 2015, Vol. 35
2015, Vol. 35



2. 钦州学院, 广西北部湾海洋生物多样性养护重点实验, 钦州 535099;
3. 广西民族大学化学化工学院, 南宁 530006
2. Guangxi Key Laboratory of Beibu Gulf Marine Biodiversity Concervation, Qinzhou University, Qinzhou 535099;
3. School of Chemistry and Chemical Engineering, Guangxi University of Nationalities, Nanning 530006
浮游植物作为海洋生态系统中的初级生产者,在生态系统的能量流动和物质循环中起着重要作用(Sathyendranath et al., 1991; Lalli et al., 1993).叶绿素a(Chl-a)是海洋中浮游植物现存量的一个良好指标(孙晓霞等,2011),在海洋调查研究中占据重要的地位(文斐等,2012).初级生产力反映了海洋浮游植物通过光合作用生产有机碳的能力,其变化在全球碳循环和海洋生物资源方面起着重要作用(Sarmiento et al., 1988; 蔡玉婷,2010).因此,开展Chl-a和初级生产力的调查,对海洋生态系统和全球气候变化的研究及海洋渔业、水产养殖的发展具有重要的科学意义(Valdéz-Holguín et al., 1987; Gong et al., 2000; Song et al., 2004).关于近海河口海湾Chl-a与初级生产力时空变化方面的研究,目前国际上已开展了大量的调查工作(Harding,1994; Lohrenzl et al., 1997; Oviatt et al., 2002; Glé et al., 2008).这些研究表明,很多河口海湾区域Chl-a浓度和初级生产力水平与营养盐输入关系密切.
钦州湾位于北部湾顶端,主要由内湾和外湾构成,中间狭小,两端开阔,呈哑铃状,海湾三面环陆,只有南面与北部湾相通,海岸线绵延520多km,水域总面积约为380 km2,平均水深约为5.2 m,潮汐类型属不规则全日潮.钦州湾周边主要有钦江、茅岭江和大风江等入海河流,地表径流的大量输入为海洋生物提供了充足的营养物质,使得该海湾鱼类资源丰富,水产养殖业发达(薛保铭等,2013).特别是近年来随着广西北部湾经济区的开放开发,钦州湾作为重点开发建设的海湾之一,现已发展成为我国南方重要的天然渔场和人工养殖基地(韦蔓新等,2001).例如,目前海湾沿岸滩涂插养及深水吊养的近江牡蛎面积已达14万多亩,而可供开发养殖的滩涂有130多万亩.为了保证这一海湾“天然牧场”的可持续发展和渔业经济效益的产出,已有学者对其水体中的营养盐(杨斌等,2012a; 2013a; 2013b)、浮游生物(蓝文陆等,2011; 王迪等,2013)、COD(杨斌等,2014a; 2014b)和重金属(韦蔓新等,2004;杨斌等,2012b)等做了相关研究,但目前有关钦州湾Chl-a和初级生产力时空变化及其影响因素方面的研究却鲜有报道.
海洋浮游植物及其初级生产过程在海洋生态系统中成为重要的驱动因子之一,其动态变化会对生态系统结构和功能产生直接影响(孙松等,2005).Chl-a浓度变化趋势能够反映海域浮游植物的现存量及其变化规律,是估算海洋浮游植物生物量及初级生产力的重要指标(李超伦等,2004; 周伟华等,2004).因此,开展Chl-a和初级生产力的调查,对于评价海湾生态环境状况及评估海湾环境容量具有重要的科学指导意义.基于此,本文通过2009年春、夏、秋、冬4个季度的月调查,研究钦州湾Chl-a和初级生产力的时空变化特征及其影响因素,以期全面了解钦州湾养殖海区的生态环境状况及海洋生态系统结构和功能,为该湾海洋生物资源的可持续利用、环境污染治理及养殖环境容量的评估提供科学依据.
