 2015, Vol. 35
2015, Vol. 35



2. 云南高原湖泊流域污染过程与管理重点实验室, 昆明 650034
2. Yunnan Key Laboratory of Pollution Process and Management of Plateau Lake-Watershed, Kunming 650034
自1970年代开始,湖泊富营养化成为全球性的环境问题并得到广泛关注(Schindler,1974; Carpenter et al., 1999;Smith,2003; Conley et al., 2009).之前的富营养化研究涉及多个方面,如:成因、影响、危害、控制及管理等,其中对成因的识别是科学确定控制与管理对策的基础.在全面评估美国北部和加拿大南部的一些湖泊的主要过程后,科学家们确定氮(N)、磷(P)营养物质的输入和累积是富营养化的主要原因(Pant et al., 1980;Paerl,2006; Carpenter and Bennett, 2011).此后,湖沼学的研究逐渐发现:造成并影响湖泊富营养化的因子主要是营养盐、水温、水力条件、气象因子(Beklioglu and Moss, 1995; Thomas and Eckmann, 2007;刘利霞,2008;万能等,2008;邱立云,2009; 杨霞等,2012);但湖泊富营养化也可能与地理、地质、气候、生态系统结构、湖泊形态及流域人为活动等其它一些因素密切相关(金相灿,2008; 吴锋等,2012).湖泊富营养化的影响因子涉及水文、水化学、水生物等方面,具有复合性和非线性特征;加之不同湖泊在自然特征、流域社会经济发展压力、水质污染特征等方面的差异,使得识别水质退化与富营养化的主要驱动因子充满挑战,亦使得富营养化防治及预防措施难以具有高度普适性(Anderson et al., 2002; Schindler et al., 2008;Finlay et al., 2013).
了解湖泊的地理位置、形态、水质、流域社会经济状况,有助于分析并预测不同影响因子与富营养化的定量关系,识别影响的关键因子,以较低的成本、较短的时间达到理想的控制效果(Fernandes et al., 2012; 刘永等,2012; Finlay et al., 2013).同时,识别多个湖泊的富营养化影响因子,比较其定量关系的共性及差异性特征,可以扩展富营养化治理措施的适用性.对于富营养化发生机理尚不明晰且缺乏长序列观测数据的湖泊,也可尝试借鉴其他自然条件与响应关系类似的湖泊的控制措施,从而节约监测支出.云南高原湖泊群是我国5大湖泊分区之一,50多个湖(库)面积大于1 km2,约占全国湖泊总淡水储量的13.4%.这些湖泊普遍具备一些基本的自然和压力特征,如:半封闭、入湖河流源近流短;流域面积与湖泊体积比值较小,水体交换周期长;入湖水量少,流域水资源和水环境承载力小、压力大;湖盆区为主要的流域人口和经济聚集区;高敏感性,对快速变化环境的响应特征与其他区域不同,具有易发生富营养化的气候特征,部分湖泊富营养化严重(蓝红林,2002;曹金玲等,2012).湖区工农业生产的发展及人口增长,资源需求量迅速增加,入湖的污染负荷也迅速增加,造成了日益严重的湖泊水污染.其中滇池、异龙湖等富营养化现象尤为突出,水质急剧下降.本文以4个云南高原湖泊(滇池、程海、抚仙湖、异龙湖)为研究对象,比较其自然条件、湖泊理化特征、流域社会经济条件的异同;在此基础上,构建包括绝对主成分多元线性回归分析模型(absolute principle components score and multivariate Linear Regression,APCS-MLR)、结构方程模型(structural equation modeling,SEM)及人工神经网络模型(artificial neural network,ANN)在内的综合分析方法,重点关注并确定浮游初级生产力的代表指标(叶绿素a,Chla)与影响因子间的定量相关关系,识别影响富营养化水平的关键因素,比较关键因子的差异性.
