 2015, Vol. 35
2015, Vol. 35



利用微藻进行污水处理的历史追溯已久. 早在20世纪50年代,Oswald等就提出利用微藻处理污水的设想(Oswald et al., 1957). 此后,以藻-菌共生体系和高效藻类塘为代表的悬浮生长藻类塘系统在分散式污水处理中得到了广泛的工程应用(Muñoz and Guieysse, 2006; Park et al., 2011; 邢丽贞等,2009). 但这类系统因占地面积大、处理效果不稳定等局限性,一直未能成为污水处理的主流工艺. 近年来,在市政污水处理厂深度净化需要以及渴望从污水中获得生物柴油的驱动下,微藻污水处理在世界范围内重获新生(Christenson and Sims, 2011; Olguín,2012; Pittman et al., 2011; 胡洪营等,2009).
微藻生长过程需要大量吸收氮(N)、磷(P)等营养元素,可直接降低二/三级出水中N、P等污染物的含量. 通过固定二氧化碳(CO2)、产生氧气(O2)、提高pH等间接作用,微藻还能创造出有效去除水中残留有机物和病原性微生物的环境条件. 此外,微藻也具有吸附重金属等有害物质的能力(胡洪营等,2009). 因此,微藻具有成为污水深度净化技术的良好潜力. 在污水二/三级处理中,去除营养元素的常见藻种包括: ①绿藻门的小球藻(Chlorella)、葡萄藻(Botryococcus)、栅藻(Scenedesmus)和微绿球藻(Nannochloris)等,其中尤以小球藻(Chlorella)和栅藻(Scenedesmus)的研究报道为多(Cai et al., 2013; 胡洪营等,2009);②蓝藻门的节旋藻属(Arthrospira sp.)、颤藻属(Oscillatoria sp.)和席藻属(Phormidium);③硅藻门的三角褐指藻(P. tricornutum)等(Cai et al., 2013).以上种属的N、P去除效果可参见Cai等的综述文章(Cai et al., 2013). 在藻种选择的基础上,微藻培养系统(反应器)的构建是实现微藻污水处理工程化应用的关键. 按微藻的生长方式不同,微藻培养系统可分为悬浮培养和附着培养两大类. 悬浮培养系统可进一步分为开放式和封闭式两类:①开放式系统主要指各类塘系统,典型的如高效藻类塘和跑道式藻类塘等;②封闭式系统主要指各类光生物反应器,分为管式(垂直、水平、螺旋)、圆柱式、薄板式和袋式等. 附着式系统包括光生物膜(平板)反应器和藻细胞固定化(Cai et al., 2013; Christenson and Sims, 2011;胡洪营等,2009). 考虑到污水处理的实际情况(水量大,建造、运行成本等),开放培养系统仍将是微藻污水处理的主流反应器构型.
如上所述,藻细胞用于生产生物柴油是微藻污水处理重获新生的主要驱动力之一(Olguín,2012; Rawat et al., 2011; 胡洪营等,2009; 蔡卓平等,2012). 通过微藻生产生物柴油具有其他任何产油作物无法比拟的优势(胡洪营等,2009):①藻细胞的光合效率高,生长速度快、周期短,其产油量为47000~190000 L · hm-2 · a-1,是农作物的7~30倍;②生物质燃油热值高,平均达33 MJ · kg-1,是木材或农作物秸秆的1.6倍;③不需占用农业用地;④生物质(藻细胞)生产和加工成本低,尤其是以污水为底物进行藻细胞培养时. 有鉴于此,美国、欧洲、澳大利亚、日本、台湾等发达国家和地区都已将微藻培养作为实现污水生态处理和可再生能源生产的战略发展目标(Christenson and Sims, 2011; Pittman et al., 2011; U.S.DOE,2010). 常见的产油藻种及其油脂含量文献已述及(胡洪营等,2009). 工业上以产油为目的的微藻培养一般采用封闭式光生物反应器,并且往往采用纯培养或单株培养的方式(Christenson and Sims, 2011). 当结合污水处理目标时,因巨大的水量及污水中复杂的成分(尤其是其中包含的混合种属),以上培养方式将很难维持.
近年来,国内外学者在开发微藻污水深度净化和可再生能源生产潜力方面进行了大量研究;在污水净化机理、藻种筛选、反应器设计、工艺条件控制及藻细胞加工利用等方面都取得了积极的进展(Abdel-Raouf et al., 2012; Christenson and Sims, 2011; Muñoz and Guieysse, 2006; 蔡卓平等,2012). 然而,无论从污水净化本身,还是能源生产来说,藻细胞的分离、采收都一直是一个悬而未决的基础性技术难题. 微藻细胞一般小于30 μm,带负电荷,密度接近于水,这些特性使得藻细胞在水中往往处于稳定的悬浮状态,很难像活性污泥那样通过重力沉淀而实现自然分离(Christenson and Sims, 2011). 结果,藻细胞会随处理水大量流失,不仅二次污染处理水,而且导致反应器内生物量难以大量维持(一般仅为0.2~0.6 g · L-1(Christenson and Sims, 2011; 胡洪营和李鑫,2010)). 低的培养密度导致去除效率低下,使得处理效果稳定性较差. 对此,往往需降低处理负荷,同时采用较长的水力停留时间(HRT),进而导致占地面积加大. 目前普遍应用的藻类塘系统HRT一般为2~6 d,当量人口占地一般>10 m2(Christenson and Sims, 2011; Muñoz and Guieysse, 2006). 显然,其占地面积要比二/三级污水处理主体单元还要庞大许多,这在用地紧张的城市中是很难被接受的.
从能源生产角度看,满足工业利用要求的藻细胞原料其最佳生物量应达到300~400 g · L-1(干质量). 因此,常规培养下的藻液需浓缩1 000倍以上后方能在工业上加以利用. 这一高能耗的分离、浓缩过程是微藻能源生产中的主要能耗成本(占微藻生物质生产总成本的20%~50%(Milledge and Heaven, 2013; 胡洪营和李鑫,2010). 过高的生产成本使得藻类生产生物柴油与化石燃料相比仍处于劣势.
可见,藻细胞分离、采收困难是限制微藻技术大规模工业化应用的重要瓶颈. 微藻分离、采收常用的方法包括离心法、过滤法(包括膜滤)、气浮法、直接重力沉降法和絮凝法等(Christenson and Sims, 2011; Milledge and Heaven, 2013; 胡洪营和李鑫,2010). 离心法是快速、可靠的分离采收方法. 但由于其极高的能耗和投资运行成本,在目前技术条件下并不具备大规模工程应用的潜力(Christenson and Sims, 2011). 过滤法仅在分离丝状藻时能耗和成本较低; 对于非丝状藻极易形成膜污染,能耗和运行成本很高,不能满足高效、低成本采收的要求(Christenson and Sims, 2011; Milledge and Heaven, 2013). 气浮法仅适用于采收单细胞藻类,在污水混合培养的条件下不能普遍适用; 此外,由于要产生大量的微小气泡,其投资和运行成本/能耗亦很高,甚至可能高过离心法(Milledge and Heaven, 2013). 直接重力沉降法是成本最为低廉的分离、采收方法. 但其耗时长,分离效果和可靠性最差(Christenson and Sims, 2011).
絮凝法是分离水中粗分散和胶体物质应用最为广泛的方法,在20世纪80年代就已经用于微藻的分离采收(Lavoie et al., 1984). 悬浮藻液经絮凝后能实现高效重力沉淀分离; 分离的藻细胞能直接被截留在反应器内,达到维持高生物量和保障出水水质的目的. 从单纯的藻细胞采收角度来说,絮凝法是处理大量稀藻液时最为经济、可行的方法. 虽然藻细胞经絮凝沉淀后还不能直接达到工业应用的要求,但已能显著降低后续浓缩过程的能耗和成本. 因此,絮凝法已被视为实现微藻大规模分离采收的最佳方法(Milledge and Heaven, 2013). 根据是否需要添加絮凝剂可分为“外加絮凝剂法”和“自发性絮凝法”两大类. 其中,外加絮凝剂法根据所使用的絮凝剂种类又可分为无机絮凝剂法、有机高分子絮凝剂法和生物絮凝剂法. 自发性絮凝按照发生机理可进一步分为高pH诱导的自发性絮凝和胞外聚合物(EPS)引起的自发性絮凝(Christenson and Sims, 2011; Pragya et al., 2013; Vandamme et al., 2013).
本文将从对微藻表面特性和絮凝机理的简要介绍出发,系统总结各种絮凝分离方法的研究应用现状,进而对各种方法进行综合比较,以期最终明确微藻絮凝分离的发展方向.
2 微藻表面特性和絮凝机理(Surface propertites of microalgae and flocculation fundamentals) 2.1 微藻悬浮液聚集稳定性的理论框架扩展DLVO(XDLVO)理论是胶体化学中描述胶体稳定性的经典理论之一,已成功应用于描述活性污泥系统微生物细胞间的粘附聚集(絮凝)过程(Bos et al., 1999; 刘晓猛,2008). 最近研究证实,该理论同样适应于描述微藻悬浮液中藻细胞的聚集过程(Ozkan and Berberoglu, 2013a,b,c). 在XDLVO理论中,胶粒间的相互作用主要考虑了以下3种非共价键的相互作用力:①范德华力(Lifshitz-van der Waals interaction),它是色散力、极性力和诱导偶极力之和; ②静电力(Electrostatic interaction),源自胶粒表面所带电荷的静电相互作用; ③Lewis酸-碱水合作用力(Lewis acid-base interaction),源自极性组分间的电子转移. 胶粒间的总表面位能(GTOT(d))为以上作用力的位能之和:
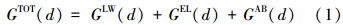
式中,GLW(d)为范德华作用力位能,GEL(d)为静电作用力位能,GAB(d)为Lewis酸-碱水合作用力位能.(d)表示作用力的大小和性质为胶粒间距的函数. 理论上,GTOT(d)>0则胶粒间相互排斥,处于聚集稳定状态; GTOT(d)<0则胶粒相互聚集(刘晓猛,2008; Ozkan and Berberoglu, 2013a). 典型的总位能曲线一般包含两个低位穴能(胶粒间距由远及近分别为第二低位穴能Em2和第一低位穴能Em1),两者之间存在一斥力能峰(Eb). 当胶粒相互靠近,到达第二低位穴能点(Em2)时,胶粒间处于一种可逆的粘附状态;外界条件稍有变化则粘附的胶粒又将相互分离,是一种不牢固的粘结状态. 只有胶粒的动能足够大,足以克服斥力能峰到达第一低位穴能(EEm1)时才能形成牢固的粘结状态,即发生絮凝(陈宗琪等,2001; 刘晓猛,2008).