2 材料与方法(Material and methods) 2.1 采样站位2009年1月(冬季)、4月(春季)、7月(夏季)和11月(秋季),在钦州湾108.49°~108.86°E和21.56°~21.87°N范围内海区分别进行了4个航次Chl-a的海洋环境因子的调查,共设了15个采样站位(图 1).其中,内湾(茅尾海)布设1~6号站位,外湾(钦州港海区)布设7~11号站位,湾外(三娘湾海区)布设12~15号站位.
 |
| 图 1 钦州湾调查海区采样站位 Fig. 1 Sampling sites in Qinzhou Bay |
现场样品的采集与保存均按照《海洋调查规范》(GB/T 12763.4—2007)进行.各站点表层(0.5 m)水样使用Niskin采水器采集,Chl-a水样先用200 μm孔径的筛绢滤去浮游动物,再用0.7 μm(GF/F,Whatman)滤膜进行减压过滤,滤膜用铝箔包裹,置于-20 ℃冰箱内冷冻保存待测.测定时在低温(4 ℃)、避光条件下,将滤膜浸泡于10 mL 90%的丙酮中超声处理10 min后萃取24 h,在实验室内取上清液用F-4500荧光分光光度计(日本日立公司生产)测定(Parsons et al., 1984).现场采样站位的水温(T)、盐度(S)、pH和溶解氧(DO)参数应用YSI6920多功能水质分析仪(美国YSI公司生产)现场测定.5项营养盐(NO-2、NO3-、NH+4、PO3-4和SiO2-3)测定均按照《海洋监测规范》(GB17378.4—2007)中的方法进行,营养盐水样经0.45 μm醋酸纤维滤膜过滤后,使用分光光度法进行分析.化学需氧量(COD)通过碱性高锰酸钾法进行分析.
2.3 初级生产力估算为了解该海区初级生产力的现状,根据水深和真光层的深度,在较有代表性的6、8和15号站进行初级生产力的测定,按100%、50%和10%光衰减深度采集3层水样.先用200 μm孔径的筛绢滤去浮游动物,倒入50 mL KIMAX培养管中(Song et al., 2004),设置二白一黑的瓶子,并分别加入14C示踪液,采用中性衰光材料控制光照强度培养4 h以上.通过现场透明度盘测量透明度并对光衰减度进行估算(李武等,1993).培养后的水样用0.7 μm(GF/F,Whatman)滤膜过滤,将样品带回实验室,使用LS6500型多功能液体闪烁计数器(美国BACKMAN公司)测定样品中14C的放射性强度,计算碳同化系数(吕瑞华,1993)的公式如下:

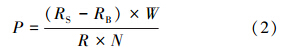
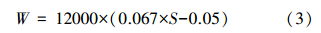
水柱同化系数的算术平均值的计算公式如下:
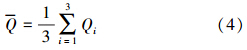
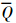 为水柱中同化系数平均值(mg · mg-1 · h-1),Qi为第i层的同化系数(mg · mg-1 · h-1).本研究中 结果介于1.4~6.2 mg · mg-1 · h-1之间,平均值为3.6 mg · mg-1 · h-1.因碳同化数现场测定受环境条件诸多因素影响变化较大,且测定的同化系数与近岸水域平均同化系数(3.7 mg · mg-1 · h-1)相近(Ryther,1969;孙军等,2003; 叶宁等,2013),同时为与其他海湾调查资料对比,本文仍采用同化系数3.7 mg · mg-1 · h-1对初级生产力进行估算,具体依据Cadée(1975)提出的简化公式:
为水柱中同化系数平均值(mg · mg-1 · h-1),Qi为第i层的同化系数(mg · mg-1 · h-1).本研究中 结果介于1.4~6.2 mg · mg-1 · h-1之间,平均值为3.6 mg · mg-1 · h-1.因碳同化数现场测定受环境条件诸多因素影响变化较大,且测定的同化系数与近岸水域平均同化系数(3.7 mg · mg-1 · h-1)相近(Ryther,1969;孙军等,2003; 叶宁等,2013),同时为与其他海湾调查资料对比,本文仍采用同化系数3.7 mg · mg-1 · h-1对初级生产力进行估算,具体依据Cadée(1975)提出的简化公式:
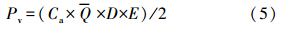
为水柱碳同化系数算术平均值(mg · mg-1 · h-1),D为昼长时间(h),E为真光层深度(m),取透明度的3倍.