2 研究对象(Study cases)滇池、程海、抚仙湖和异龙湖同属云南高原湖泊,选择其作为分析对象的原因在于:①4个湖泊均面临一定程度的富营养化压力,但营养状态不同;②其年均气温、气候条件等有一定的差异(表 1),湖泊形态和水文特征差异较大(表 2);③流域社会经济发展状况不同,对湖泊的压力差异明显.滇池流域人口最多(表 3),以城镇人口为主,第一产业所占比重最低;程海的人口数量仅次于滇池流域,但农业人口比例最高,支柱产业为农业及农特产品加工业;抚仙湖与异龙湖流域的人口数量较少,以农业人口为主.对上述的4个湖泊进行富营养化的影响因子对比分析,有助于识别关键的驱动因子;同时,还可识别湖泊形态、水文特征、社会经济活动对于湖泊水质恶化、富营养化影响的差异性作用,并可为云南高原其他监测序列不足的湖泊的富营养化控制对策提供借鉴.在前期的研究中(颜小品等,2013),以滇池外海为例(1998—2008年,247个样本)开展了富营养化影响因子的识别.在此基础上,本文将对其他3个湖泊进行分析,并对4个湖泊的结果与影响因素进行对比.研究采用1998—2009年间的实测水质数据,其中:异龙湖1998—2009年共144个样本,抚仙湖2005—2009年共188个样本,程海2005—2009年共204个样本.
| 表1 4个湖泊自然环境特征对比 Table 1 Comparison of the natural characteristics of four lakes |
| 表2 4个湖泊形态和水文特征对比 Table 2 Comparison of the shape and hydrographical characteristics of four lakes |
| 表3 4个湖泊人口与社会经济状况对比 Table 3 Comparison of the population and economy of four lakes |
本研究的主要方法与步骤包括(图 1):①在分析湖泊的自然环境、湖泊形态及流域状况后,建立APCS-MLR模型,降低指标维度,筛选出对Chla影响较大的指标;②利用SEM判别核心驱动因子与Chla间的定量关系,通过模型评价筛选不符合条件的指标;③利用ANN对SEM的结果作辅助检验和判定.对指标数较少的湖泊不作降维处理,直接进行SEM建模.上述过程分别在LISEREL87和MATLAB 7.11.0(R2010b)中实现.与多元回归模型相比,ANN方法的优点在于无需预先设定输入-输出参数间的响应关系(邹锐等,2012),在本文中被用来对比验证SEM模型结论的可靠性;但由于其应用较多,本文在此不对其做进一步的阐述,仅对APCS-MLR和SEM做简要分析.
 |
| 图 1 技术路线图 Fig. 1 Methodological framework |
利用APCS-MLR筛选出影响Chla的共性因子,确定其对Chla的贡献权重和影响(Thurston and Spengler, 1985;Singh et al., 2005),该方法的核心是确定各主成分的绝对得分(APCSjk)(Singh et al., 2005;Zhou et al., 2007).在确定APCSjk后,对因变量(观测点的Chla浓度)与自变量(各主成分绝对得分APCSjk)做多元回归分析(Singh et al., 2008).