2.2 藻细胞表面特性与聚集稳定性决定总表面位能(式(1))的3种基本作用力中,范德华力一般表现为引力,其大小取决于胶粒间距、单位体积内的粒子数量和和粒子的极化率等. 而静电力和Lewis酸-碱水合作用力的性质和大小则取决藻细胞的表面电势和亲/疏水性等表面特性.
2.2.1 表面电势藻细胞表面覆盖着一层复杂的EPS,其主要成分为碳水化合物(EPSC)和蛋白质(EPSP),其他成分包括腐殖质(Humus-like Substances)、核酸(Nucleic Acids)、糖醛酸(Uronic Acids)等. 这些成分导致藻细胞表面富集了大量羧基(—COOH)和氨基(—NH2)等功能团(De Philippis et al., 2001; Liu and Fang, 2003;Olguín,2012; Ozkan and Berberoglu, 2013b). 这些功能团随体系pH不同能接收或失去质子(H+),由此形成表面电荷及电势. 例如: 当体系处于低pH条件时,羧基和氨基都将接收H+(质子化,protonation),形成正的表面电荷;相反,当体系处于高pH条件时,羧基将失去H+(去质子化,deprotonation),形成负的表面电荷;在特定pH条件下,可以形成羧基失H+而氨基得H+的情况,表面净电荷为零,即等电点. 对于微藻,其等电点一般在pH=3. 而实际微藻培养系统的pH一般在7以上. 所以,藻细胞一般带负电(Ozkan and Berberoglu, 2013b),即式(1)中的静电作用力项表现为斥力.
胶粒表面电势无法直接测量,只能测量出胶粒的Zeta电位后通过计算间接得出. Zeta电位是胶粒双电层结构中滑动面与水溶液之间的电位差,是表征分散体系稳定性的重要指标. Zeta电位绝对值越高,胶粒之间的排斥力越大,体系越稳定(Vandamme et al., 2013). 实际培养条件下藻类的Zeta电位一般在-35~-15 mV之间(Ozkan and Berberoglu, 2013b). 因此,藻细胞间的静电斥力一般较大,是藻细胞在水溶液中保持聚集稳定性的主要原因.
2.2.2 亲/疏水性藻细胞的表面亲/疏水性决定了式(1)中Lewis酸-碱水合作用力的性质和大小,具体有如下规律: 疏水性藻细胞间的Lewis酸-碱水合作用力表现为引力; 亲水性藻细胞间的Lewis酸-碱水合作用力表现为斥力; 亲水和疏水藻细胞间的Lewis酸-碱水合作用力性质则取决于藻细胞的相对亲/疏水程度,可为引力或斥力; 亲/疏水程度越高,Lewis酸-碱水合作用力的值越大(Ozkan and Berberoglu, 2013c). 以上规律可通俗地理解为疏水细胞在水溶液中将受到水分子的“排斥”作用,因而细胞间有相互团聚(吸引)的趋势; 而亲水细胞则各自受到水分子的“吸引”,因而细胞间有分散在水溶液中的趋势(相互排斥)(Jorand et al., 1998). 藻细胞间的静电斥力一般大于范德华引力,因此在没有外加絮凝剂消除静电斥力的情况下,Lewis酸-碱水合作用力(表面亲/疏性)的性质和大小对微藻悬浮液的聚集稳定性就具有决定性的影响,例如: 亲水性藻细胞间的Lewis酸-碱水合作用力为斥力,因此该类藻细胞悬浮液总是能保持聚集稳定性; 只有Lewis酸-碱水合作用力为引力时(疏水藻细胞之间及特定亲水-疏水藻细胞组合),微藻悬浮液才有可能发生絮凝(Ozkan and Berberoglu, 2013c). 细胞亲/疏水性取决于其表面功能团: 表面富含长链烃类的微藻种属(如,葡萄藻属)表现为疏水性,因为长链烃类主要包含甲基和亚甲基等疏水基团,而羟基和羧基等亲水基团只占很小一部分; 表面富含糖醛酸、中性糖和葡糖胺等成分的微藻种属(如小球藻)则表现为亲水性,因为这些成分能形成大量羟基、羧基和氨基等亲水基团(Ozkan and Berberoglu, 2013a,b).
2.3 絮凝机理根据上述XDLVO理论,微藻絮凝的基本原理就是要通过降低/消除静电斥力(Zeta电位),使Lewis酸-碱水合作用力表现为引力等措施消除/降低藻细胞之间表面能的排斥能峰,使藻细胞能相互靠近到达第一低位穴能,从而紧密地粘结在一起形成絮体. 其中,外加无机絮凝剂的主要作用机理就是中和藻细胞表面的电负性,降低/消除静电斥力(Vandamme et al., 2013; 严煦世和范瑾初,1999). 外加高分子有机絮凝剂则主要通过吸附架桥原理起作用: 链状高分子物质(少数情况也可能是无机絮凝剂形成的大胶粒)在静电引力、范德华力和氢键力的作用下,一端吸附了某一胶粒后,另一端又吸附了另一胶粒,从而把不同的胶粒连接起来而形成絮体(图 1). 生物絮凝剂和EPS诱导的自絮凝则可能是通过Lewis酸-碱水合作用力中的疏水引力及吸附架桥原理的综合作用实现絮凝. 最后,投加絮凝剂形成的沉淀物和絮体等还可通过网捕和卷扫等物理作用进一步促进藻细胞的絮凝沉降.
 |
| 图 1 吸附架桥作用示意图(Banerjee et al., 2013) Fig. 1 Schematic of bridging mechanism(Banerjee et al., 2013) |
以铁盐和铝盐为代表的多价金属盐类和聚合金属盐类是传统水处理中应用最为广泛的絮凝剂,也是微藻絮凝中应用最早的外加絮凝剂. 典型絮凝剂包括:硫酸铝,硫酸亚铁,氯化铁,聚合氯化铝,聚合硫酸铝,聚合硫酸铝铁,聚合氯化铝铁,聚合硫酸氯化铝铁等(Sanyano et al., 2013; 张亚杰等,2010; 雷国元等,2007).
金属盐类絮凝剂主要是通过电性中和作用破坏藻细胞的聚集稳定性. Al3+、Fe3+等游离阳离子及其各种带正电荷的水解产物能中和藻细胞表面所带的负电荷,从而促进藻细胞碰撞聚集形成絮体,发生絮凝沉淀.此外,Al3+,Fe3+等金属盐还能形成[Al(OH)3]n、[Fe(OH)3]n等聚合体,以吸附架桥形式作用于藻细胞. 在特定pH下,这些金属盐类还可形成大量Al(OH)3及Fe(OH)3等沉淀物,以网捕卷扫作用促进微藻的絮凝沉降. 聚合金属盐类絮凝微藻的主要机理则是吸附架桥作用,同时也有电性中和及网捕卷扫作用(Vandamme et al., 2013; 王九思等,2002).
3.1.2 无机絮凝剂的絮凝效果和影响因素表 1总结了几种典型无机絮凝剂在微藻分离采收中的絮凝条件及效果. 由于各研究采用了不同的计量基准(如,生物量以细胞个数 · mL-1或mg · L-1计)及不同的藻种和絮凝条件,无法直接进行横向比较,但仍可以总结出一些要点:①铝盐、铁盐等多价金属絮凝剂在合适的条件下都可有效絮凝(>80%)常见的微藻种属;对于典型的稀藻液(浓度~0.5 g · L-1),药剂投加量一般要达几百mg · L-1藻液. ② 铝盐比铁盐的絮凝效率更高;金属氯化物比金属硫酸盐的絮凝效率更高;这反映在达到类似絮凝效果,铝盐和金属氯化物的投加量更小且所需絮凝时间更短. ③聚合金属盐类比非聚合金属盐类的混凝效率高,且在更广的pH值范围内有效(Jiang et al., 1993; Molina Grima et al., 2003; Papazi et al., 2010).
| 表 1 无机絮凝剂在微藻分离采收中的絮凝条件与效果 Table 1 Flocculation conditions and performance of inorganic flocculants in microalgae separation & harvesting |
影响无机絮凝剂絮凝效率的因素主要有:絮凝剂的种类、pH、藻液浓度和投加量等. 絮凝剂种类对絮凝效果的影响在上文已有所论述. 一般而言絮凝剂所带的电荷密度越高,絮凝效果越好(Garzon-Sanabria et al., 2013);这正是铝盐、铁盐的絮凝效果要远远好于钙、镁和铵离子的原因所在(Papazi et al., 2010). 此外,絮凝剂的水溶性也对其絮凝效果有显著影响,如:氯离子的水溶性好于硫酸根,所以金属氯化物的絮凝效果要好于金属硫酸盐.水溶性效应还可以解释摩尔质量对絮凝效果的影响,如:尽管Fe3+和Al3+ 一样带正3价电荷,但铁离子的摩尔质量大于铝离子;摩尔质量越大,水溶性越差;所以铁盐的絮凝效率要低于铝盐(Papazi et al., 2010). 铝盐的絮凝效果对pH高度敏感,最佳pH为4~5,这是因为在此pH条件下铝的水解产物以带正电的多核羟基配合物形式存在且最稳定;中性条件下,铝的水解产物以Al(OH)3沉淀为主;pH>8.5时,水解产物将以带负电的[Al(OH)4]-为主,无法形成有效絮凝(严煦世和范瑾初,1999). 一定范围内絮凝效果与絮凝剂的投加量成正比; 但过量投加会使胶粒吸附过多的反离子,重新带电而再次稳定; 因此絮凝剂使用存在一个最佳投加量,这是絮凝分离中早已得到深入分析的典型现象(严煦世和范瑾初,1999). 但最佳投加量随藻液浓度的变化规律文献中还存在不统一之处:一般而言所需絮凝剂的投加量随藻液浓度的升高而线性增加,这符合电性中和的絮凝机理(Chen et al., 2013; Wyatt et al., 2012). 然而,Garzon-Sanabria等在使用氯化铝絮凝微绿球藻(Nannochloris oculata)时发现,当藻液浓度很高时,达到同样絮凝效果的投加量却数倍地低于稀藻液(Garzon-Sanabria et al., 2013). Wyatt等在用氯化铁絮凝小球藻(Chlorella zofingiensis)时也得出了类似的结论(Wyatt et al., 2012). 其原因很可能是在高藻液浓度时形成了显著的网捕和卷扫作用. 最后,最新研究表明微藻代谢产生的有机物(Algogenic Organic Matter,AOM)对絮凝过程有显著的抑制作用,其存在将成倍地增加絮凝剂投加量(Garzon-Sanabria et al., 2013; Vandamme et al., 2012b),这将显著增加絮凝成本并对藻细胞的后续加工利用造成负面影响.
