 2016, Vol. 36
2016, Vol. 36



2. 无锡科技职业学院, 无锡 214122
2. Wuxi Professional College of Science and Technology, Wuxi 214122
Corresponding author, E-mail: liuhe@jiangnan.edu.cn
据统计,至“十二五”期末我国湿污泥量(含湿量80%)将突破4600万t(王锋等,2011),而污泥厌氧消化技术以其低能耗、高产出的经济优势成为污泥资源化利用的主要技术之一.除厌氧消化产甲烷以外,污泥产挥发性脂肪酸(VFA)也是实现污泥资源化的有效途径,近年来,越来越多的学者开始关注污泥厌氧发酵产挥发性脂肪酸(戴晓虎,2012; Pilli et al., 2015).目前,有关污泥厌氧发酵产酸的研究主要集中在通过改进装置构型(王田田,2013)、产酸微生物生态(王晋,2013)、优化控制运行条件,如控温(Mottet et al., 2009; Feng et al., 2009)、pH(Wang et al., 2014; Liu et al., 2013)等条件因素来提高产酸效率.已有研究表明,通过调节发酵污泥底物的C/N比可增加发酵产酸量并调控其产酸类型(任南琪等,2005;刘晓玲,2008),然而,目前研究人员对污泥厌氧发酵产酸过程中不同C/N比与关键酶酶活及有机酸产酸量间的关系并不清楚.仅有为数不多的研究,如优化C/N比条件作为酒精发酵的实验模型研究(Manikandan et al., 2010),而对于数学模型则没有报道.数学模型法作为现代科学研究的重要手段,它有助于描述和理解生物处理系统的反应过程,可为工程设计提供理论上的指导;还有助于工艺的优化和控制,从而更好地指导实际生产运行.
多元线性回归是一种理想的描述多个因素之间关系的数学方法,能较好地确定被解释变量和解释变量之间的关系,在很多领域得到了应用(常盛等,2011).因此,本研究通过设置不同C/N比条件来调控污泥厌氧发酵产酸,在Matlab7.0平台上建立多元线性回归函数模型,拟合C/N比、关键酶酶活和产酸类型之间的关系,以期为今后污泥发酵产酸条件调控研究和工程放大提供参考.
2 材料与方法(Materials and methods) 2.1 实验材料 2.1.1 污泥与种泥原始污泥取自无锡市太湖新城污水处理厂,发酵底物是经过热碱预处理的污泥上清液.污泥采集后置于阴凉处,风干10 d,采用机械粉碎仪粉碎,再过30目筛,密封置于-15 ℃冰柜中保存.
接种污泥来源于无锡某柠檬酸厂上流式厌氧污泥反应器(UASB)中的厌氧颗粒污泥.在100 ℃下煮沸2 h以杀死产甲烷菌(Logan et al., 2002),然后导入有效容积为2 L的UASB中进行驯化,活化种泥中的产酸菌(郭磊等,2008),驯化温度为35 ℃.每日监测驯化种泥的pH值,待种泥驯化后出水pH值降低至4.0左右,稳定3~5 d后,认为种泥驯化成功.原始污泥和污泥预处理液及接种种泥的性质见表 1.
| 表 1 原始污泥、污泥预处理液和接种污泥的性质 Table 1 Characteristics of the original sludge, the pretreated sludge liquor and the seed sludge |
调节热-碱预处理后离心液pH为10.0,取500 mL离心液置于1000 mL的厌氧反应瓶中,分别加入不同量葡萄糖,以使得底物混合液的初始C/N比为12、56、69、156.接入驯化后的种泥,种泥接种量为10%(种泥和待处理水的体积比).污泥发酵前充氮气10 min以去除氧气,然后迅速密封置于转速为120 r·min-1和温度(35±1)℃的摇床厌氧发酵.在发酵期间,每12 h调节pH为初始的10.0(Logan et al., 2002).每24 h取样1次,用针管吸出部分发酵液,取样完成后调节pH并充氮气保持厌氧.
2.1.3 实验药品和材料本实验采用的药品包括4-甲基戊酸(0.83 g·L-1)、磷酸溶液(3 mol·L-1)、NaOH(3 mol·L-1)、HCL(3 mol·L-1)等;主要仪器包括pH计(Mettler Toledo,Switzerland)、气相色谱仪(GC-2010 Shimadzu Corporation,Tokyo,Japan)、马弗炉、凯氏定氮仪(Buchi,Switzerland),离心机(Eppendorff,Germany)等.