SEM是实现对潜变量分析的重要工具,是验证性因子模型和因果模型的结合(Sobel,1982; Bollen,1998;Barbara,2013).因子模型部分称为测量模型,其中的方程称为测量方程,描述潜变量与指标之间的关系.因果模型部分称为潜变量模型(或结构模型),其中的方程称为结构方程,描述潜变量之间的关系.SEM有一系列重要的假设:①测量方程误差项ε、δ和结构方程残差项ζ的均值为0;②误差项ε、δ与潜变量η、ξ之间不相关,ε与δ不相关;③结构方程残差项ζ与ξ、ε、δ间不相关.在SEM中,需要估计的参数矩阵共有8个:Λx、Λy、B、Γ、Φ、Ψ、Θε、Θδ. 全部指标组成的协方差矩阵可表示为8个参数矩阵的函数(Bollen,1989):
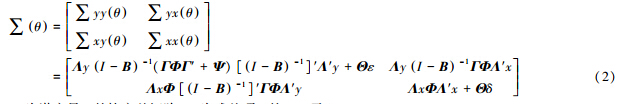

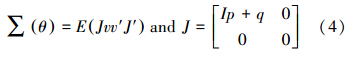
如果理论模型为真,则∑ θ等于总体的协方差矩阵∑,即∑ = ∑ θ ,从而观测变量的方差和协方差都是模型参数的函数.要评价模型的拟合效果,需要比较再生协方差和样本协方差矩阵的差异.这2个矩阵的整体差异,可用拟合指数表示:
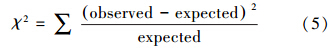
| 表4 拟合指数与评价标准 Table 4 Modeling index st and ards |
以抚仙湖为例加以分析.在数据经标准化处理后,对189个样本进行APCS分析,用最大方差法旋转得到主成分负荷和主成分得分系数表(表 5);将每个绝对主成分得分对Chla做多元回归:

可选出PC2~PC5为对叶绿素有重要影响的4个主成分,对应的高载荷监测指标分别为T、DOS、CODMn、EC;P、DO、LAS;SD、pH.同理,在异龙湖的APCS-MLR分析中,载荷较大的监测指标为悬浮物(SS)、总硬度(THW)、DO、CODMn、BOD5、NH3-N、亚硝氮(NO2-N)、硝态氮(NO3-N)、VPh、As、Cr6+、Cd、Cu、TP、TN、F、色度(Chro)、EC、细菌总数(TPC)和大肠杆菌(Coli).滇池外海已有的研究则发现,在5个主成分中,载荷较大的监测指标有14个,分别是 T、pH、DO、BOD5、SD、CODMn、CODCr、TN、锌(Zn)、EC、OT、Cu、Cr6+、NH3-N(颜小品等,2013).由于程海的水质监测指标数量较少,因此不作降维处理,直接进行SEM分析.
| 表5 抚仙湖旋转后各指标的主成分负荷表 Table 5 Principle components loadings of the measured water quality index on VARIMAX rotated factors in Lake Fuxian |
分别依据APCS-MLR筛选出的主要驱动因子,对4个湖泊单独构建SEM模型,并进行模型结构修正,其中滇池的结果见颜小品等(2013),抚仙湖、程海、异龙湖的结果见图 2和表 6.
 |
| 图 2 抚仙湖、程海、异龙湖的SEM模拟结果 Fig. 2 SEM results of Lake Fuxian, Lake Chenghai, and Lake Yilong |
| 表6 4个湖泊数据特征和SEM拟合指数 Table 6 Data characteristics and SEM model index of four lakes |
对4个湖泊的SEM模型的路径系数对比发现(表 7):在4个湖泊中,与Chla浓度变化相关关系最大的均为理化因子,但各个湖泊理化因子(T、pH、SD、DO、SS等)对Chla浓度影响的正、负方向不同.温度、溶解氧与Chla的相关关系较大,如:滇池的藻类以蓝藻植物门中的微囊藻属、束丝藻属、颤藻属及绿藻植物门中的衣藻属、转板藻属为优势属.实验模拟得出在20-35 ℃之间,这些藻的生长曲线随温度的变化而差异显著(刘玉生等,1995);因此温度对湖泊水动力学过程和生态系统产生直接影响,从而促进了藻类水华的暴发.溶解氧与湖泊生态系统间则存在复杂的反馈关系,溶解氧的降低会危害水生动物、加速内源释放,加剧湖体的富营养化;而藻类增多或光合作用的增强亦可能导致溶解氧浓度的增加.
| 表7 4个湖泊SEM路径系数对比 Table 7 Comparison of SEM path coefficients of four lakes |
滇池、程海的SEM结果显示透明度与Chla呈负相关.从机理推断,透明度的降低会导致湖泊光合作用水平的下降,降低初级生物量;但与此同时,浮游植物的增加将降低水体透明度,此时浮游植物和透明度为反向关系,SEM的模型结果正说明了二者间的反向关系.