3.2 有机高分子絮凝剂法 3.2.1 种类与作用机理有机高分子絮凝剂在微藻分离采收中亦很早便得到了应用(Bilanovic et al., 1988). 目前商业化的有机高分絮凝剂主要为人工合成,以聚丙烯酰胺(Polyacrylamide)为代表. 近年来,天然高分子有机絮凝剂,如,壳聚糖(Chitosan)、阳离子淀粉(Cationic Starch)和纤维素等得到了越来越多的关注(Lam and Lee, 2012). 有机高分子絮凝剂的作用机理主要为吸附架桥作用. 因藻细胞带负电的表面特性,高效的高分子絮凝剂必须为阳离子型的. 阴离子及非离子型的聚合高分子单独使用时不能使微藻发生有效絮凝(Chen et al., 2011). 除架桥作用外,阳离子型高分子絮凝剂还可能局部逆转藻细胞表面的电负性,使其某些部位带负电而另一部位带正电;从而使不同的藻细胞能直接通过静电引力结合在一起,形成所谓的静电互补聚集(Electrostatic Patch Aggregation)(Vandamme et al., 2013). 以下介绍几种代表性的有机高分子絮凝剂.
a. 聚丙烯酰胺
聚丙烯酰胺分子量在400~2000万之间,具有阳性基团(—CONH2). 该基团既是亲水基团,又是吸附基团,所以能对微藻产生吸附电中和及架桥作用. 除桥连作用外,聚丙烯酰胺还有包络作用. 发生桥连和包络的高分子能形成三维网状结构,通过卷扫网捕作用使微藻沉降分离.
b. 壳聚糖
壳聚糖是对甲壳素进行脱乙酰基而得到的,是少数阳离子型的天然高聚物(Vandamme et al., 2013). 其结构单元是2-氨基-2脱氧葡萄糖,通过β-1-4糖苷键连接起来(图 2). 在酸性条件下,壳聚糖分子链上所带的大量氨基以带正电荷的胺离子形式存在,能中和藻细胞的电负性,同时借助高分子链的吸附架桥作用使藻体絮凝沉降. 当溶液呈现碱性时,壳聚糖表面所带胺基非离子化或呈弱负电性,从而降低了其絮凝效率(李若慧等,2012).
 |
| 图 2 壳聚糖结构(李若慧等,2012) Fig. 2 Structure of chitosan |
c. 阳离子淀粉
阳离子淀粉是在淀粉骨架中引入季铵基团,这样就使得淀粉呈正电性. 又因淀粉分子固有的聚合结构,使阳离子淀粉具有电性中和及吸附架桥的双重作用(Anthony and Sims, 2013). 阳离子淀粉和壳聚糖一样,也具有无毒、无污染、可生物降解的特点. 与壳聚糖比较而言,阳离子淀粉原料价格更低,更容易获得. 最为显著的是其季胺基团不受pH的影响,从而使其可在很宽的pH值范围内适用(Vandamme et al., 2010; Vandamme et al., 2013).
3.2.2 絮凝效果与影响因素表 2列出了几种典型有机高分子絮凝剂在微藻分离采收中的絮凝条件与效果. 表 2显示,聚丙烯酰胺虽然是水处理中应用最成熟的高分子絮凝剂,但其对微藻的絮凝效率却并不理想; 在相对较高的投加量下(20~80 mg · L-1藻液)也仅能实现50%左右的絮凝效果. 这可能是因为其电荷密度较低所致(薛蓉等,2012). 因此对聚丙烯酰胺进行改性是一个重要的研究方向,即,通过在聚丙烯酰胺上引入胺类分子,生成季胺型阳离子以进一步提高絮凝效率和适用范围(秦丽娟和陈夫山,2004). 此外,Chen等认为聚丙烯酰胺的高分子链展开程度不足是导致其絮凝效果不佳的原因.因此,从结构上对聚丙烯酰胺进行改进,增强高分子链的展开程度也是强化其絮凝效果的方向之一(Chen et al., 2013).
| 表 2 有机高分子絮凝剂在微藻分离采收中的絮凝条件及效果 Table 2 Flocculation conditions and performance of organic flocculants in microalgae separation & harvesting |
壳聚糖和阳离子淀粉对淡水藻类都有非常高的絮凝效率: 一般在10~30 mg · L-1藻液的投加量下就可以达到80%以上的絮凝效果; 对于个别藻种甚至在1 mg · L-1左右的投加量下就能达到90%以上的絮凝效果. 这比达到同样絮凝效果的无机絮凝剂投加量(表 1)要低一个数量级以上. 其原因主要是高分子絮凝剂具有显著的吸附架桥作用,因此可以在藻细胞负电性远未被中和的情况下(Zeta电位<<0)就实现高效絮凝(Anthony and Sims, 2013).
影响高分子絮凝剂絮凝效果的主要因素有:摩尔质量、电荷密度、投加量、藻细胞浓度、离子强度/盐度、pH和搅拌强度等(Chen et al., 2011; Molina Grima et al., 2003). 摩尔质量较高的高分子絮凝剂具有更多的吸附架桥结合点,因此一般具有更好的絮凝效果. 电荷密度高的高分子絮凝剂具有更强的电性中和能力; 此外,高的电荷密度还有助于高分子链的充分展开,增强架桥能力. 投加量不足,絮凝效果不充分; 但投加过量,又会对胶粒起到稳定保护作用; 这与无机絮凝剂中最佳投加量的概念一致(Ahmad et al., 2011; Vandamme et al., 2010). 高的藻细胞浓度使颗粒间的碰撞更加频繁,在一定范围内将促进絮凝作用. 与无机絮凝剂相比,阳离子型高聚物的一个特点是,高的离子强度/盐度对其絮凝效果有显著的抑制作用.这是因为在高离子强度/盐度情况下,阳离子型高聚物有团聚在一起的趋势,架桥作用将显著减弱(Molina Grima et al., 2003). 这使其在采收海洋微藻时受到限制(Bilanovic et al., 1988; Chen et al., 2011). 壳聚糖一般在酸性条件下絮凝效果才显著,这往往超出了微藻培养体系的正常pH范围,从而限制了其应用(翟玥等,2009). 而阳离子淀粉基本不受pH影响,在pH 5~10的范围内都能维持+15 mV左右的Zeta电位,具有普遍的适用性(Anthony and Sims, 2013; Vandamme et al., 2010). 低速搅拌对形成大的絮体有利; 过强的搅拌将破坏已形成的絮体(Ahmad et al., 2011).
3.3 生物絮凝剂法生物絮凝剂(Bioflocculant)是近几年微藻絮凝的研究热点之一. 生物絮凝剂一般是指微生物代谢活动中产生的具有絮凝效果的胞外聚合物(EPS). 细菌、真菌和放线菌都是能产生生物絮凝剂的常见微生物(Lam and Lee, 2012). 生物絮凝剂在微藻采收中的具体应用方式主要包括以下几种:①投加絮凝微生物的混合培养液(微生物细胞+培养液);②菌-藻混合培养(需在微藻培养系统添加有机碳源);③絮凝微生物的胞外抽取液(离心后的上清液)作为絮凝剂;④分离纯化后的胞外提取物作为絮凝剂;⑤直接投加絮凝微生物细胞作为絮凝剂.
表 3总结了各种生物絮凝剂在微藻絮凝分离中的应用情况. 表 3显示,生物絮凝剂的絮凝效率亦很高,一般在投加量为10~30 mg · L-1藻液时便可达到>80%的絮凝效果,作用明显好于无机絮凝剂(表 1). 但在絮凝效果的影响因素上却存在不少相互矛盾之处. 一些研究显示絮凝效果会随pH升高明显加强(Oh et al., 2001; Powell and Hill, 2013),但也有研究显示絮凝效果基本不受pH影响(Zheng et al., 2012);多数研究表明,多价阳离子能显著促进絮凝甚至是形成絮凝的必要条件(Kim et al., 2011; Oh et al., 2001; Powell and Hill, 2013),但在少数研究中多价阳离子对絮凝效果基本没影响(Wan et al., 2013);Lee等显示絮凝微生物利用不同碳源产生的絮凝剂絮凝效果基本一致(Lee et al., 2009),而Wang等以不同碳源为底物产生的生物絮凝剂絮凝效果迥异(Wan et al., 2013). 这些矛盾可能是各种生物絮凝剂在种类、组成及絮凝机理上的不同而导致的.