2.4 数据处理
数据采用SPSS19.0软件进行分析,Chl-a与各环境因子的相互关系采用Pearson相关性分析,显著性水平α=0.05,本文中数据均以算术平均值±标准偏差(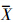 ± SD)表示.
± SD)表示.
由图 2可见,钦州湾Chl-a浓度周年变化在0.83~32.5 mg · m-3之间,平均浓度为5.39 mg · m-3.钦州湾春季(4月)Chl-a浓度范围为1.59~7.12 mg · m-3,高值出现在内湾的5和6号站及三娘湾海区的12、13和14号站,钦州港海区站位浓度较低.夏季(7月)Chl-a浓度范围为1.58~32.5 mg · m-3,各站位之间变化幅度较大,空间分布由内湾向三娘湾海区呈现高-低-高的变化趋势,最高值出现在三娘湾海区的14号站,最低值出现在钦州港海区的10号站.秋季(11月)Chl-a浓度范围为0.83~3.95 mg · m-3,空间分布与春、夏季恰相反,总体呈现由内湾向三娘湾海区呈现低-高-低的变化趋势.冬季(1月)Chl-a浓度范围为2.04~4.25 mg · m-3,各站位之间变化幅度最小,总体分布与春、夏季相一致,呈现内湾高、钦州港海区低和三娘湾海区高的特征.
 |
| 图 2 钦州湾不同站位叶绿素a浓度 Fig. 2 Variation of chlorophyll-a concentration at each station in Qinzhou Bay |
夏季钦州湾Chl-a平均浓度为(12.21±10.93)mg · m-3,是一年四季中的最高峰(图 3);春季和冬季次之,平均浓度分别为(3.42±1.46)mg · m-3和(3.21±0.70)mg · m-3;秋季浓度最低,平均浓度为(2.74±1.18)mg · m-3.夏季和秋季表层海水Chl-a浓度之间差异显著(p<0.0001),可见Chl-a浓度存在显著的季节变化特征,这与王迪等(2013)在2008—2009年对钦州湾浮游植物平均丰度季节调查结果相一致,均表现出夏季>春季>冬季>秋季,反映了该海湾浮游植物现存量由冬季开始,经春季逐渐增大至夏季达到最大值,秋季开始逐渐下降的变化趋势.
 |
| 图 3 钦州湾叶绿素a含量的季节变化 Fig. 3 Seasonal variation of chrolophyll-a in Qinzhou Bay |
由图 4可见,钦州湾初级生产力周年变化平均值(范围)为425.1(92.3~1494.5)mg · m-2 · d-1(以C计,下同).海湾初级生产力有着明显的空间分布特征,各季节的分布状况基本与相应季节Chl-a浓度变化相一致.钦州湾春季(4月)初级生产力为105.9~418.2 mg · m-2 · d-1,各站位之间变化幅度相对较小,高值位于内湾的1、2、5和6号站及三娘湾海区的12和13号站,钦州港海区站位浓度较低.夏季(7月)初级生产力为130.3~1494.5 mg · m-2 · d-1,各站位之间变化幅度较大,表现出内湾、三娘湾海区高,钦州港海区低的变化趋势,最高值出现在三娘湾海区的13号站,最低值出现在钦州港海区的8号站.秋季(11月)初级生产力为92.3~604.3 mg · m-2 · d-1,高值区分布在钦州港海区,但最高值出现在湾颈处6号站,低值区则分布在内湾和三娘湾海区,最低值出现在茅岭江口1号站.冬季(1月)初级生产力为271.7~791.0 mg · m-2 · d-1,空间分布与春、夏季基本一致,总体呈现内湾高、钦州港海区低、三娘湾海区高的分布特征.