4.3 基于ANN结果的辅助验证以影响因子作为输入,Chla作为输出,对4个湖泊进行ANN分析,以辅助对比并验证SEM,模拟结果显示ANN分析得出的隐层权重与SEM路径系数基本一致.以程海为例,将SEM的所有指标,即NH3-N、TP、T、pH、DO和SD作为神经网络的输入因子,输入因子数量为6,输出因子为Chla,神经网络设3层隐层.6个因子对第一隐层的权重如表 8所示.
| 表8 程海神经网络驱动因子权重表 Table 8 Weight of driving factors in neural networks in Lake Chenghai |
就输入节点初始权重而言,pH值所占权重最大,T所占权重最小,此结果与SEM路径系数一致;而营养盐中NH3-N所占系数高于TP,也与SEM路径系数一致(表 8),说明SEM分析结果较为可信.对于滇池而言,ANN结果表明,理化因子组4个指标在各层隐层神经元中所占权重大小排序是T>DO>SD>pH,与SEM的结果一致;异龙湖的ANN分析显示N的权重值高于P,从而对SEM的结果进行了合理的验证.
4.4 基于湖泊特征分析的Chla影响因子对比分析由表 1~表 3可知,抚仙湖为贫营养,Chla浓度低,未发生全湖性的藻类暴发的现象,营养物质的作用不明显;其余3个湖泊中N、P营养盐对Chla均有显著影响.结合4个湖泊多年平均N、P浓度和相应分析(表 9),滇池的P不是藻类生长的限制性因素,而N则因为含量偏高而与藻类生长有非常微弱的负向关系.异龙湖N含量最高,氮磷比也最高,但和贫营养的抚仙湖相比,异龙湖的P浓度仍高一个数量级.程海为中营养湖泊,但也会发生偶发性的蓝藻暴发.程海的N浓度与抚仙湖在一个数量级,N的增加会促进藻类生长,P含量则远高于抚仙湖.分别总结各湖泊理化因子对Chla的影响并建立定量表达关系,可在一定程度上为蓝藻暴发的预警提供支持.
| 表9 4个湖泊氮磷浓度对比 Table 9 Concentrations of nitrogen and phosphorus of four lakes |
Harpole 等(2011)通过对641个湖泊的研究表明,65%的湖泊更易发生N限制(包括单纯N限制和N、P共限制),28%的湖泊更容易发生N、P共限制现象.而本研究的结论为滇池为单纯N限制湖泊,程海和异龙湖为N、P共限制湖泊,与其结论一致.另有研究表明(沈颂东,2003;顾启华,2006;向律成等,2009),氮磷比例低于15时,营养物质增加会造成藻类物质的增加,加重富营养化水平;而当氮磷比例超过15时,营养物质增加则会对藻类生长起到一定程度的抑制作用,但抑制作用非常有限.
本研究的4个湖泊均属内陆高原湖泊,随着流域内污染负荷的增加,湖体水质都存在不同程度的恶化并呈加剧态势.4个湖泊的流域面积、湖泊形态各异(表 2),流域污染源排放、污染负荷及湖体的水动力条件不同,影响了营养物质在湖体中的迁移转换,造成了富营养化响应的差异性.流域湖泊面积比越大,说明单位水面面积负载的流域面积越大、污染负荷也越重,更易超出水体的纳污能力而发生富营养化现象.流域面积/湖泊面积比值最大的异龙湖和滇池都是富营养化湖泊,而比值较小的程海和抚仙湖则分别为中营养和贫营养湖.