| 表 3 各种生物絮凝剂在微藻分离采收中的应用 Table 3 Application of various bioflocculants in microalgae separation & harvesting |
在絮凝机理上,绝大部分文献中只考虑了静电作用力,把生物絮凝归结于吸附架桥作用,具体又可细分为以下两种机制:①长链EPS在不同部位吸附多个带负电的藻细胞形成架桥作用(Bridging); ②短链EPS在局部逆转藻细胞的电负性,从而形成所谓的静电互补效应(Patching)(图 3)(Salim et al., 2011). 这一理论的基础是将生物絮凝剂(EPS)默认为阳离子型高聚物. 但如2.2.1节所述,EPS在中性及碱性条件下本身是带负电的. 那么,生物絮凝剂是如何实现阳离子化的呢?一个相对成熟的理论为二价阳离子架桥理论(Divalent Cation Bridging(DCB)Theory)(Powell and Hill, 2013; Sobeck and Higgins, 2002; Surendhiran and Vijay, 2013),可结合图 4说明如下: EPS本身具有多个带负电的活性部位,这使其能强烈吸附环境中的二价阳离子. 被吸附的二价阳离子所带正电荷只被EPS中和了一半,所以能另外吸附一个带负电的藻细胞. 由此,多个藻细胞通过二价阳离子的架桥作用连接在EPS上,形成大的絮体. 这一理论能很好地解释为什么多价阳离子对生物絮凝具有显著的强化作用甚至是絮凝形成的必要条件. 这一理论也能解释絮凝效果随pH升高而增强的现象: pH升高,EPS电负性增强,吸附二价阳离子的能力增强,所以架桥作用增强. 但如上所述,在部分研究中阳离子的存在对絮凝效果根本就没有影响(Wan et al., 2013). 这就无法用DCB理论解释了. DCB理论最根本的缺陷在于,理论上应该是所有带负电的EPS都能通过二价阳离子的架桥作用形成絮凝. 但这显然是与事实不符的. 这一问题将在4.3节部分得到进一步讨论.
 |
| 图 3 生物絮凝剂的絮凝机理(Salim et al., 2011) Fig. 3 Flocculation mechanisms of bioflocculants(Salim et al., 2011) |
 |
| 图 4 二价阳离子架桥原理 Fig. 4 Divalent cation bridging(DCB)theory |
微藻有时能在没有添加任何絮凝剂的情况下发生絮凝,这一现象被称为自发性絮凝(Auto-flocculation)(Besson and Guiraud, 2013; Christenson and Sims, 2011; Gonzalez-Fernandez et al., 2013). Golueke和Oswald(1965)首先描述了这一现象. 他们发现:藻类塘中的微藻在温度较高且光线充足的时候能自然地形成絮体(Golueke and Oswald, 1965). 此后,很多学者都证实了类似现象的存在并开展了相关研究(Besson and Guiraud, 2013). 目前形成的基本共识为,微藻自发性絮凝是由两种不同机理引发的(Besson and Guiraud, 2013; Christenson and Sims, 2011; Gonzalez-Fernandez et al., 2013):
(1)在高pH下,钙、镁等离子形成带正电的沉淀物,起到电性中和作用从而引发絮凝. 文献中的自发性絮凝一般即指此类. 高pH可以是由微藻光合作用消耗水中无机碳(Inorganic Carbon,IC)自然形成,也可通过人工添加碱性物质(石灰,氢氧化钠等)而形成. 严格来说,只有前者才符合自发性絮凝的定义. 但考虑到两者的实质都是形成带正电的沉淀物,本文在此将两者一并纳入高pH诱导的自发性絮凝范畴.
(2)部分藻种在其生理活动中能产生大量具有絮凝作用的胞外聚合物,起到生物絮凝剂的作用(参见3.3节部分)从而引发絮凝. 高效藻类塘中常见的集星藻属(Actinastrum)、微芒藻属(Micractinium)、栅藻属(Scenedesmus)、空星藻属(Coelastrum)、盘星藻属(Pediastrum)及胶网藻属(Dictyosphaerium)等常通过该机理形成大的群落结构(50~200 μm)而得以自然沉降(Pragya et al., 2013). 文献中常将其归为生物絮凝一类(参见3.3节部分). 因为该情况下的生物絮凝剂为藻细胞自身所产生,本文在此也将其纳入自发性絮凝并定义为EPS引起的自发性絮凝.
4.2 高pH诱导的自发性絮凝如上所述,高pH诱导的自发性絮凝其实质为所生成的带正电沉淀物的电性中和作用. 因此,诱导此类自发性絮凝的关键就是明确在微藻正常培养条件下所能形成的沉淀物种类和性质. 污水中一般含有大量的钙、镁、碳酸根和磷酸根等离子,在碱性条件下比较容易形成的沉淀物主要包括:磷酸钙,氢氧化镁和碳酸钙. 大量研究证实(Beuckels et al., 2013; Elmaleh et al., 1991; Gonzalez-Fernandez and Ballesteros, 2013; Smith and Davis, 2012; Sukenik and Shelef, 1984),碳酸钙本身带负电,最多只能通过网捕和卷扫作用实现非常有限的絮凝效果. 磷酸钙和氢氧化镁带正电,理论上都可诱导自发性絮凝. 但两者形成的具体条件差别较大,以致文献中的结论常常容易引起误解. 为此,以下予以详述.
Sukenik和Shelef(1984)首次定量、系统性地研究了高pH下的微藻自絮凝现象,结论性认为,磷酸钙是诱导自絮凝的关键沉淀物(Sukenik and Shelef, 1984). 他们的试验包括两部分:①户外培养系统(二形栅藻,Scenedesmus dimorphus)的自絮凝试验: 采用间歇培养,周期为8 d; 前7 d连续通入CO2使pH维持在7.0; 第8天停止CO2供应和搅拌,监测絮凝效果和相关水质参数(pH,TSS,PO43--P,Ca2+,Mg2+,碱度)的变化情况. ②室内絮凝试验(Jar Test): 取处于对数增长期的二形栅藻和小球藻置于和户外培养系统相同的培养基,调节pH为2.5~10.5进行絮凝试验(80 r · min-1 1 min,30 r · min-1 15min,沉淀15 min),监测絮凝效果和相关水质参数变化. 户外培养试验结果表明,系统停止CO2供应和搅拌24 h后,体系pH迅速升至8.9,PO43--P、Ca2+同步降低,高效絮凝(96%)形成,而Mg2+维持不变(表 4). 由此初步证明,自发性絮凝是由高pH下的磷酸钙沉淀诱导的. 室内絮凝试验结果显示:pH 5.0~7.5时无絮凝发生. pH>8.5后形成了高效絮凝(~98%),与户外试验结果一致. 进一步试验显示,在PO43--P为6.2 mg · L-1的情况下,Ca2+(2.0 mmol · L-1)在pH≥8.5时就能生成沉淀并引发絮凝;而Mg2+(2.0 mmol · L-1)要在pH≥10.5后才能生成沉淀进而引发絮凝. 如果没有PO43--P存在,则Ca2+在整个碱性范围内都不能引起絮凝.
| 表 4 自絮凝前后的水质参数变化(Sukenik et al., 1984) Table 4 Changes of water quality parameters before and after autoflocculation |
这些实验结果充分证明,在微藻生长的正常pH范围内(8~10),磷酸钙是诱导自絮凝的关键沉淀物. 更具工程意义的是,Sukenik和Shelef(1984)发现自絮凝的临界pH值(絮凝效果达50%的pH)将随PO43-(表 5)和Ca2+(未显示)浓度的上升而下降; 在含高浓度Ca2+和PO43- 的培养液中,微藻自絮凝甚至可以在中性条件下就发生. 这说明当以含磷较高的市政污水为底物时,可相对较容易地在微藻生长的正常pH值范围内,通过生成磷酸钙沉淀同步实现自发性絮凝和高效除磷.
| 表 5 小球藻在不同磷浓度下的自絮凝临界pH值(钙浓度为100 mg · L-1)(Sukenik & Shelef,1984) Table 5 Critical pH values of chlorella autoflocculation depending on phosphate concentrations(calcium: 100 mg · L-1) |
然而,除少数学者继续证实磷酸钙在诱导微藻自絮凝中的核心作用外(Beuckels et al., 2013; Moutin et al., 1992),大部分研究都显示氢氧化镁才是诱导自絮凝的关键沉淀物(Besson and Guiraud, 2013; Castrillo et al., 2013; Chen et al., 2013; Elmaleh et al., 1991; Schlesinger et al., 2012; Sirin et al., 2012; Smith and Davis, 2012; Vandamme et al., 2012a; Wu et al., 2012; Yahi et al., 1994),代表性研究如下:
(1)Vandamme等(2012a)在研究小球藻自絮凝时发现,在pH≤10.5时均无自絮凝发生; 当pH调至11时,实现了75%的絮凝效果; pH≥11.5后絮凝效果显著,达95%以上(表 6). 此时若加入EDTA(最终浓度0.5M)掩蔽Ca2+、Mg2+等二价离子,则絮凝效果急剧下降至<20%. 这初步证明了钙/镁沉淀在诱导自絮凝中的关键作用. 进一步研究表明,若溶液中只存在Ca2+(1~100 mg · L-1),在高pH下(10.5~12)确实也能形成沉淀物,但始终不能形成有效絮凝(<20%). 而当溶液中存在Mg2+(1.8 mg · L-1)时,在pH=10.5就可实现25%的絮凝效果; 在pH=12时絮凝效果显著增至85%. 当Mg2+浓度≥3.6 mg · L-1后,pH=10.5就可达90%以上的絮凝效果(注:该实验条件(藻细胞浓度等)与表 6不同,故临界pH和絮凝效果有所差异)(Vandamme et al., 2012a).
| 表 6 以氢氧化镁沉淀诱导的自发性絮凝 Table 6 Autoflocculation induced by magnesium hydroxide precipitate |
(2)Sirin等(2012)在研究三角褐指藻的自絮凝时发现,在pH=10.5时絮凝后溶液中的Mg2+下降了约16%,而Ca2+基本不变; 当pH增至11时,溶液中基本已没有Mg2+残留,而Ca2+仅降低了14%(Sirin et al., 2012). 这说明在pH=10.5~11时生成的沉淀物主要为镁沉淀物.
(3)Smith和Davis(2012)发现,Mg2+、Ca2+和CO32-离子中,只有Mg2+(9.6 mmol · L-1)存在时才能在高pH下(>10)实现高效絮凝;而Mg2+缺乏时(Ca2+、CO32-均为9.6 mmol · L-1),即使Ca2+沉淀了75%也不能形成有效絮凝(Smith and Davis, 2012).