 |
| 图 4 钦州湾不同站位初级生产力 Fig. 4 Variation of primary productivity at each station in Qinzhou Bay |
钦州湾初级生产力均值的季节变化如图 5所示,其高峰值出现在夏季(7月),平均值为671.3 mg · m-2 · d-1;冬季(1月)和秋季(11月)次之,平均值分别是515.6 mg · m-2 · d-1和296.2 mg · m-2 · d-1;低谷值出现在春季(4月),均值为217.4 mg · m-2 · d-1.夏季和秋季初级生产力之间呈现显著性差异(p<0.05),可见初级生产力存在显著的季节变化特征.整个海域初级生产力周年变化呈现双峰型,分别在夏季和冬季出现两个高峰,符合亚热带海域一年双峰的普遍现象,这与叶宁等(2013)在2008—2009年对广东流沙湾初级生产力调查结果相一致.
 |
| 图 5 钦州湾初级生产力季节变化 Fig. 5 Seasonal variation of primary productivity in Qinzhou Bay |
为了解钦州湾海域环境因子对浮游植物生长的影响,对钦州湾4个季节Chl-a浓度与同步测定的各项环境因子进行相关性分析,结果如表 1所示.夏季海水温度和pH值与Chl-a分别呈显著负相关(p<0.05)和显著正相关(p<0.01);盐度与Chl-a分别在夏(p<0.01)、秋(p<0.05)季呈显著负相关;而DO与Chl-a分别在夏(p<0.01)、冬(p<0.05)季呈显著正相关.营养盐及其摩尔比值方面,仅夏、秋季NH+4-N与Chl-a呈显著正相关(p<0.05);秋季Si/N比和冬季N/P比与Chl-a分别呈显著负相关和正相关(p< 0.01).
| 表1 钦州湾表层叶绿素a浓度与环境因子的相关系数 Table 1 Correlation coefficient between Chl-a concentrations and environmental factors in Qinzhou Bay |
空间分布上,春、夏、冬季钦州湾Chl-a浓度均在茅尾海和三娘湾海区出现高值区,低值区出现在钦州港海区,秋季出现相反的变化趋势.茅岭江和钦江是茅尾海两大主要入海河流,大风江是三娘湾海区主要入海河流,陆源输入带来了大量的营养物质进入海湾,促进了海洋浮游植物的生长繁殖.钦州湾是广西沿海港湾中平均涨落潮流速最大的海湾,涨潮时流向偏北,落潮时流向偏南,钦州港海区属强潮流区(韦蔓新等,2002),强烈的潮流不利于浮游植物稳定生长繁殖(Zuenko et al., 2006).因此,陆源输入和潮流作用是影响春、夏、冬季Chl-a浓度在内湾和三娘湾海区出现高值、在钦州港海区出现低值的重要因素.秋季正处于茅尾海和三娘湾海区近江牡蛎等贝类养殖生长最快时期,摄取了大量的浮游植物作为饵料.贝类滤食压力远大于浮游动物的摄食压力,对浮游植物生物量起到关键的控制作用,这是造成Chl-a浓度较低的重要原因(Huang et al., 2008;郝林华等,2012),因而秋季出现内湾和三娘湾海区Chl-a浓度低于钦州港海区的现象.由此可见,贝类养殖通过滤食活动已能够影响钦州湾Chl-a浓度的空间分布.
浮游植物现存量不仅受优势种季节演替等生物学因素的影响,还受到各种生物及非生物环境因子方面的影响(Sathyendranath et al., 1991; Reynolds,1994; Agawin et al., 2000).春季,正值春耕春种时节,沿岸入海河流携带大量营养物质进入海湾,而海水温度的上升及光照强度的增强促进浮游植物的大量繁殖(Harding,1994; Glé et al., 2008).钦江和茅岭江夏季入海径流量是春、秋季节的3倍左右,是冬季的6倍左右(中国海湾志编纂委员会,1993).此时水体中丰富的营养盐在强烈光照和最高水温下直接刺激内湾和三娘湾海区优势藻种大量繁殖,出现浮游植物数量最高峰(蓝文陆等,2011;王迪等,2013).尽管7月份正值近江牡蛎幼苗繁殖高峰期,且存在浮游动物对浮游植物的滤食作用,但其摄食压力远低于浮游植物的增长繁殖速度(Huang et al., 2008;郝林华等,2012).因此,浮游植物现存量仍处于一年中的最高峰,故Chl-a浓度最高值出现在夏季.进入秋冬季节,水温下降和光照强度减弱,陆地径流和降雨量不断减少,海水中营养盐的含量相对较低,浮游植物生物量开始下降,Chl-a浓度也相应降低.秋季正处于钦州湾贝类养殖的快速生长期,此时贝类滤食压力对浮游植物现存量起着重要的控制作用(Huang et al., 2008),导致秋季Chl-a浓度低于冬季,这与王迪等(2013)对钦州湾浮游植物丰度及其影响因素的分析结果一致.