程海和抚仙湖为深水湖,滇池和异龙湖为浅水湖泊.研究表明,水深是湖泊大型水生植物能否大面积分布的主要影响因素(倪兆奎等,2011).在浅水湖泊,随着营养盐含量的增加,水生植物群落结构简单化,高等植物种类和数量将出现下降趋势,浮游植物种类也出现下降趋势,但生物量会迅速增加.深水湖泊中,营养盐的输入对水质变化很重要,但水位变化同样不可忽视.随着营养盐的增加和水位的波动,耐污群落结构的增长趋势将十分明显,直到营养盐达到水生植物的阈值(刘永等,2006).因此相对而言,浅水湖泊更容易发生富营养化.
湖泊所在流域的社会经济状况会影响到流域的负荷产生量与入湖量.数据表明(表 3),人口规模越大,污染负荷产生量越多,但污染负荷多的湖泊不一定发生富营养化现象.4个湖泊的比较结果也显示,城市人口比例越高,城市生活源的比例越高,越容易发生富营养化现象.滇池流域人口最多、城市人口比例最高,以第二、三产业为主,污染源以城市生活源为主,富营养化情况最为严峻.其他3个湖泊城市人口的比例较低,污染源以农业面源污染为主;城市人口比例较低的抚仙湖和程海并未发生富营养化.
2001—2008年的监测数据显示,滇池水体的多年平均氮磷比为15.88,程海、抚仙湖和异龙湖水中的氮磷比分别为25.26、21.29和56.87;滇池、抚仙湖与异龙湖3个湖泊污染源的氮磷比与水质氮磷比的大小关系一致.单从污染源的氮磷比来看,由于农业面源中P含量远高于城镇生活源的P含量,因此城镇生活源的氮磷比高于农业面源污染.但由于滇池流域面积远远大于其他湖泊,农业用地面积也非常大,且流域南部为富磷区;因此即便污染源中城镇生活源的比例较高,其P污染也非常多,相应的氮磷比也较低,N限制作用明显.而程海和异龙湖由于P较少,呈现出N、P共限制的营养盐-藻类动力结构.
5 结论(Conclusions)1)本研究选取滇池、程海、抚仙湖和异龙湖4个高原湖泊,建立了表征湖泊富营养水平的Chla与相关影响因素间的定量关系模型,得知:4个湖泊中,对Chla浓度变化影响最大的为理化因子,但在各湖中该影响的正、负性及不同理化因子的贡献有较大差异.
2)基于湖泊特征分析可知:流域污染来源的不同,一定程度影响入湖负荷的差异,从而使湖泊表现出不同的N、P限制性特征;水位对湖泊生物群落结构有着重要影响,浅水湖泊生态系统较为脆弱,湖水与底泥间物质交换强烈、沉积较为缓慢,具有较低的污染负荷承载能力;4个湖泊的流域面积、湖泊形态差异,流域污染源排放及湖体水动力条件均影响营养盐在湖体中的迁移转换,从而造成4个湖泊富营养化的差异性特征;4个湖泊中城市人口比例较高的滇池和异龙湖,其富营养化最严重.
3)本研究采用APCS-MLR、SEM和ANN分析方法探究不同湖泊Chla与影响因子的定量关系,结合机理过程对比验证模型结果,一定程度上克服了统计模型所固有的缺点.然而,要揭示Chla与其它因子间的内在因果关联,则必须依靠更为深入的机理过程加以确认.