以上实验结果似乎均已证明,高pH下只有Mg沉淀物才能有效诱导自絮凝. 然而,仔细考察这些研究中的特定离子浓度(表 6)就可发现,其Ca2+、PO43-离子中至少有一项或两项的浓度均较低(与表 5对比),因此,在pH=8~10的范围内不能形成大量磷酸钙沉淀; 只能在更高pH下(>10.5)生成Mg(OH)2沉淀时才能诱导自絮凝. 综合以上所有结果,可以得出如下结论: 磷酸钙和氢氧化镁沉淀都可有效诱导出自发性絮凝. PO43--P、Ca2+浓度均较高时,磷酸钙沉淀在相对较弱的碱性条件下(pH 8~10)就可生成并诱导出显著的自絮凝;PO43--P/Ca2+浓度较低时,则需进一步提升pH至10.5以上,产生氢氧化镁沉淀后才能诱导出自絮凝.
4.3 胞外聚合物(EPS)引起的自发性絮凝20多年以前,籍EPS形成自发性絮凝的藻种便在微藻分离采收中得到重视(Olguín,2003). Borowitzka和Borowitzka(1988)分离出了蓝藻门的一株胶鞘藻. 它能分泌出大量具有絮凝作用的EPS,其主要成分包括多聚糖、脂肪酸和蛋白质(Borowitzka and Borowitzka, 1988). 此后,自絮凝藻种如鲍氏席微藻(Phormidium bohneri)(Dumas et al., 1998; Talbot and Delanoue, 1993)及丝状藻Chlorhormidium(Sérodes et al., 1991)等在污水处理中都得到了应用. 但时至今日,对此类自絮凝的研究总体来说还十分有限.
在实际应用上,一般思路为将自絮凝藻种投入非自絮凝藻种培养系统以实现絮凝分离. Salim等(2011)研究了淡水自絮凝藻种镰形纤维藻(Ankistrodesmus falcatus)、斜生栅藻(Scenedesmus obliquus)对小球藻(Chlorella vulgaris)絮凝作用及海洋自絮凝藻种四鞭片藻(Tetraselmis suecica)对富油新绿藻(Neochloris oleoabundans)的絮凝作用(Salim et al., 2011). 实验结果显示,四鞭片藻的絮凝效果最佳,可达70%左右;斜生栅藻次之,絮凝效果可达30%左右;镰形纤维藻絮凝效果最差,约20%. Guo等(2013)考察了自絮凝斜生栅藻藻株S. obliquus AS-6-1对非絮凝淡水藻株S. obliquus FSP-3、C. vulgaris CNW-11和海洋微藻N. oceanica DUT01的絮凝作用. S. obliquus AS-6-1对淡水藻株均取得了80%以上的良好絮凝效果;而对海洋微藻的絮凝效果则相对较差,低于60%(Guo et al., 2013). 以上实验结果表明,自絮凝藻株的絮凝效果会因目标藻种而异,这在实际应用中存在着很大的局限性.
目前对于微藻EPS诱导自絮凝的机理的研究更为有限,一般只是笼统地认为与生物絮凝剂的机理一致(参见3.3节部分). 在相互作用力上,绝大部分研究只考虑了基于DCB原理的静电作用力(Salim et al., 2011),只有最近极少数研究考虑了Lewis酸-碱水合作用力(Ozkan and Berberoglu, 2013a,b,c). 如3.3节所述,单纯地用DCB原理(图 4)解释藻细胞EPS的絮凝机理将存在很大的缺陷. 因为按此理论所有产生EPS的微藻种属都应该能通过DCB原理发生自絮凝,而实际上只有某些特定藻种产生的EPS才有絮凝作用. 一个可能的解释为:非絮凝藻种产生的EPS数量较少,架桥能力有限;而自絮凝藻种能产生大量EPS,所以絮凝效果显著. 另一个可能的解释为,非絮凝藻种和自絮凝藻种所产生的EPS在组成和性质上有所不同. 例如,Guo等(2013)发现,在EPS各组分中只有多聚糖为絮凝的活性成分(Guo et al., 2013). 因此,如果不同藻种产生的EPS中多聚糖的含量不同,那其絮凝效果就可能有显著差异. 但Guo等(2013)进一步发现:无论是投加Ca2+,还是用EDTA掩蔽Ca2+,对絮凝效果都没有影响,这就从根本上与DCB理论相悖了(Guo et al., 2013).
Ozkan和Berberoglu(2013a,b,c)则从藻细胞的亲/疏水性表面特性出发,在静电作用力的基础上增加了对藻细胞间Lewis酸-碱水合作用力的考察. 他们的研究结果表明: Lewis酸-碱水合作用力在XDLVO的3种基本作用力中最强,在微藻的自絮凝中具有关键作用. 当微藻悬浮液中存在适量疏水性较强的微藻种属(如,布朗葡萄藻)时,即使是电负性较大的亲水-疏水混合藻液也能形成絮凝; 如果只考虑范德华力和静电力,这一现象将无法得到解释(GTOT(d)= GLW(d)+ GEL(d)> 0,理论上不发生絮凝); 而考虑Lewis酸-碱水合作用力后,理论预测与实际观察得到了很好的吻合(GTOT(d)= GLW(d)+ GEL(d)+ GAB(d)< 0,理论预测为发生絮凝). 这些研究成果为理解EPS诱导自絮凝的机理和促进自絮凝效果提供了非常有前景的思路. 因此,后续无论对生物絮凝还是EPS诱导的自絮凝,都应在完整的XDLVO理论框架内考察范德华力、静电力和Lewis酸-碱水合作用力的综合作用. 对活性污泥EPS的研究发现: EPS中的蛋白质是形成疏水性的主因,而碳水化合物是形成亲水性的主因; EPS的数量和组成受生长阶段、底物水平等因素影响,对微生物絮凝有关键影响(Jorand et al., 1998; Liao et al., 2001; Liu and Fang, 2003). 而对藻细胞EPS各组分的产生、变化规律及其对表面特性影响的系统研究几乎还是空白. 这方面的研究无疑将为理解和调控微藻自絮凝提供非常有价值的信息.
4.4 影响因素自发性絮凝的发生机理决定了其影响因素. 对于高pH诱导的自发性絮凝,其根本因素为微藻光合作用提升pH所能到达的程度和所能形成带正电沉淀物的特定离子浓度. 对于EPS引起的自发性絮凝,其影响因素则更加复杂,理论上包括所有影响EPS产生和组成的因素.
4.4.1 光照光照是微藻生长繁殖的基本要素,对高pH和EPS诱导的自发性絮凝都具有重要影响. 首先,光照直接决定了微藻光合作用的程度. 光照越强,光合作用越充分,水中无机碳消耗越彻底,pH上升越高,越有利于高pH诱导的自絮凝发生. 其次,光照也是影响EPS产生的关键因子(Gonzalez-Fernandez and Ballesteros, 2013; Moreno et al., 1998; Rebolloso Fuentes et al., 1999). Moreno等(Moreno et al., 1998)发现光照强度由345 μmol · m-2 · s-1增加到460 μmol · m-2 · s-1后,鱼腥藻的EPS含量增加了4倍. Rebolloso-Fuentes等(1999)也发现,在较高的外部光照条件下,紫球藻EPS的含量显著增加. 因此,充分的光照是诱导自发性絮凝的有利因素.
4.4.2 特定离子Ca2+、Mg2+和PO43-等特定离子的浓度决定了沉淀物的种类和产生的临界pH值,对高pH诱导的自发性絮凝具有决定性影响. 从发生机理来看:PO43--P/Ca2+离子都大量存在时,在较弱的碱性条件下(pH 8~10)磷酸钙沉淀就可大量生成并成为自絮凝主导因素;PO43--P/Ca2+的其中之一浓度较低时,则需进一步提升pH至10.5以上,产生氢氧化镁沉淀后才能诱导出自絮凝(参见4.2节部分). 从絮凝效果来看:以上离子浓度越高,生成带正电的沉淀物越多,电性中和能力越强,絮凝就越充分. 因此,将上述特定离子维持在较高水平对实现高pH诱导的自发性絮凝至关重要.
4.4.3 温度和生长阶段温度对微藻EPS形成具有重要影响(Gonzalez-Fernandez and Ballesteros, 2013; Lupi et al., 1991; Moreno et al., 1998). 高温刺激EPS的形成,而低温下由于细胞新陈代谢降低EPS的形成受到抑制. 但EPS产生的最佳温度因藻种不同而异,如:布朗葡萄藻(Botryococcus braunii)在温度低于23 ℃几乎不分泌EPS,其最佳温度为30~33 ℃(Lupi et al., 1991);而鱼腥藻(Anabaena sp.)在30~35 ℃范围内EPS产量都很少,只有在40 ℃以上EPS才大量产生(Moreno et al., 1998).
微藻所处生长阶段对藻细胞密度、表面性质和EPS的产量及成分等都有显著影响(Gonzalez-Fernandez et al., 2013). Lavoie和de la Noüe发现,老龄化的(Aging)的藻细胞密度增大,易于沉淀(Lavoie and Delanoue, 1987). Zhang等发现,小球藻从对数增长期进入稳定期后,其表面电负性减弱,易于发生絮凝(Zhang et al., 2012). Lavoie和de la Noüe(Lavoie and Delanoue, 1987)、Zhang等(Zhang et al., 2012)和Salim等(Salim et al., 2013)都发现微藻在对数期EPS产量很少,而在稳定期或衰减期产量则显著增加. Salim等还进一步证实EPS的组成将随生长周期的不同而变化(Salim et al., 2013). 因此,与活性污泥类似,微藻处于稳定期或衰减期时自絮凝效果较好(Gonzalez-Fernandez et al., 2013; Salim et al., 2013). 在实际培养中可将微藻的生长阶段控制在稳定期或衰减期以促进自发性絮凝的形成.
4.4.4 底物水平N、P等营养元素的缺乏将刺激微藻EPS的生产(Gonzalez-Fernandez et al., 2013; Lee et al., 2009),这与细菌、真菌等微生物一致. 基于此,在运行中可采用高密度培养以获得较低的F/M值,以自然形成底物受限的工艺条件. 而微藻生长的另一重要底物——无机碳(IC)受限则将抑制EPS的生产(Cordoba-Castro et al., 2012; Gonzalez-Fernandez and Ballesteros, 2013). 如,Cordoba-Castro等发现斜生栅藻(Scenedesmus obliquus)的EPS生产随CO2的供给而增加:在高CO2供给条件下(4%),微藻的生长和EPS产量都最大;而CO2供给下降后EPS的生产也随之降低(Cordoba-Castro et al., 2012). 从强化EPS生产的角度来看,在实际运行中无疑应加强IC的供给. 然而,如Pragya等所指出,为强化基于高pH的自絮凝则应限制IC供给,以达到尽可能高的pH条件(Pragya et al., 2013). 因此,对IC的调控应权衡其对高pH和EPS两种自絮凝正反两方面的综合效应.