2.2 试验方法 2.2.1 污泥初始指标测定污泥预处理前后的总固体(TS)、溶解性固体(SCOD)及污泥挥发性固体(VS)的测定采用重量法(Bligh et al., 1959),具体操作详见GB11901-89和《水和废水监测分析方法》.pH值测定参照国标法.
污泥中的总脂类物质采用Bligh-Dyer方法提取后,在 80 ℃下干燥直至溶剂完全挥发后,采用重量法测定(Bligh et al., 1959).总氮采用凯氏定氮法测定.总蛋白含量通过凯氏氮减去氨氮后再乘以 6.25计算得到(Miron et al., 2000).氨氮采用纳氏试剂比色法测定,污泥中的总碳水化合物采用甲醛离心法提取后(Aquino et al., 2004),再用苯酚-硫酸法测定(Dubois et al., 1956).用 Liquid TOC 分析仪测定总有机碳,详见《水质总有机碳的测定燃烧氧化-非分散红外吸收法》(HJ/T71-2001).污泥中的总磷含量用钼酸铵分光光度法测定,详见《水质总磷的测定钼酸铵分光光度法》(GB11893-89).
2.2.2 关键酶活测定乙酸激酶(AK)的活性采用文献(Rose,1955)的方法提取并测定.磷酸转移乙酰酶(PTA)的提取方法同乙酸激酶,活性测定参照文献(Andersch et al., 1983)方法.丁酸激酶(BK)微生物细胞的破壁方法和提取方法同乙酸激酶,活性测定采用文献(Zhu et al., 2003)方法.磷酸转移丁酰酶(PTB)微生物细胞的破壁和提取方法同乙酸激酶,活性测定采用文献(Zhu et al., 2003)方法. 甲基丙二酰CoA变位酶(MCM)活性测定采用文献(Kellermeyer et al., 1969)方法.
2.2.3 挥发性短链脂肪酸的测定采用GC法检测挥发性短链脂肪酸的质量浓度,样品处理及色谱条件等参见文献(Liu et al., 2008).为方便不同条件下产酸效率的比较,将测得的VFAs浓度折算成 COD值,换算方法参见文献(Liu et al., 2008).
3 结果与讨论(Results and discussion) 3.1 有机酸浓度的变化分别设定底物初始C/N比值为12、56、156,进行厌氧发酵.发酵过程中时,体系中有机酸的产量分别如图 1所示.由图 1可以看出,底物初始C/N比不同,厌氧发酵产生的末端酸化产物也不同.C/N比为12时,产量最大的为乙酸,在第5 d达到9.45 kg·m-3(以COD计,下同);其次是丙酸,第5 d时可以达到3.55 kg·m-3;最低是丁酸,第5 d时产量约为2.35 kg·m-3(图 2a).当C/N比为56时,产量最大的为丙酸,在第5 d可达到10.36 kg·m-3;其次是乙酸,可以达到7.79 kg·m-3;最低是丁酸,产量约为2.79 kg·m-3(图 2b).当C/N比为156时,产量最大的为丁酸,在第5 d达到13.59 kg·m-3;其次是乙酸,可以达到5.89 kg·m-3;最低是丁酸,产量约为4.72 kg·m-3(图 2c).
 |
| 图 1 C/N比对发酵产酸的影响(a.C/N=12,b.C/N=56,c. C/N=156) Fig. 1 VFAs at different C/N ratio in fermentation(a.C/N=12,b.C/N=56,c. C/N=156) |
 |
| 图 2 底物发酵产酸的代谢途径 Fig. 2 Metabolic pathways and key enzymes for organic acids production |
在发酵过程中,通过设定不同C/N比条件,测定不同C/N比下关键酶的酶活和产酸量,则三者可以建立函数关系.短链脂肪酸的生成途径如图 2所示(Feng et al., 2009),乙酸生成的关键酶分别为乙酸激酶(AK)和磷酸转移乙酰酶(PTA),丁酸合成的关键酶有丁酸激酶(BK)和磷酸转移丁酰酶(PTB);丙酮酸转化为乙酸过程中,关键酶为甲基丙二酰CoA变位酶(MCM).由于底物C/N比的改变会导致产酸微生物体内的酶活性改变,从而改变微生物不同代谢途径的代谢通量,并最终导致各种短链脂肪酸的生成受到影响,由此产生了不同的产酸类型.因此,C/N比是自变量,而关键酶活性和产酸类型是因变量.