4.2 初级生产力时空变化及其影响因素钦州湾初级生产力介于92.3~1494.5 mg · m-2 · d-1 之间,平均为425.1 mg · m-2 · d-1.调查结果在国内外近海海区初级生产力估算范围之内(Menzel et al., 1959~1960;Raymont,1980;Gong et al., 2000;蔡玉婷,2010;李斌等,2012),且初级生产力平均值与日本近海和胶州湾调查估算结果相当(Guo et al., 1992;孙松等,2005).空间分布上,钦州湾初级生产力与Chl-a浓度分布相似,茅尾海和三娘湾海区是春、夏、冬季初级生产力高值区,秋季出现低值区.茅尾海和三娘湾海区常年受茅岭江、钦江和大风江等河流输入的影响,大量营养物质的输送调控着该海湾浮游植物的初级生产过程(宋星宇等,2011).春、夏、冬季茅尾海和三娘湾海区也是浮游植物细胞数量的高值区(王迪等,2013),说明营养盐丰富的海区有利于基础营养阶层浮游生物的生长(Harding,1994; Glé et al., 2008).因此,春、夏、冬季茅尾海和三娘湾海区的初级生产力高值区主要受到上行效应的控制.秋季正值滤食性近江牡蛎等贝类生长旺盛期,加上浮游动物等上层捕食者的摄食压力,对浮游植物生长产生由上至下的影响(Huang et al., 2008),浮游植物生物量显著降低,秋季初级生产力内湾和三娘湾海区低值区主要受下行效应的控制.
钦州湾初级生产力在周年季节调查中表现出季节变化趋势:夏季>冬季>秋季>春季,并形成2个明显的高峰(1月和7月,图 5),与Chl-a浓度只在7月形成一个高峰(图 3)不一致,这表明初级生产力高的月份,其Chl-a浓度和浮游植物生物量并不一定高.水体中Chl-a浓度反映的仅是浮游植物现存量,对其初级生产力水平却不能完全体现(Gaxiola-Castro et al., 1995;叶宁等,2013).初级生产力水平相对较高的季节通常水体中浮游植物生物量也会相应升高,但钦州湾水产养殖业发达,常年养殖的近江牡蛎、对虾、青蟹和石斑鱼等海产及大量存在的浮游动物等对浮游植物的摄食压力很大,也会造成相反情况的出现.这种浮游植物现存量和生产量之间的差异从某种程度上也反映了海湾生态系统中能量的转换效率(叶宁等,2013).
4.3 调查海域环境要素对浮游植物生长的影响海水中Chl-a和初级生产力的时空变化反映了温度、盐度和营养盐等海洋环境要素对浮游植物生长的影响(Bronk et al., 1994; Lohrenzl et al., 1997; Glé et al., 2008).整个调查海区,夏季水温与Chl-a呈显著负相关,这与高爽等(2009)夏季在北黄海的调查结果一致.通常情况下,光照充足时,经过一定光照时间会导致水温上升,加快光合作用(Ryther,1969; Postma et al., 1970).但在夏季,强烈的光辐射会使表层海水升温的同时,也会对浮游植物产生光抑制作用,导致水体中Chl-a浓度有所下降(Falkowski,1992).夏、秋季盐度和Chl-a呈显著负相关,说明夏、秋季低盐水域有利于浮游植物的生长(文斐等,2012).