4.4.5 微藻种属无论是基于高pH的自絮凝还是基于EPS的自絮凝,其絮凝条件和效果都将随目标藻种不同而异(参见4.2节和4.3节部分). 这可能是由于藻细胞在表面特性和生理特性上的不同而导致的. 如,电负性较高的藻细胞需要更多带正电的沉淀物生成. 又如多细胞和大型丝状藻种比单细胞藻种更容易絮凝沉降. 在这方面需要综合考虑微藻种属的污水净化能力、藻细胞的利用价值等,选择性富集易于絮凝沉降的藻种.
4.4.6 溶解性有机物与外加混凝剂类似(参见3.1.2部分),水中溶解性有机物(DOM)对自絮凝也会产生显著的抑制作用. 这些DOM既可能是原水中带来的腐殖质,也可能是藻类代谢产生的有机物(AOM). Beuckels等(2013)表明,腐殖酸和藻酸盐将显著抑制磷酸钙诱导的自絮凝,而葡萄糖和乙酸等小分子却没有影响(Beuckels et al., 2013). 同样,Wu等(2012)发现AOM对氢氧化镁诱导的自絮凝有强烈的抑制作用,当AOM从零增加至70 mg · L-1时,小球藻的自絮凝效率从92%降低至7%(Wu et al., 2012). 其原因很可能是DOM将优先与Ca2+、Mg2+等离子结合,从而阻止了磷酸钙和氢氧化镁等沉淀物的产生. 另外,由于DOM本身带负电,因此会额外增加电性中和所需的絮凝剂用量. 鉴于此,Beuckels等(2013)指出,DOM的抑制作用很可能是很多实际情况下,磷酸钙/氢氧化镁等沉淀物的相关生成条件都已超过临界值,但自絮凝却没有发生的原因所在(Beuckels et al., 2013). 在这方面迫切需要更进一步的系统研究.
5 各种絮凝分离方法的比较与展望(Comparison and perspective of different flocculation methods)以铁盐和铝盐为代表的金属絮凝剂是各种絮凝方法中应用最为成熟的技术.其主要优点是药剂生产简单,絮凝条件容易控制,絮凝效果有保障. 但无机絮凝剂的用量一般很大(几百mg · L-1藻液),从而产生大量污泥. 再者,絮凝效果受pH影响较大,其最佳pH值很可能超出微藻培养系统的正常pH范围,且无机絮凝剂仅对部分微藻种属有效(Chen et al., 2011; Pragya et al., 2013). 最不利的效果是,金属盐类往往对藻细胞具有毒害作用: Chen等在使用硫酸铝和氯化铁絮凝栅藻时发现,当投加量较高时,藻细胞在24 h后全部死亡(Chen et al., 2013);Papazi等也发现铝盐虽然絮凝效果最好,但会引起藻细胞裂解(Papazi et al., 2010). 此外,金属盐类残留在藻细胞中还将对藻细胞的利用和最终处置造成不利影响(Chen et al., 2011; Vandamme et al., 2013). 因此,从微藻培养的角度来看,金属盐类絮凝剂并不是最佳的技术选择. 有鉴于此,无机金属絮凝剂似乎不可能成为微藻分离采收的主要发展方向.
与无机絮凝剂相比,有机高分子絮凝剂具有更高的絮凝效率(10~30 mg · L-1藻液),产生的污泥量小,能适用于更广泛的微藻种属(Christenson et al., 2011; Pragya et al., 2013). 其中,聚丙烯酰胺虽然是水处理中应用最成熟的高分子絮凝剂,但其对微藻的絮凝效果却不如壳聚糖、阳离子淀粉等天然高分子絮凝剂. 且其在使用中可能会释放出一定量具有强烈毒性的单体丙烯酰胺(Vandamme et al., 2013; 秦丽娟和陈夫山,2004),因此其应用前景有限. 天然高分子絮凝剂无毒,易生物降解,对微藻培养和藻细胞的后续利用基本无负作用,在微藻的分离采收中具有良好的应用潜力. 但天然高聚物中只有壳聚糖等少数是阳离子型的. 壳聚糖的絮凝效率很高,但其絮凝条件一般为酸性,超出了微藻生长的正常pH范围. 考虑到对大量藻液进行酸化所需投加的化学药剂用量,壳聚糖很可能在经济上不具备选择性(Vandamme et al., 2013). 阳离子淀粉在原料上可大量获取,价格低廉,投加量非常小(几mg · L-1藻液),絮凝效果优异且基本不受pH影响(Vandamme et al., 2010),具有良好的工程化应用潜力. 后续研究的重点应在于优化其阳离子化过程,以进一步提高其适用性和絮凝效率,并显著降低加工制造成本.
利用细菌、真菌等微生物生产的生物絮凝剂具有高效、无毒、可生物降解等优点. 但其各种利用方式都存在明显缺陷:1)直接投加微生物细胞或菌-藻共同培养有对微藻培养系统造成污染的风险;2)投加培养液、抽取液、提取物等方式需要一个微藻培养系统以外的单独培养体系,尤其是后两者还涉及到复杂的分离和加工问题,这无疑会增加利用难度和成本. 可见,生物絮凝剂一般所宣称的低成本优势可能在实践中难以能成为现实(Salim et al., 2011). 生物絮凝剂另一缺点是,某一生物絮凝剂可能只对某些特定藻种絮凝效果较好(Oh et al., 2001; Wan et al., 2013). 利用自絮凝藻种产生的生物絮凝剂不需要额外的培养体系,且无污染微藻培养之虞. 但自絮凝种属的生长速度一般低于非自絮凝种属,其污水净化能力和产油潜力也可能不如非絮凝藻种(Salim et al., 2011). 因此,控制自絮凝藻种在系统中的比例至关重要. 这就提出了在混合培养中进行种群控制的复杂要求. 与细菌、真菌等微生物产生的生物絮凝剂类似,自絮凝藻株的絮凝效果也将随目标藻种的不同而异. 理解自絮凝(EPS诱导)藻种的絮凝机理对促进其应用具有关键意义. 目前在这方面的研究还非常不足,基本上还处于对EPS的成分分析上. DCB理论虽然能解释很多实验现象,但存在不能解释为什么只有特定藻种才具有絮凝作用这一根本缺陷. 在此方面,由藻细胞亲/疏水性决定的Lewis酸-碱水合作用力是非常有前景的理论,应该成为后续研究的重点.
氢氧化镁沉淀虽然能有效诱导出自发性絮凝,但其形成一般要在pH>10.5以上. 而大部分微藻在pH>9时光合作用就会受到显著抑制甚至完全停止,所以微藻的自然生长很可能达不到氢氧化镁沉淀的生成条件(Spilling et al., 2011). 事实上,几乎所有基于氢氧化镁沉淀的自絮凝都是通过外加碱性物质达到所需pH值(表 6),这无疑会带来额外的成本. 高pH还可能对藻细胞造成严重损伤,例如,高产油藻种Skeletoma costatum在pH = 10. 2时絮凝率达 80%,但回收藻体中相当一部分细胞发生解体及胞内成分外泄,严重影响后续二十碳五烯酸(EPA)提取工艺(Blanchemain and Grizeau, 1999);螺旋藻(Spirulina platensis)在pH高于13时,细胞絮凝得又快又彻底,但此时藻细胞颜色发黄,表明其细胞已受到较为严重的“pH损伤”(曾文炉等,2003).此外,高pH值还很可能超出排放标准,需要再加酸调节到容许的范围. 因此,基于氢氧化镁沉淀的自絮凝只适用于单纯的微藻采收,对微藻污水处理系统来说并不是一个合适的选择.
基于磷酸钙沉淀的自絮凝无需任何额外投入,在微藻自然生长的pH范围内(8~10)就能形成,能同步实现除磷,对微藻活性和藻细胞的后续加工利用几乎没有不利影响. 因此,无论是从污水深度处理,还是从藻细胞的采收利用等角度来看都是最合适的分离采收方法之一. 尤其是随着磷酸盐浓度提高,其临界pH值将显著下降;而这在实际污水处理中恰恰是一个很容易控制的工艺条件. 鉴于此,可以提出以下几个强化基于磷酸钙沉淀自絮凝的思路:①在污水处理的主体工艺中取消强化生物除磷,为后续微藻处理系统保留高磷浓度;②采用高密度间歇培养方式,并完全或在反应周期末端取消外部CO2供给,以迅速且自然地形成高pH条件;③通过适当延长反应周期、强化光合作用(光照、底物浓度)等进一步促进pH的提升. 目前存在的问题是大部分研究都只是考察高pH下瞬时的絮凝效果. 如果微藻培养系统长期处于诱导自絮凝所需的高pH环境,几个非常值得关注的问题是:①微藻种群结构是否会发生显著变化?② 目标藻种能否维持优势?③ 微藻的生理特性(净化能力、油脂含量等)是否会发生改变?这些都需要进一步的系统研究来明确.
6 结论(Conclusions)用于微藻分离、采收的理想絮凝剂应该具有无毒(对藻细胞本身及环境)、低成本、广谱高效、不影响藻细胞后续利用等特点. 依此标准,金属盐类絮凝剂因投加量大、对藻细胞有毒性及影响藻细胞的后续利用等缺点不能成为微藻絮凝的主要发展方向. 有机高分子絮凝剂中聚丙烯酰胺对微藻的絮凝效果较差,可能释放有毒的丙烯酰胺单体,因此其应用前景有限. 壳聚糖絮凝效率较高,但适应的絮凝pH一般为酸性,超出了微藻培养的正常范围,具有很大的局限性. 而阳离子淀粉在原料上可大量获取,价格低廉,投加量非常小,絮凝效果优异且基本不受pH影响,具有良好的工程化应用潜力. 后续研究的重点应在于优化其阳离子化过程,以进一步提高其适用性和絮凝效率,并显著降低加工制造成本.