根据以上理论分析,为了建立C/N比、关键酶活、不同酸产量之间的函数关系模型,采用多元非线性回归(霍倩等,2002)的方法,建立了二元二次多项式模型(1)、二元三次多项式模型(2)、二元三次多项式模型(3).
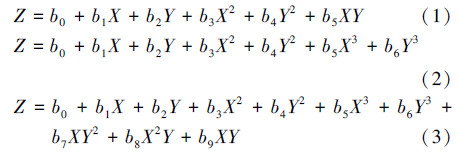
式中,Z为因变量,表示有机酸产量,X和Y为自变量,分别表示C/N比和关键酶活性,bi表示函数中的常数.根据实测数据对以上3种多项式模型进行拟合优度的检验,结果如表 2所示.
| 表 2 模型拟合优度分析 Table 2 Analysis of goodness of model fitting |
表 2中,A、B、C分别表示3种关系式模型,乙酸-AK、乙酸-PTA等分别表示各种产物酸与其对应的关键酶.根据优度拟合理论,R2大于 0.9的较好,因此,在多项式A乙酸-AK、B乙酸-AK和C乙酸-AK的R2检验中,C乙酸-AK效果最好,为0.9327.同样的,C乙酸-PTA、C丙酸-MCM、C丁酸-BK、C丁酸-PTB的R2分别为0.9348、0.9494、0.9880和0.9771,均具有最高的R2.模型拟合优度检验同时要求模型残差平方和越小越好,因此,在多项式A乙酸-AK、B乙酸-AK和C乙酸-AK的残差平方和检验中,C乙酸-AK的效果最好,为0.0292.同样的,C乙酸-PTA、C丙酸-MCM、C丁酸-BK、C丁酸-PTB的残差平方和分别为0.0076、1.8556、0.0038和3.1362,均具有最小的残差平方和.综合以上分析,可以判定二元三次多项式C模型拟合度最好.依据二元三次多项式C标准结构,结合实验测得的不同C/N比条件下的关键酶活和各种有机酸产量,建立了可视化的三者之间的曲面模型.
3.3 C/N比-关键酶-乙酸曲面模型的建立根据上述多项式C的形式,利用Matlab软件进行拟合,得到多项式当中相应bi的值,进而得出C/N比-关键酶AK-乙酸之间的定量关系表达式(4)和C/N比-关键酶PTA-乙酸之间的定量关系表达式(5).

根据定量关系式(4)和(5)绘制得到图 3的可视化曲面模型.从图 3的曲面模型可以看出:在初始C/N比为10~50时,酶活性较低,乙酸激酶(AK)和磷酸转乙酰酶(PTA)平均酶活性分别为1.06 U·mg-1和0.67 U·mg-1,而乙酸浓度从2.8 kg·m-3(以COD计,下同)上升到7.8 kg·m-3.在初始C/N比为50~150时,AK和PTA酶活性较高,平均值分别为2.79 U·mg-1和1.08 U·mg-1,而乙酸浓度为2.6 kg·m-3.从以上数据可知,酶活性水平和乙酸产量不一致,说明在此C/N比条件下,乙酸主要不是通过丙酮酸途径合成,而可能是通过丙酸、丁酸的转化形成.其他学者的研究都表明(任南琪等,2005;刘晓玲,2008),C/N比通过直接或间接影响产能过程及NADH(或NADPH)/NAD+(或 NADP+)的氧化还原偶联过程,促使不同发酵产酸类型的形成.在低C/N比条件下,乙酸的产生主要是通过氨基酸之间的Stickland反应形成,本文的曲面模型很好地解释了这一结论.
 |
| 图 3 C/N比、产乙酸关键酶和不同酸产量之间的曲面模型(a.乙酸激酶(AK),b.磷酸转移乙酰酶(PTA)) Fig. 3 Model of C/N ratio,acetic key enzyme and acid production (a.Acetate Kinase,b.Acetyl Phosphate) |
C/N比-关键酶MCM-丙酸之间的定量关系表达式如式(6)所示.由C/N比-关键酶MCM-丙酸曲面模型可以看出,当 C/N比为 55~70时,甲基丙二酰CoA变位酶(MCM)活性都有大幅度增加,同时丙酸的产量也开始上升,这与任南琪等(2005)的研究一致,表明丙酸型发酵中,丙酸产生于糖酵解丙酮酸途径.根据已有的理论研究,通常情况下,丙酸型发酵代谢途径有利于NADH+H+的氧化,而丁酸型发酵缺乏对 NADH 的再生能力.所以当 C/N 值处于56~69(王勇等,2004;任南琪等,2005)时,微生物的细胞合成速率较小,丙酸型发酵比丁酸型发酵有更高的稳定性,产酸结果就形成丙酸型发酵,这也解释了本文曲面模型当中丙酮酸的产量和关键酶活性大幅提升的原因.