| [1] | Anderson D M, Glibert P M, Burkholder J M. 2002. Harmful algal blooms and eutrophication: nutrient sources, composition, and consequences[J]. Estuaries, 25(4): 704-726 |
| [2] | Beklioglu M, Moss B. 1995. The impact of pH on interactions among phytoplankton algae, zooplankton and perch (Perca fluviatilis) in a shallow, fertile lake[J]. Freshwater Biology, 33(3): 497-509 |
| [3] | Bollen K A. 1989. Structural Equations with Latent Variables[M]. New York: Wiley and Sons, Inc. |
| [4] | Bollen K A. 1998. Structural equation models[M]. New York: John Wiley & Sons, Inc. |
| [5] | Barbara M B. 2013.Structural Equation Modeling With AMOS: Basic Concepts, Applications, and programming.[M] USA: Routledge.23-42 |
| [6] | Burnham K P, Anderson D R. 2002. Model selection and multimodel inference: a practical information-theoretic approach[M]. New York: Springer |
| [7] | 曹金玲, 席北斗, 许其功, 等. 2012. 地理气候及湖盆形态对我国湖泊营养状态的影响[J]. 环境科学学报, 32(6): 1512-1519 |
| [8] | Carpenter S R, Bennett E M. 2011. Reconsideration of the planetary boundary for phosphorus[J]. Environmental Research Letters, 6(1): 14009 |
| [9] | Carpenter S R, Ludwig D, Brock W A. 1999. Management of eutrophication for lakes subject to potentially irreversible change[J]. Ecological Applications, 9(3): 751-771 |
| [10] | Conley D J, Paerl H W, Howarth R W, et al. 2009. Controlling eutrophication: nitrogen and phosphorus[J]. Science, 323(5917): 1014-1015 |
| [11] | Fernandes J A, Kauppila P, Uusitalo L, et al. 2012. Evaluation of reaching the targets of the Water Framework Directive in the Gulf of Finland[J]. Environmental Science & Technology, 46(15): 8220-8228 |
| [12] | Finlay J C, Small G E, Sterner R W. 2013. Human Influences on Nitrogen Removal in Lakes[J]. Science, 342(6155): 247-250 |
| [13] | 顾启华. 2006. 富营养化水体中藻类水华成因分析与研究[D]. 天津: 天津大学, 41-44 |
| [14] | Harpole W S, Ngai J T, Cleland E E, et al. 2011. Nutrient co-limitation of primary producer communities[J]. Ecology Letters, 14(9): 852-862 |
| [15] | 金相灿. 2008. 湖泊富营养化研究中的主要科学问题——代"湖泊富营养化研究"专栏序言[J]. 环境科学学报, 28(1): 21-23 |
| [16] | 蓝红林. 2002. 云南高原湖泊的治理与保护[J]. 水电站设计, 18(4): 58-60 |
| [17] | 刘利霞. 2008. 滇池水体富营养化成因及控制措施探讨[J]. 菏泽学院学报, 30(2): 86-89 |
| [18] | 刘永, 郭怀成, 周丰, 等. 2006. 湖泊水位变动对水生植被的影响机理及其调控方法[J]. 生态学报, 26(9): 3117-3126 |
| [19] | 刘永, 阳平坚, 盛虎, 等. 2012. 滇池流域水污染防治规划与富营养化控制战略研究[J]. 环境科学学报, 32(8): 1962-1972 |
| [20] | 刘玉生, 韩梅, 梁占彬, 等. 1995. 光照、温度和营养盐对滇池微囊藻生长的影响[J]. 环境科学研究, 8(6): 7-11 |
| [21] | Mcardle J J, Mcdonald R P. 1984. Some algebraic properties of the reticular action model for moment structures[J]. British Journal of Mathematical and Statistical Psychology, 37(2): 234-251 |
| [22] | Mcdonald R P, Ho M H R. 2002. Principles and practice in reporting structural equation analyses[J]. Psychological Methods, 7(1): 64-82 |
| [23] | 倪兆奎, 王圣瑞, 金相灿, 等. 2011. 云贵高原典型湖泊富营养化演变过程及特征研究[J]. 环境科学学报, 31(12): 2681-2689 |
| [24] | Paerl H W. 2006. Assessing and managing nutrient-enhanced eutrophication in estuarine and coastal waters: Interactive effects of human and climatic perturbations[J]. Ecological Engineering, 26(1): 40-54 |