利用细菌、真菌等微生物生产的生物絮凝剂具有高效、无毒、可生物降解等优点. 但其各种利用方式都存在明显缺陷,例如,可能对微藻培养造成污染,需要额外的培养系统,涉及到复杂的分离纯化过程等. 此外,生物絮凝剂的絮凝机理和条件都还不甚明确. 因此,生物絮凝剂距工程化应用还存在很大距离. 同理,基于胞外聚合物的微藻自絮凝因絮凝机理复杂、絮凝条件不明确及涉及到复杂的种群控制要求等,可靠性较差,在微藻絮凝分离中可能只能起到锦上添花的作用.
基于氢氧化镁沉淀的自絮凝要求很高的pH值(>10.5). 这往往超出微藻正常生长所能达到的pH值范围,需要额外投加碱性物质,可能对藻细胞的活性和后续利用造成不利影响,因此,对微藻污水处理系统可能并不是合适的选择. 基于磷酸钙沉淀的自絮凝无需任何额外投入,在微藻自然生长的pH范围内(8~10)就能形成,能同步实现化学除磷,对微藻活性和藻细胞的后续加工利用几乎没有不利影响. 因此,无论是从污水深度处理,还是从藻细胞的采收利用等角度来看都是最合适的分离采收方法之一. 后续研究应进一步明确其形成条件,并结合污水处理主体工艺的调整(取消强化除磷)、微藻培养方式的改进(高密度间歇培养、取消CO2供给,调节反应周期)等对其进行强化. 此外,还应重点关注微藻培养系统长期处于高pH条件下可能发生的种群结构和生理特性变化.
| [1] | Abdel-Raouf N, Al-Homaidan A A, Ibraheem I B M. 2012. Microalgae and wastewater treatment[J]. Saudi Journal of Biological Sciences, 19(3): 257-275 |
| [2] | Ahmad A L, Yasin N H M, Derek C J C, et al. 2011. Optimization of microalgae coagulation process using chitosan[J]. Chemical Engineering Journal, 173(3): 879-882 |
| [3] | Anthony R, Sims R. 2013. Cationic starch for microalgae and total phosphorus removal from wastewater[J]. Journal of Applied Polymer Science, 130(4): 2572-2578 |
| [4] | Banerjee C, Ghosh S, Sen G, et al. 2013. Study of algal biomass harvesting using cationic guar gum from the natural plant source as flocculant[J]. Carbohydrate Polymers, 92(1): 675-681 |
| [5] | Besson A, Guiraud P. 2013. High-pH-induced flocculation-flotation of the hypersaline microalga Dunaliella salina[J]. Bioresource Technology, 147: 464-470 |
| [6] | Beuckels A, Depraetere O, Vandamme D, et al. 2013. Influence of organic matter on flocculation of Chlorella vulgaris by calcium phosphate precipitation[J]. Biomass & Bioenergy, 54: 107-114 |
| [7] | Bilanovic D, Shelef G, Sukenik A. 1988. Flocculation of microalgae with cationic polymers-effect of medium salinity[J]. Biomass, 17(1): 65-76 |
| [8] | Blanchemain A, Grizeau D. 1999. Increased production of eicosapentaenoic acid by Skeletonema costatum cells after decantation at low temperature[J]. Biotechnology Techniques, 13(7): 497-501 |
| [9] | Borowitzka M, Borowitzka L. 1988. Micro-algal biotechnology[M]. UK: Cambridge University Press |
| [10] | Bos R, van der Mei H C, Busscher H J. 1999. Physico-chemistry of initial microbial adhesive interactions-its mechanisms and methods for study[J]. FEMS Microbiology Reviews, 23(2): 179-230 |
| [11] | 蔡卓平, 段舜山, 朱红惠. 2012. "污水-微藻-能源"串联技术新进展[J]. 生态环境学报, 21(7): 1380-1386 |
| [12] | Cai T, Park S Y, Li Y. 2013. Nutrient recovery from wastewater streams by microalgae: Status and prospects[J]. Renewable and Sustainable Energy Reviews, 19: 360-369 |
| [13] | Castrillo M, Lucas-Salas L M, Rodriguez-Gil C, et al. 2013. High pH-induced flocculation-sedimentation and effect of supernatant reuse on growth rate and lipid productivity of Scenedesmus obliquus and Chlorella vulgaris[J]. Bioresource Technology, 128: 324-329 |
| [14] | Chen C Y, Yeh K L, Aisyah R, et al. 2011. Cultivation, photobioreactor design and harvesting of microalgae for biodiesel production: A critical review[J]. Bioresource Technology, 102(1): 71-81 |
| [15] | Chen L, Wang C, Wang W, et al. 2013. Optimal conditions of different flocculation methods for harvesting Scenedesmus sp cultivated in an open-pond system[J]. Bioresource Technology, 133: 9-15 |
| [16] | 陈宗琪, 王光信, 徐桂英. 2001. 胶体与界面化学[M]. 北京: 高等教育出版社. 190 |
| [17] | Christenson L, Sims R. 2011. Production and harvesting of microalgae for wastewater treatment, biofuels, and bioproducts[J]. Biotechnology Advances, 29(6): 686-702 |
| [18] | Cordoba-Castro N M, Montenegro-Jaramillo A M, Prieto R E, et al. 2012. Analysis of the effect of the interactions among three processing variables for the production of exopolysaccharides in the microalgae scenedesmus obliquus (UTEX 393)[J]. Vitae-Revista De La Facultad De Quimica Farmaceutica, 19(1): 60-69 |
| [19] | De Philippis R, Sili C, Paperi R, et al. 2001. Exopolysaccharide-producing cyanobacteria and their possible exploitation: A review[J]. Journal of Applied Phycology, 13(4): 293-299 |
| [20] | Dumas A, Laliberte G, Lessard P, et al. 1998. Biotreatment of fish farm effluents using the cyanobacterium Phormidium bohneri[J]. Aquacultural Engineering, 17(1): 57-68 |
| [21] | Elmaleh S, Coma J, Grasmick A, et al. 1991. Magnesium induced algal flocculation in fluidized-bed[J]. Water Science and Technology, 23(7/9): 1695-1702 |
| [22] | Garzon-Sanabria A J, Ramirez-Caballero S S, Moss F E P, et al. 2013. Effect of algogenic organic matter (AOM) and sodium chloride on Nannochloropsis salina flocculation efficiency[J]. Bioresource Technology, 143: 231-237 |
| [23] | Golueke C, Oswald W J. 1965. Harvesting and processing sewage-grown planktonic algae[J]. Journal Water Pollution Control Federation, 37(4): 471-498 |
| [24] | Gonzalez-Fernandez C, Ballesteros M. 2013. Microalgae autoflocculation: an alternative to high-energy consuming harvesting methods[J]. Journal of Applied Phycology, 25(4): 991-999 |
| [25] | Guo S L, Zhao X Q, Wan C, et al. 2013. Characterization of flocculating agent from the self-flocculating microalga Scenedesmus obliquus AS-6-1 for efficient biomass harvest[J]. Bioresource Technology, 145: 285-289 |
| [26] | 胡洪营, 李鑫, 杨佳. 2009. 基于微藻细胞培养的水质深度净化与高价值生物质生产耦合技术[J]. 生态环境学报, 18(3): 1122-1127 |
| [27] | 胡洪营, 李鑫. 2010. 利用污水资源生产微藻生物柴油的关键技术及潜力分析[J]. 生态环境学报, 19(3): 739-744 |
| [28] | Jiang J Q, Graham N J D, Harward C. 1993. Comparison of polyferric sulphate with other coagulants for the removal of algae and algae-derived organic matter[J]. Water Science & Technology, 27(11): 221-230 |
| [29] | Jorand F, Boue-Bigne F, Block J C, et al. 1998. Hydrophobic/hydrophilic properties of activated sludge exopolymeric substances[J]. Water Science and Technology, 37(4/5): 307-315 |
| [30] | Kim D G, La H J, Ahn C Y, et al. 2011. Harvest of Scenedesmus sp with bioflocculant and reuse of culture medium for subsequent high-density cultures[J]. Bioresource Technology, 102(3): 3163-3168 |
| [31] | Lam M K, Lee K T. 2012. Microalgae biofuels: A critical review of issues, problems and the way forward[J]. Biotechnology Advances, 30(3): 673-690 |
| [32] | Lavoie A, Delanoue J. 1987. Harvesting of Scenedesmus obliquus in wastewaters: Auto-or bioflocculation?[J]. Biotechnology and Bioengineering, 30(7): 852-859 |
| [33] | Lavoie A, Delanoue J, Serodes J B. 1984. Harvesting of microalgae from waste-water-comparative-study of different flocculation agents[J]. Canadian Journal of Civil Engineering, 11(2): 266-272 |
| [34] | Lee A K, Lewis D M, Ashman P J. 2009. Microbial flocculation, a potentially low-cost harvesting technique for marine microalgae for the production of biodiesel[J]. Journal of Applied Phycology, 21(5): 559-567 |
| [35] | 雷国元, 张晓晴, 王丹鸶. 2007. 聚合铝盐混凝剂混凝除藻机理与强化除藻措施[J]. 水资源保护, 23(5): 50-54 |
| [36] | 李若慧, 叶晓, 程艳玲. 2012. 壳聚糖絮凝微藻富集的研究进展[J]. 安徽农业科学, 40(3): 1626-1628 |
| [37] | Liao B Q, Allen D G, Droppo I G, et al. 2001. Surface properties of sludge and their role in bioflocculation and settleability[J]. Water Research, 35(2): 339-350 |
| [38] | 刘晓猛. 2008. 微生物聚集体的相互作用及形成机制 [D]. 合肥: 中国科学技术大学. 12-17 |
| [39] | Liu Y, Fang H H P. 2003. Influences of extracellular polymeric substances (EPS) on flocculation, settling, and dewatering of activated sludge[J]. Critical Reviews in Environmental Science and Technology, 33(3): 237-273 |
| [40] | Lupi F M, Fernandes H M L, Sacorreia I, et al. 1991. Temperature profiles of cellular growth and exopolysaccharide synthesis by botryococus-braunii kutz uc-58[J]. Journal of Applied Phycology, 3(1): 35-42 |
| [41] | Milledge J J, Heaven S. 2013. A review of the harvesting of micro-algae for biofuel production[J]. Reviews in Environmental Science and Bio-Technology, 12(2): 165-178 |
| [42] | Molina Grima E, Belarbi E H, Acién Fernández F G, et al. 2003. Recovery of microalgal biomass and metabolites: process options and economics[J]. Biotechnology Advances, 20(7/8): 491-515 |