C/N比-关键酶BK-丁酸之间的定量关系如式(7)所示,C/N比-关键酶PTB-丁酸之间的定量关系如式(8)所示.
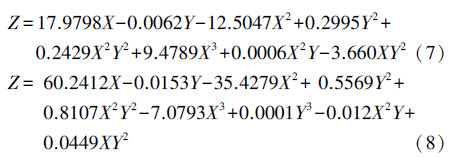
在初始C/N比为150及更高时,丁酸激酶(BK)和磷酸转移丁酰酶(PTB)的活性均有大幅度增加.以前的研究表明,丁酸的产生主要由糖酵解丙酮酸形成(任南琪等,2005).随着C/N值升高到156,碳作为微生物细胞的重要组成元素,促使了细胞合成代谢速率的提高,有机质分解代谢过程中产生的部分NADH(或NADPH)能够被厌氧微生物迅速地用于细胞合成而得以再生,所以,呈现较稳定的丁酸型发酵类型.
已有研究表明,通过调控污泥预处理液的初始 C/N 值可实现乙酸、丙酸和丁酸不同厌氧发酵类型之间的转变.在初始C/N比为10~50时,此时发酵是乙酸型发酵.而当 C/N 值为 55~70时,此时厌氧发酵类型则转变为丙酸型.在初始C/N比为150及更高时,此时,丙酸型发酵类型则转变为丁酸型(任南琪等,2005).本文图 3~5建立的曲面模型结果表明,所建立的模型能很好地反映这一趋势,说明模型能较好地模拟C/N比调控下的污泥发酵产酸实验结果.
 |
| 图 4 C/N比、丙酸关键酶-甲基丙二酰CoA变位酶和不同酸产量之间的曲面模型 Fig. 4 Model of C/N ratio, propionate key enzyme-Methylmalonyl-CoA mutase and propionate production |
 |
| 图 5 C/N比、丁酸关键酶活和不同酸产量之间的曲面模型(a.丁酸激酶(BK),b.磷酸转乙酰酶(PTB)) Fig. 5 Model of C/N ratio,key enzyme and butyrate production(a.Butyrate Kinase,b.Phosphate Butryltransferase) |
1)采用多元非线性回归的方法建立了不同C/N比、关键酶活和酸产量之间的函数关系模型.经拟合优度、显著性和残差平方和等指标的比较,发现二元三次多项式模型的拟合优度系数和显著性高、残差平方和较小,表明该模型能较好地拟合三因素之间的定量关系.
2)调节污泥水解液的C/N比分别为12、56、156,将测定的关键酶酶活和乙酸、丙酸、丁酸的结果代入建立的多项式模型,应用Matlab 7.0 软件得到了相应的定量关系式和可视化的曲面模型,曲面模型的结果很好地验证了不同C/N比条件下的产酸代谢途径.
