the characters of sediment and the factors in the overlaying water in Jiangdong Reservoir in 2011 |
沉积物微藻丰度和沉积物中硅藻、绿藻、隐藻、甲藻孢囊丰度显著相关,相关系数分别为0.590、0.738、0.489、0.618(p<0.01,n=27),说明沉积物微藻主要由几种藻类组成(表 3).沉积物微藻丰度、甲藻孢囊分别与沉积物含水率、粉粒呈显著正相关.沉积物的硅藻和总碳、总氮、总磷有显著的正相关关系;当含水率高、粉粒多时沉积物微藻的丰度高.表层沉积物的含水率较高,沉积物颗粒的孔隙度较大,说明该断面主要是以细颗粒为主,也就是此处水流速度较慢,有利于藻体与颗粒结合后沉降下来,因此,该断面的微藻丰度较高,反映了沉积物微藻沿着水流方向的积累.表 3的分析表明,TN与TC呈正相关,相关系数为0.855(p<0.01),这有可能是由于沉积物中TN、TC都来源于同一污染源.Fe/Al结合态磷与上覆水水温呈显著负相关,相关系数为-0.574(p<0.05),与上覆水溶解氧呈显著正相关,相关系数为0.624(p<0.01).高温和缺氧等状态下,Fe/Al结合态磷容易溶解,容易被藻类利用,而水体中藻类旺盛生长,因此,也有利于沉积物中铁/铝结合态磷释放进入上覆水(Hou et al.,2014).
 |
| 图 6 九龙江江东水库沉积物微藻图片(a:粗壮双菱藻(Surirella robusta);b、c:羽纹藻(Pinnularia sp.);d:针杆藻(Symedra sp.);e:埃伦桥弯藻(Cymbella ehrenbergii);f:尖头舟形藻(Navicula cuspidate);g:颗粒直链藻(Melosira granulata);h:针状蓝纤维藻(Dactylococcopsis acicularis);i:拟多甲藻孢囊(Peridiniopsis Lemmermann);j:四尾栅藻(Scenedesmus quadricauda)k:链丝藻(Hormidium flaccidum);l:小球藻(Chlorella vulgaris),Scales: 10 μm) Fig. 6 The morphology of microalgae in surface sediment in Jiangdong Reservoir |
上覆水中甲藻丰度和沉积物Fe/Al结合态磷含量呈负相关,相关系数为-0.529(p<0.05,n=16),表明沉积物中的Fe/Al结合态磷释放的活性磷可以供给甲藻生长.同时,上覆水甲藻丰度和沉积物中Ca-P含量呈显著负相关,可能的原因是Ca2+对甲藻的增殖和甲藻孢囊的形成有重要作用,Ca-P的生成减少了活性的Ca2+,相应地影响了甲藻细胞的增殖(Tsim et al.,1997).另外,甲藻孢囊丰度与沉积物隐藻丰度显著正相关,有研究发现,水体中甲藻可以吞噬隐藻作为营养补充或者食物(张清春等,2012),两者之间的种间关系有待进一步深入研究.
4 结论(Conclusions)1) 2011年九龙江北溪江东库区上覆水温变化范围为14.5~29.5 ℃,溶解氧含量为4.0~10.0 mg·L-1,全年上覆水中浮游植物生物量在105 ~106 cells·L-1以上.上覆水中不同季节藻类以硅藻门、绿藻门、隐藻门、裸藻门、甲藻门、蓝藻门为主.4月的藻类细胞丰度为全年最高,隐藻门和裸藻门分别占水体藻类的17.61%~37.83%和29.24%~42.29%;6月蓝藻门占水体浮游植物的32.2%~63.89%;9月绿藻门和裸藻门分别占水体浮游植物的10.00%~40.00%、23.85%~49.02%;12月藻类丰度为105 cells·L-1,为全年最低,多样性比较好.
2) 该区域表层沉积物平均粒径在4~10 μm之间,大部分为粉砂质粘壤土.沉积物总碳含量为9140.2 ~17760.0 mg·kg-1,总氮含量为950.0~1600.4 mg·kg-1,总磷含量为483.7~856.2 mg·kg-1.该区域各断面表层沉积物微藻细胞丰度范围在7.41×105~2.17×106 cells·g-1(以底泥干重计)之间,主要为硅藻门、绿藻门,分别占微藻的45.52%~86.42%、5.96%~35.55%.
3) 统计分析结果显示,沉积物微藻丰度与上覆水体浮游植物呈显著负相关,反映了沉积物微藻对上覆水体浮游植物的贡献.沉积物中蓝藻、隐藻、甲藻孢囊丰度分别和上覆水体水温呈显著负相关,分别与溶解氧呈显著正相关,与12月沉积物中的微藻细胞丰度明显高于其他月份相符.沉积物微藻丰度和沉积物含水率、粉粒分别呈显著正相关,反映了沉积物微藻沿着水流方向的积累.