除光照、温度和盐度的影响之外,营养盐含量及其比值也是浮游植物生长的重要影响因子(Hecky et al., 1988;Agawin et al., 2000).夏、秋季NH+4-N与Chl-a呈显著正相关,这说明作为优先被浮游植物吸收的NH+4-N大大促进了浮游植物的生长繁殖.Bronk等(1994)通过藻类营养吸收动力学实验也证实了这个观点.调查期间,浮游植物的大量生长造成了P的大量消耗,全年PO3-4-P含量始终处于较低水平,而DIN和SiO2-3-Si含量相对于PO3-4-P较高(林美芳等,2014),因此,钦州湾浮游植物的生长可能受到P限制.海水中N/P比若过高会导致浮游植物受某一相对较低含量元素的限制(Redfield et al., 1963),此次冬季N/P比(35 ∶ 1)高于海水中浮游植物生长N/P的Redfield比值(16 ∶ 1),表明冬季调查海域营养盐限制因子是P.冬季整个调查海区Chl-a浓度与N/P比值呈显著的正相关,也充分说明了钦州湾冬季PO3-4-P可能是限制浮游植物生长的一个重要因子.
5 结论(Conclusions)1)春、夏、冬季钦州湾海域Chl-a浓度和初级生产力均在茅尾海和三娘湾海区出现高值区,秋季出现相反的分布趋势.Chl-a浓度季节变化特征为夏季>春季>冬季>秋季,初级生产力季节变化特征为夏季>冬季>秋季>春季.
2)钦州湾海域Chl-a和初级生产力的时空变化受陆源输入、水文环境、营养盐水平、浮游植物优势藻种以及滤食性贝类养殖活动等综合因素的影响,其中,陆源输入及贝类养殖活动是影响Chl-a和初级生产力时空变化的重要因素.
3)调查期间,水温、盐度和营养盐等海洋环境要素影响着浮游植物的生长,冬季PO3-4-P可能是限制浮游植物生长的一个重要因子.
致谢: 在样品的分析过程中,得到了防城港市海洋环境监测预报中心工作人员的大力协助,谨致谢忱!
| [1] | Agawin N S R,Duarte C M,Agustí S.2000.Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production[J].Limnology and Oceanography,45(3): 591-600 |
| [2] | Bronk D A,Glibert P M,Ward B B.1994.Nitrogen uptake,dissolved organic nitrogen release,and new production[J].Science,265(5180): 1843-1846 |
| [3] | Cadée G C.1975.Primary production off the Guyana coast[J].Netherlands Journal of Sea Research,9(1): 128-143 |
| [4] | 蔡玉婷.2010.福建近海叶绿素a和初级生产力的分布特征[J].农业环境科学学报,29(增刊): 174-179 |
| [5] | Falkowski P G.1992.Molecular ecology of phytoplankton photosynthesis// Falkowski P G,Woodhead A D.Primary productivity and biogeochemical cycles in the sea[M].New York: Plenum Press.47-67 |
| [6] | 高爽,李正炎.2009.北黄海夏、冬季叶绿素和初级生产力的空间分布和季节变化特征[J]. 中国海洋大学学报(自然科学版),39(4):604-610 |
| [7] | Gaxiola-Castro G,Garcia-Cordova J,Valdez-Holguin J E,et al.1995.Spatial distribution of chlorophyll a and primary productivity in relation to winter physical structure in the Gulf of California[J].Continental Shelf Research,15(9): 1043-1053; 1055-1059 |
| [8] | Glé C,Del Amo Y,Sautour B,et al.2008.Variability of nutrients and phytoplankton primary production in a shallow macrotidal coastal ecosystem (Arcachon Bay,France)[J].Estuarine,Coastal and Shelf Science,76(3): 642-656 |
| [9] | Gong G C,Shiah F K,Liu K K,et al.2000.Spatial and temporal variation of chlorophyll a,primary productivity and chemical hydrography in the southern East China Sea[J].Continental Shelf Research,20(4/5): 411-436 |
| [10] | Guo Y J,Yang Z Y.1992.Primary Productivity//Liu R Y.The Ecology and Biological Resource in the Jiaozhou Bay[M].Beijing: Academic Press of China.110-126 |