| [1] | Andersch W, Bahl H, Gottschalk G.1983.Level of enzymes involved in acetate, butyrate, acetone and butanol formation by Clostridium acetobutylicum[J].European Journal of Applied Microbiology and Biotechnology, 18(6):327-332 |
| [2] | Aquino S F, Stuckey D C.2004.Soluble microbial products formation in anaerobic chemostats in the presence of toxic compounds[J].Water Research, 38(2):255-266 |
| [3] | Bligh E G, Dyer W J.1959.A rapid method of total lipid extraction and purification[J].Canadian Journal of Biochemistry and Physiology, 37(8):911-917 |
| [4] | 常盛, 朱亚玲.2011.基于SPSS的多元线性回归算法建模的实例研究[J].数字技术与应用, (10):120 |
| [5] | 戴晓虎.2012.我国城镇污泥处理处置现状及思考[J].给水排水, 38(2):451-50 |
| [6] | Dubois M, Gilles K A, Hamilton J K, et al.1956.Colorimetric method for determination of sugars and related substances[J].Analytical Chemistry, 28(3):350-356 |
| [7] | Feng L Y, Wang H, Chen Y G, et al.2009.Effect of solids retention time and temperature on waste activated sludge hydrolysis and short-chain fatty acids accumulation under alkaline conditions in continuous-flow reactors[J].Bioresource Technology, 100(1):44-49 |
| [8] | Feng L Y, Chen Y G, Zheng X.2009.Enhancement of waste activated sludge protein conversion and volatile fatty acids accumulation during waste activated sludge anaerobic fermentation by carbohydrate substrate addition:the effect of pH[J].Environmental Science & Technology, 43(12):4373-4380 |
| [9] | 郭磊, 刘和, 李秀芬, 等.2008.多级逆流工艺促进城市污泥厌氧发酵生产挥发性脂肪酸[J].生物工程学报, 24(7):1233-1239 |
| [10] | 霍倩, 李书全, 王文元, 等.2002.遗传算法应用于多元非线性回归模型求参的研究[J].河北农业大学学报, 25(2):107-110 |
| [11] | Kellermeyer R W, Wood H G.1969.2-methylmalonyl-CoA mutase from Propionibacterium shermanii (methylmalonyl-CoA isomerase)[J].Methods in Enzymology, 13:207-215 |
| [12] | Liu X G, Dong B, Dai X H.2013.Hydrolysis and acidification of dewatered sludge under mesophilic, thermophilic and extreme thermophilic conditions:Effect of pH[J].Bioresource Technology, 148:461-466 |
| [13] | 刘晓玲.2008.城市污泥厌氧发酵产酸条件优化及其机理研究[D].无锡:江南大学 |
| [14] | Liu X L, Liu H, Chen Y Y, et al.2008.Effects of organic matter and initial carbon-nitrogen ratio on the bioconversion of volatile fatty acids from sewage sludge[J].Journal of Chemical Technology and Biotechnology, 83(7):1049-1055 |
| [15] | Logan B E, Oh S E, Kim I S, et al.2002.Biological hydrogen production measured in batch anaerobic respirometers[J].Environmental Science & Technology, 36(11):2530-2535 |
| [16] | Manikandan K, Viruthagiri T.2010.Optimization of C/N ratio of the medium and fermentation conditions of ethanol production from tapioca starch using co-culture of Aspergillus niger and Sachormyces cerevisiae[J].International Journal of ChemTech Research, 2(2):947-955 |
| [17] | Miron Y, Zeeman G, Van Lier J B, et al.2000.The role of sludge retention time in the hydrolysis and acidification of lipids, carbohydrates and proteins during digestion of primary sludge in CSTR systems[J].Water Research, 34(5):1705-1713 |
| [18] | Mottet A, Steyer J P, Déléris S, et al.2009.Kinetics of thermophilic batch anaerobic digestion of thermal hydrolysed waste activated sludge[J].Biochemical Engineering Journal, 46(2):169-175 |
| [19] | Pilli S, Yan S, Tyagi R D, et al.2015.Thermal pretreatment of sewage sludge to enhance anaerobic digestion:a review[J].Critcal Reviews in Environmental Science and Technology, 45(6):669-702 |
| [20] | 任南琪, 王爱杰, 马放.2005.产酸发酵微生物生理生态学[M].北京:科学出版社.51-135 |
| [21] | Rose I.1955.Acetate kinase of bacteria (acetokinase)[J].Methods in Enzymology, 1:591-595 |
| [22] | 王锋, 卢雪丽, 陈杰, 等.2011.城市生活污水污泥减量化和资源化研究概述[J].北方环境, 23(10):153-157 |
| [23] | 王晋.2013.厌氧发酵产酸微生物种群生态及互营关系研究[D].无锡:江南大学 |
| [24] | Wang K, Yin J, Shen D S, et al.2014.Anaerobic digestion of food waste for volatile fatty acids (VFAs) production with different types of inoculum:Effect of pH[J].Bioresource Technology, 161:395-401 |
| [25] | 王田田.2013.高含固率城市污泥厌氧消化工艺研究[D].济南:山东建筑大学 |
| [26] | 王勇, 孙寓姣, 任南琪, 等.2004.C/N对细菌产氢发酵类型及产氢能力的影响[J].太阳能学报, 25(3):375-378 |
| [27] | Zhu Y, Yang S T.2003.Adaptation of Clostridium tyrobutyricum for enhanced tolerance to butyric acid in a fibrous-bed bioreactor[J].Biotechnology Progress, 19(2):365-372 |