| [25] | Pant M C, Sharma A P, Sharma P C. 1980. Evidence for the increased eutrophication of Lake Nainital as a result of human interference[J]. Environmental Pollution Series B, Chemical and Physical, 1(2): 149-161 |
| [26] | 邱立云. 2009. 富营养化湖泊环境影响因子相关性分析研究[D]. 武汉: 华中科技大学, 25 |
| [27] | Schindler D W. 1974. Eutrophication and recovery in experimental lakes: implications for lake management[J]. Science, 184(4139): 897-899 |
| [28] | Schindler D W, Hecky R E, Findlay D L, et al. 2008. Eutrophication of lakes cannot be controlled by reducing nitrogen input: results of a 37-year whole-ecosystem experiment[J]. Proceedings of the National Academy of Sciences of the United States of America, 105(32): 11254-11258 |
| [29] | 沈颂东. 2003. 氮磷比对小球藻吸收作用的影响[J]. 淡水渔业, 33(1): 23-25 |
| [30] | Singh K P, Malik A, Kumar R, et al. 2008. Receptor modeling for source apportionment of polycyclic aromatic hydrocarbons in urban atmosphere[J]. Environmental Monitoring and Assessment, 136(1/3): 183-196 |
| [31] | Singh K P, Malik A, Sinha S. 2005. Water quality assessment and apportionment of pollution sources of Gomti river (India) using multivariate statistical techniques-a case study[J]. Analytica Chimica Acta, 538(1/2): 355-374 |
| [32] | Smith V H. 2003. Eutrophication of freshwater and coastal marine ecosystems a global problem[J]. Environmental Science and Pollution Research, 10(2): 126-139 |
| [33] | Sobel M E. 1982. Asymptotic confidence intervals for indirect effects in structural equation models[J]. Sociological Methodology, 13: 290-312 |
| [34] | Steiger J H. 1990. Structural model evaluation and modification: An interval estimation approach[J]. Multivariate Behavioral Research, 25(2): 173-180 |
| [35] | Thomas G, Eckmann R. 2007. The influence of eutrophication and population biomass on common whitefish (Coregonus lavaretus) growth-the Lake Constance example revisited[J]. Canadian Journal of Fisheries and Aquatic Sciences, 64(3): 402-410 |
| [36] | Thurston G D, Spengler J D. 1985. A quantitative assessment of source contributions to inhalable particulate matter pollution in metropolitan Boston[J]. Atmospheric Environment (1967), 19(1): 9-25 |
| [37] | 万能, 宋立荣, 王若南, 等. 2008. 滇池藻类生物量时空分布及其影响因子[J]. 水生生物学报, 32(2): 184-188 |
| [38] | 吴锋, 战金艳, 邓祥征, 等. 2012. 中国湖泊富营养化影响因素研究——基于中国22个湖泊实证分析[J]. 生态环境学报, 21(1): 94-100 |
| [39] | 向律成, 郝虎林, 杨肖娥, 等. 2009. 多年生漂浮植物对富营养化水体的响应及净化效果研究[J]. 水土保持学报, 23(5): 152-155 |
| [40] | 颜小品, 李玉照, 刘永, 等. 2013. 基于结构方程模型的滇池叶绿素a与关键影响因子关系识别[J]. 北京大学学报(自然科学版), 49(6): 1031-1039 |
| [41] | 杨霞, 胡兴娥, 陈磊, 等. 2012. 藻类水华暴发影响因子研究综述[A]//2012中国环境科学学会学术年会论文集(第2卷)[C]. 北京: 中国环境科学学会. 1477-1478 |
| [42] | Zhou F, Huang G H, Guo H C, et al. 2007. Spatio-temporal patterns and source apportionment of coastal water pollution in eastern Hong Kong[J]. Water Research, 41(15): 3429-3439 |
| [43] | 邹锐, 董云仙, 张祯祯, 等. 2012. 程海富营养化机理的神经网络模拟及响应情景分析[J]. 生态学报, 32(2): 448-456 |