| [11] | 郝林华,孙丕喜,郝建民,等.2012.桑沟湾海域叶绿素a的时空分布特征及其影响因素研究[J].生态环境学报,21(2): 338-345 |
| [12] | Harding L W.1994.Long-term trends in the distribution of phytoplankton in Chesapeake Bay: roles of light,nutrients and streamflow[J].Marine Ecology Progress Series,104: 267-267 |
| [13] | Hecky R E,Kilham P.1988.Nutrient limitation of phytoplankton in freshwater and marine environments: a review of recent evidence on the effects of enrichment[J].Limnology and Oceanography,33(4): 796-822 |
| [14] | Huang C H,Lin H J,Huang T C,et al.2008.Responses of phytoplankton and periphyton to system-scale removal of oyster-culture racks from a eutrophic tropical lagoon[J].Marine Ecology Progress Series,358: 1-12 |
| [15] | Lalli C M,Parsons T R.1993.Biological Oceanography: An Introduction[M].New York: Pergamon Press |
| [16] | 蓝文陆,王晓辉,黎明民.2011.应用光合色素研究广西钦州湾丰水期浮游植物群落结构[J].生态学报,31(13): 3601-3608 |
| [17] | 李超伦,张芳,申欣,等.2004.胶州湾叶绿素的浓度、分布特征及其周年变化[J].海洋与湖沼,36(6): 499-506 |
| [18] | 李斌,谷伟丽,勒洋,等.2012.烟台四十里湾叶绿素a和初级生产力的分布特征[J]. 渔业科学进展,33(2): 19-23 |
| [19] | 李武,梁懋钢,陈涛.1993.海中准直光衰减系数与透明度盘深度的经验关系[J].海洋学报,15(5): 130-135 |
| [20] | 林美芳,钟秋平.2014.广西钦州湾营养状况季节分析与评价研究[J].环境科学与管理,39(6): 159-163 |
| [21] | Lohrenzl S E,Fahnenstiel G L,Redalje D G,et al.1997.Variations in primary production of northern Gulf of Mexico continental shelf waters linked to nutrient inputs from the Mississippi River[J].Marine Ecology Progress Series,155: 45-54 |
| [22] | 吕瑞华.1993.山东沿海浮游植物的同化系数[J].青岛海洋大学学报,23(3): 49-54 |
| [23] | Menzel D W,Ryther J H.1959—1960.The annual cycle of primary production in the Sargasso Sea off Bermuda[J].Deep Sea Research (1953),6: 351-367 |
| [24] | Oviatt C,Keller A,Reed L.2002.Annual primary production in Narragansett Bay with no bay-wide winter-spring phytoplankton bloom[J].Estuarine,Coastal and Shelf Science,54(6): 1013-1026 |
| [25] | Parsons T R,Maita Y,Lalli C M.1984.A manual for chemical and biological methods for seawater analysis[M].Oxford: Pergamon Press |
| [26] | Postma H,Rommets J W.1970.Primary production in the Wadden Sea[J].Netherlands Journal of Sea Research,4(4): 470-493 |
| [27] | Raymont J E.1980.Plankton and productivity// Ryther J H,Yentsch C S.The Oceans (2nd ed) [M].Oxford: Pergamon Press |
| [28] | Redfield A C,Ketchum B H,Richards F A.1963.The influence of organisms on the composition of sea water// Hill M N.The Sea,volume 2.New York: Interscience Press.26-77 |
| [29] | Reynolds C S.1994.The long,the short and the stalled: On the attributes of phytoplankton selected by physical mixing in lakes and rivers[J].Hydrobiologia,289(1/3): 9-21 |