| [1] | Anderson D M, Stock CA, Keafer B A, et al. 2005.Alexandrium fundyense cyst dynamics in the Gulf of Maine[J]. Deep-Sea Research II, 52 (19/21): 2522–2542. |
| [2] | Bostr m B, Petterson A K, Ahlgren I. 1989.Seasonal dynamics of a cyanobacteria-dominated microbial community in surface sediments of a shallow,eutrophic lake[J]. Aquatic Sciences, 51 : 153–178. |
| [3] | Brunberg A K, Blomqvist P. 2002.Benthic overwintering of Mycrocystis colonies under different environmental conditions[J]. Plankton Research, 24 : 1247–1252. |
| [4] | 付永虎.2009.典型海域沉积物中浮游植物休眠体及其萌发研究[D].广州:暨南大学 |
| [5] | 黄清辉, 王磊, 王子健.2006.中国湖泊水域中磷形态转化及其潜在生态效应研究动态[J].湖泊科学, 18 (3):199–206. |
| [6] | Ishikawa A, Hattori M, Ishii K I, et al. 2014..In situ dynamics of cyst and vegetative cell populations of the toxic dinoflagellate Alexandrium catenella in Ago Bay, central Japan[J]. Journal of Plankton Research, 36 (5): 1333–1343. |
| [7] | Jolanda M H, Eveline O F M, Petra M, et al. 2005.Benthic-pelagic coupling in the population dynamics of the harmful cyanobacterium Microcystis[J]. Freshwater Biology, 50 : 854–867. |
| [8] | 姜霞, 王书航, 钟立香.2010.巢湖藻类生物量季节性变化特征[J].环境科学, 31 (9):2056–2062. |
| [9] | Hou D K, He J, Lu H W, et al. 2014.Spatial variations and distributions of phosphorus and nitrogen in bottom sediments from a typical north-temperate lake, China[J]. Environ Earth Sci, 71 : 3063–3079. |
| [10] | 鲁婷, 陈能汪, 陈朱虹, 等.2013.九龙江河流-库区系统沉积物磷特征及其生态学意义[J].环境科学, 34 (9):2430–3436. |
| [11] | 刘培怡.2011.黄河上游沉积物中磷形态分布及其对磷的吸附特征研究[D].呼和浩特:内蒙古师范大学 |
| [12] | Matsuoka K, Fukuyo Y. 2000. Technical guide for modern Dinoflagellate Cyst study[M]. Tokyo: Japan Society for the Promotion of Science . |
| [13] | Mohamed Z A, Al-Shehri A M. 2011.Occurrence and germination of dinoflagellate cysts in surface sediments from the Red Sea off the coasts of Saudi Arabia[J]. Oceanologia, 53 (1): 121–136. |
| [14] | Moore S K, Bill B D, Hay L R, et al. 2015.Factors regulating excystment of Alexandrium in Puget Sound, WA, USA[J]. .Harmful Algae, 43 : 103–110. |
| [15] | Persson A. 2001.Proliferation of cryptic protists and germination of resting stages from untreated sediment samples with emphasis on dinoflagellates[J]. Ophelia, 55 (3): 151–166. |
| [16] | Preston T, Stewart W D P, Reynolds C S. 1980.Bloom-forming cyanobacterium Microcystis aeruginosa overwinters on sediment surface[J]. Nature, 1288 : 365–367. |
| [17] | 全国农业技术推广服务中心. 2006. 土壤分析技术规范[M]. 第2版. 北京: 中国农业出版社 . |
| [18] | Reynolds C S. 1984.The Ecology of Freshwater Phytoplankton[J]. New York:Cambridge University Press, : 384. |
| [19] | Sildever S, Andersen T J, Ribeiro S, et al. 2015.Influence of surface salinity gradient on dinoflagellate cyst community structure, abundance and morphology in the Baltic Sea, Kattegat and Skagerrak.Estuarine[J]. Coastal and Shelf Science, 155 : 1–7. |
| [20] | Tett P.2004.Detecting undesirable disturbance in the context of eutrophication marine ecosystems.Developing management, monitoring and indicators to deliver the ecosystem approach in the coastal and marine environment[C].A Coast NET Conference.London: 24-41 |
| [21] | Tsim S T, Wong J T, Wong Y H. 1997.Calcium ion dependency and the role of inositol phosphates in melatonin-induced encystment of dinoflagellates[J]. Journal of cell science, 110 (12): 1387–1393. |
| [22] | 吴天灵.2009.大亚湾海域休眠期浮游植物的萌发及其与赤潮形成之间的关系[D].广州:华南师范大学 |
| [23] | 万能, 汤俊, 宋立荣.2010.微囊藻休眠体复苏机制的研究进展[J].水生生物学报, 3 (4):113–117. |
| [24] | 王朝晖, 康伟.2014.柘林湾表层沉积物中甲藻孢囊的分布与浮游植物休眠体萌发研究[J].环境科学学报, 34 (8):2043–2050. |
| [25] | 肖兴复, 李文奇, 刘娜.2005.富营养化水体中蓝藻毒素的危害及其控制[J].中国水利水电科学研究院学报, 3 (2):116–123. |
| [26] | 阎荣, 孔繁翔, 韩小波.2004.太湖底泥表层越冬藻类群落动态的荧光分析法初步研究[J].湖泊科学, 16 (2):163–168. |
| [27] | 赵汉取, 韦肖杭, 姚伟忠.2009.蓝藻爆发后南太湖水域浮游生物及富营养化[J].浙江海洋学院学报:自然科学版, 28 (1):21–25. |
| [28] | 张琪, 缪荣丽, 刘国祥.2012.淡水甲藻水华研究综述[J].水生生物学报, 36 (2):352–359. |
| [29] | 张清春, 于仁成, 宋静静, 等.2012.几株赤潮甲藻的摄食能力[J].生态学报, 32 (2):402–413. |
| [30] | 周亮进, 由文辉.2008.福建省山仔水库富营养化特征[J].水资源保护, 24 (2):26–29. |