| [43] | Moreno J, Vargas M A, Olivares H, et al. 1998. Exopolysaccharide production by the cyanobacterium Anabaena sp. ATCC 33047 in batch and continuous culture[J]. Journal of Biotechnology, 60(3): 175-182 |
| [44] | Moutin T, Gal J Y, Elhalouani H, et al. 1992. Decrease of phosphate concentration in a high-rate pond by precipitation of calcium-phosphate-theoretical and experimental results[J]. Water Research, 26(11): 1445-1450 |
| [45] | Muñoz R, Guieysse B. 2006. Algal-bacterial processes for the treatment of hazardous contaminants: A review[J]. Water Research, 40(15): 2799-2815 |
| [46] | Oh H M, Lee S J, Park M H, et al. 2001. Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp AM49[J]. Biotechnology Letters, 23(15): 1229-1234 |
| [47] | Olguín E J. 2012. Dual purpose microalgae-bacteria-based systems that treat wastewater and produce biodiesel and chemical products within a Biorefinery[J]. Biotechnology Advances, 30(5): 1031-1046 |
| [48] | Olguín E J. 2003. Phycoremediation: key issues for cost-effective nutrient removal processes[J]. Biotechnology Advances, 22(1/2): 81-91 |
| [49] | Oswald W J, Gotaas H B, Golueke C G, et al. 1957. Algae in waste treatment[J]. Sewage and Industrial Wastes, 29(4): 437-455 |
| [50] | Ozkan A, Berberoglu H. 2013a. Adhesion of algal cells to surfaces [J]. Biofouling, 29(4): 469-482 |
| [51] | Ozkan A, Berberoglu H. 2013b. Physico-chemical surface properties of microalgae[J]. Colloids and surfaces (B: Biointerfaces), 112: 287-293 |
| [52] | Ozkan A, Berberoglu H. 2013c. Cell to substratum and cell to cell interactions of microalgae[J]. Colloids and Surfaces (B: Biointerfaces), 112: 302-309 |
| [53] | Papazi A, Makridis P, Divanach P. 2010. Harvesting Chlorella minutissima using cell coagulants[J]. Journal of Applied Phycology, 22(3): 349-355 |
| [54] | Park J B K, Craggs R J, Shilton A N. 2011. Wastewater treatment high rate algal ponds for biofuel production[J]. Bioresource Technology, 102(1): 35-42 |
| [55] | Pittman J K, Dean A P, Osundeko O. 2011. The potential of sustainable algal biofuel production using wastewater resources[J]. Bioresource Technology, 102(1): 17-25 |
| [56] | Powell R J, Hill R T. 2013. Rapid Aggregation of Biofuel-Producing Algae by the Bacterium Bacillus sp Strain RP1137[J]. Applied and Environmental Microbiology, 79(19): 6093-6101 |
| [57] | Pragya N, Pandey K K, Sahoo P K. 2013. A review on harvesting, oil extraction and biofuels production technologies from microalgae[J]. Renewable & Sustainable Energy Reviews, 24: 159-171 |
| [58] | 秦丽娟, 陈夫山. 2004. 有机高分子絮凝剂的研究进展及发展趋势[J]. 上海造纸, 35(1): 41-43, 46 |
| [59] | Rawat I, Ranjith Kumar R, Mutanda T, et al. 2011. Dual role of microalgae: Phycoremediation of domestic wastewater and biomass production for sustainable biofuels production[J]. Applied Energy, 88(10): 3411-3424 |
| [60] | Rebolloso Fuentes M M, Garcia Sanchez J L, Fernandez Sevilla J M, et al. 1999. Outdoor continuous culture of Porphyridium cruentum in a tubular photobioreactor: Quantitative analysis of the daily cyclic variation of culture parameters[J]. Journal of Biotechnology, 70(1/3): 271-288 |
| [61] | Salim S, Bosma R, Vermue M H, et al. 2011. Harvesting of microalgae by bio-flocculation[J]. Journal of Applied Phycology, 23(5): 849-855 |
| [62] | Salim S, Shi Z, Vermue M H, et al. 2013. Effect of growth phase on harvesting characteristics, autoflocculation and lipid content of Ettlia texensis for microalgal biodiesel production[J]. Bioresource Technology, 138: 214-221 |
| [63] | Sanyano N, Chetpattananondh P, Chongkhong S. 2013. Coagulation-flocculation of marine Chlorella sp. for biodiesel production[J]. Bioresource Technology, 147: 471-476 |
| [64] | Schlesinger A, Eisenstadt D, Bar-Gil A, et al. 2012. Inexpensive non-toxic flocculation of microalgae contradicts theories; overcoming a major hurdle to bulk algal production[J]. Biotechnology Advances, 30(5): 1023-1030 |
| [65] | Sérodes J B, Walsh E, Goulet O, et al. 1991. Tertiary treatment of municipal wastewater using bioflocculating micro-algae[J]. Canadian Journal of Civil Engineering, 18(6): 940-944 |
| [66] | Sirin S, Trobajo R, Ibanez C, et al. 2012. Harvesting the microalgae Phaeodactylum tricornutum with polyaluminum chloride, aluminium sulphate, chitosan and alkalinity-induced flocculation[J]. Journal of Applied Phycology, 24(5): 1067-1080 |
| [67] | Smith B T, Davis R H. 2012. Sedimentation of algae flocculated using naturally-available, magnesium-based flocculants[J]. Algal Research, 1(1): 32-39 |
| [68] | Sobeck D C, Higgins M J. 2002. Examination of three theories for mechanisms of cation-induced bioflocculation[J]. Water Research, 36(3): 527-538 |
| [69] | Spilling K, Seppälä J, Tamminen T. 2011. Inducing autoflocculation in the diatom Phaeodactylum tricornutum through CO2 regulation[J]. Journal of Applied Phycology, 23(6): 959-966 |
| [70] | Sukenik A, Shelef G. 1984. Algal autoflocculation—verification and proposed mechanism[J]. Biotechnology and Bioengineering, 26(2): 142-147 |
| [71] | Surendhiran D, Vijay M. 2013. Influence of bioflocculation parameters on harvesting Chlorella salina and its optimization using response surface methodology[J]. Journal of Environmental Chemical Engineering, 1(4): 1051-1056 |
| [72] | Talbot P, Delanoue J. 1993. Tertiary-treatment of waste-water with phormidium-bohneri (schmidle) under various light and temperature conditions[J]. Water Research, 27(1): 153-159 |
| [73] | U.S.DOE. 2010. National algal biofuels technology roadmap[D]. U.S. Department of Energy, Office of Energy Efficiency and Renewable Energy, Biomass Program |
| [74] | Vandamme D, Foubert I, Fraeye I, et al. 2012a. Flocculation of Chlorella vulgaris induced by high pH: Role of magnesium and calcium and practical implications[J]. Bioresource Technology, 105: 114-119 |
| [75] | Vandamme D, Foubert I, Fraeye I, et al. 2012b. Influence of organic matter generated by Chlorella vulgaris on five different modes of flocculation[J]. Bioresource Technology, 124: 508-511 |
| [76] | Vandamme D, Foubert I, Meesschaert B, et al. 2010. Flocculation of microalgae using cationic starch[J]. Journal of Applied Phycology, 22(4): 525-530 |
| [77] | Vandamme D, Foubert I, Muylaert K. 2013. Flocculation as a low-cost method for harvesting microalgae for bulk biomass production[J]. Trends in Biotechnology, 31(4): 233-239 |
| [78] | Wan C, Zhao X Q, Guo S L, et al. 2013. Bioflocculant production from Solibacillus silvestris W01 and its application in cost-effective harvest of marine microalga Nannochloropsis oceanica by flocculation[J]. Bioresource Technology, 135: 207-212 |
| [79] | 王九思, 陈学民, 肖举强, 等. 2002. 水处理化学[M]. 北京: 化学工业出版社 |
| [80] | Wu Z, Zhu Y, Huang W, et al. 2012. Evaluation of flocculation induced by pH increase for harvesting microalgae and reuse of flocculated medium[J]. Bioresource Technology, 110: 496-502 |
| [81] | Wyatt N B, Gloe L M, Brady P V, et al. 2012. Critical conditions for ferric chloride-induced flocculation of freshwater algae[J]. Biotechnology and Bioengineering, 109(2): 493-501 |
| [82] | 邢丽贞, 马清, 李哿, 等. 2009. 藻类技术在污水深度处理中的应用[J]. 净水技术, 28(6): 44-49 |
| [83] | 薛蓉, 陆向红, 卢美贞, 等. 2012. 絮凝法采收小球藻的研究[J]. 可再生能源, 30(9): 80-84 |
| [84] | Yahi H, Elmaleh S, Coma J. 1994. Algal flocculation-sedimentation by pH increase in a continuous reactor[J]. Water Science and Technology, 30(8): 259-267 |
| [85] | 严煦世, 范瑾初. 1999. 给水工程[M]. 北京: 中国建筑工业出版社 |
| [86] | 翟玥, 杨哲, 安阳, 等. 2009. 壳聚糖凝聚去除景观水中微囊藻的研究[J]. 净水技术, 28(6): 58-60, 68 |
| [87] | Zhang X, Amendola P, Hewson J C, et al. 2012. Influence of growth phase on harvesting of Chlorella zofingiensis by dissolved air flotation[J]. Bioresource Technology, 116: 477-484 |
| [88] | 张亚杰, 罗生军, 蒋礼玲, 等. 2010. 阳离子絮凝剂对小球藻浓缩收集效果的研究[J]. 可再生能源, 28(3): 35-38 |
| [89] | 曾文炉, 李宝华, 蔡昭铃, 等. 2003. 微藻细胞的连续气浮法采收[J]. 水生生物学报, 27(5): 507-511 |
| [90] | Zheng H, Gao Z, Yin J, et al. 2012. Harvesting of microalgae by flocculation with poly (gamma-glutamic acid)[J]. Bioresource Technology, 112: 212-220 |