| [30] | Ryther J H.1969.Photosynthesis and fish production in the sea[J].Science,166(3901): 72-76 |
| [31] | Sarmiento J L,Toggweiler J R,Najjar R,et al.1988.Ocean carbon-cycle dynamics and atmospheric pCO2[J].Philosophical Transactions of the Royal Society of London Series A,325: 3-21 |
| [32] | Sathyendranath S,Gouveia A D,Shetye S R,et al.1991.Biological control of surface temperature in the Arabian Sea[J].Nature,349(6304): 54-56 |
| [33] | Song X Y,Huang L M,Zhang J L,et al.2004.Variation of phytoplankton biomass and primary production in Daya Bay during spring and summer[J].Marine Pollution Bulletin,49(11/12): 1036-1044 |
| [34] | 宋星宇,谭烨辉,黄良民,等.2011.三亚湾春夏季初级生产力的分布特征及环境影响研究[J].海洋环境科学,30(1): 19-23 |
| [35] | 孙军,刘东艳,柴心玉,等.2003.1998-1999年春秋季渤海中部及其邻近海域叶绿素a浓度及初级生产力估算[J].生态学报,23(3): 517-526 |
| [36] | 孙松,张永山,吴玉霖,等.2005.胶州湾初级生产力周年变化[J].海洋与湖沼,36(6): 481-486 |
| [37] | 孙晓霞,孙松,张永山,等.2011.胶州湾叶绿素a及初级生产力的长期变化[J].海洋与湖沼,42(5): 654-661 |
| [38] | Valdéz-Holguín J E,Lara-Lara J R.1987.Primary productivity in the Gulf of California effects of El Niño 1982-1983 event[J].Ciencias Marinas,13(2): 34-50 |
| [39] | 王迪,陈丕茂,逯晶晶,等.2013.钦州湾浮游植物周年生态特征[J].应用生态学报,24(6): 1686-1692 |
| [40] | 韦蔓新,童万平,赖廷和,等.2001.钦州湾内湾贝类养殖海区水环境特征及营养状况初探[J].黄渤海海洋,19(4): 51-55 |
| [41] | 韦蔓新,赖廷和,何本茂.2002.钦州湾近20a来水环境指标的变化趋势Ⅰ平水期营养盐状况[J].海洋环境科学,21(3): 49-52 |
| [42] | 韦蔓新,何本茂.2004.钦州湾近20a来水环境指标的变化趋势Ⅲ微量重金属的含量分布及其来源分析[J].海洋环境科学,23(1): 29-32 |
| [43] | 文斐,孙晓霞,郑珊,等.2012.2011年春、夏季黄、东海叶绿素a和初级生产力的时空变化特征[J].海洋与湖沼,43(3): 438-444 |
| [44] | 薛保铭,杨惟薇,王英辉,等.2013.钦州湾水体中磺胺类抗生素污染特征与生态风险[J].中国环境科学,33(9): 1664-1669 |
| [45] | 杨斌,方怀义,钟秋平,等.2012a.钦州湾夏季营养盐的分布特征及富营养化评价[J].海洋通报,31(6): 640-645 |
| [46] | 杨斌,钟秋平,李宗活,等.2012b.钦州湾表层海水重金属分布特征及其污染评价[J].广州化工,40(11): 146-147; 173 |
| [47] | 杨斌,钟秋平,鲁栋梁,等.2013a.钦州湾秋季营养盐分布特征及营养状态分析研究[J].环境科学与管理,38(12): 40-44 |
| [48] | 杨斌,钟秋平,张晨晓,等.2013b.钦州湾春季水质营养状况分析与评价[J].中国环境监测,29(5): 103-106 |
| [49] | 杨斌,鲁栋梁,钟秋平,等.2014a.钦州湾近岸海域水质状况及富营养化分析[J].中国环境监测,30(3): 60-64 |
| [50] | 杨斌,钟秋平,鲁栋梁,等.2014b.钦州湾海域COD时空分布及对富营养化贡献分析[J].海洋科学,38(3): 20-25 |
| [51] | 叶宁,王彦,申玉春,等.2013.广东流沙湾叶绿素a含量和初级生产力状况的研究[J].海洋与湖沼,44(5): 1306-1310 |
| [52] | 中国海湾志编纂委员会.1993.中国海湾志,第十二分册: 广西海湾[M].北京: 海洋出版社 |
| [53] | 周伟华,袁翔城,霍文毅,等.2004.长江口邻域叶绿素a和初级生产力的分布[J].海洋学报,26(3): 143-150 |
| [54] | Zuenko Y,Selina M,Stonik I.2006.On conditions of phytoplankton blooms in the coastal waters of the North-Western East/Japan Sea[J].Ocean Science Journal,41(1): 31-41 |