 2014, Vol. 34
2014, Vol. 34



2. 白银市环保局, 白银 730900
2. Baiyin Environmental Protection Bureau, Baiyin 730900
Ni是植物生长必需的微量元素(龙新宪和杨肖娥,2000),适量的Ni能够促进植物生长,但是过量的Ni抑制植物的生长并产生毒害作用(张秀玲和宋光煜,2004;Molas,1997).Ni能在农作物中累积,并通过食物链对人体产生危害,比如导致心肌、脑、肺、肝和肾退行性变(唐荣莉等,2012).Ni矿是我国甘肃省的主要有色金属矿产.Ni矿的开采冶炼加工等工业的兴起和发展,极大地推动了当地经济发展和社会进步,但是也带来了严重的环境污染问题(南忠仁等,2011).其中,土壤Ni污染问题最为普遍.有研究显示,河西走廊绿洲农田土壤中Ni含量平均为41.39 mg · kg-1(赵翠翠等,2010),高于甘肃省土壤背景值35.2 mg · kg-1(中国环境监测总站,1990).干旱区绿洲土壤中Ni含量不仅总量较高,而且Ni的各种赋存形态的含量也与其他土壤有很大不同,这导致了绿洲土-植物系统中Ni的生物有效性也发生了变化.土壤-植物系统中重金属的迁移特性和生物有效性主要与重金属在土壤中的赋存形态有关,不同形态的重金属往往具有不同的生物可利用性,一般认为可交换态是生物有效态(杨军等,2005).但是干旱区绿洲土壤中碳酸盐含量较高,碳酸盐结合态重金属也会被植物大量吸收利用(赵转军等,2010).因此,对于河西走廊绿洲区Ni在土壤-植物系统中的迁移及其生物效应的研究对污染土壤的修复和人体健康风险评价均具有重要意义.
目前,关于Ni在土壤-植物系统中的迁移和积累的研究较多,如Ni导致豌豆的根瘤菌数量明显减少且丧失本来的能力(Simon,2000),过量的Ni可降低小麦叶肉的厚度(Kovačević et al., 1999),张璇等(2008)研究了不同pH条件下Ni对大麦的急性毒性,王晓瑞等(2011)研究了长江三角洲地区小麦的重金属分布情况,罗丹等(2010)研究了Ni对多种蔬菜的毒害临界值等;关于Ni对小麦的作用研究也多集中于Ni对小麦幼苗的生长影响上(王启明和王素平,2010;丁竹红等,2010).对西北绿洲区同类研究则较少.本研究以西北干旱区绿洲灌淤土为供试土壤,采用盆栽模拟试验研究Ni对小麦生长的影响及土壤中Ni向小麦的富集迁移作用,并分析了小麦中Ni含量与土壤中Ni的各种形态的相关关系,以期为干旱区绿洲土壤的重金属污染管理提供一定的科学依据.
2 材料与方法(Materials and methods) 2.1 供试土壤与作物供试作物为小麦,种子购自张掖市种子公司.供试土壤为甘肃省河西走廊地区的绿洲灌淤土,属农田耕作土.供试土壤的基本理化性质为:pH 8.16,有机质含量15.49 g · kg-1,阳离子交换量8.1 cmol · kg-1,碳酸盐含量56.4 g · kg-1,全N 0.07%,全P 0.061%,全K 2.32%.土壤中Ni的背景值36.9 mg · kg-1,达到土壤环境质量标准(国家环境保护局,1995)中适用于一般农田的Ⅱ类标准(Ni含量60 mg · kg-1).供试土壤自然风干后压碎过2 mm尼龙筛,称取若干份4.0 kg土壤待用.
2.2 试验设计根据长期对研究区域土壤Ni污染调查中发现的最大浓度(406.58 mg · kg-1),并且考虑到污染还有继续增加的可能,研究中对Ni浓度的上限做适当的延伸,同时结合土壤环境质量标准(国家环境保护局,1995)中关于Ni的分级标准进行梯度划分,确定采用Ni的7种添加质量分数(表 1),其中CK为对照组,T1~T6为不同添加质量分数组.每个添加水平设3个平行样.
| 表1 试验土壤中Ni的质量分数添加设计 Table.1 Concentrations of Ni added into soil samples |
每个花盆加入4.0 kg供试土壤,Ni以硝酸盐的形式加入土壤,平衡30 d.每盆加入6.00 g羊粪(羊粪中Ni含量1.54 mg · kg-1,有机质含量25%,全N 0.7%,全P 0.48%,全K 0.43%)作为基肥,然后加入蒸馏水使其含水量为田间持水量的60%,保持2 d.每盆播种小麦种子30粒,于三叶期定苗15株.小麦生长期每3 d浇水1次,以保持其含水量为田间持水量的60%.每盆于分蘖期和拔节期分别追施3.0 g专用肥(N-P2O5-K2O含量14-9-13),生长管理同大田,生长约120 d后收获.收获后用去离子水洗净其根、茎叶和穗,烘干至恒重后分别称量根、茎叶、穗和籽粒的重量,再分别粉碎后测定根、茎叶、颖壳和籽粒中的Ni含量,同时采用四分法取收获后土样100 g左右,经自然风干、磨细后过100目尼龙筛,装袋备用.
2.3 样品分析土壤理化性质测试与分析采用土壤农业化学常规分析方法(鲁如坤,1999).小麦样品采用GB/T 5009中规定的HNO3-HClO4体系进行消解.土样采用HNO3-HF-HClO4三酸法消解(中国环境检测总站,2004),利用Tessier五步连续提取法开展形态分析(Tessier et al., 1979),分别提取土壤重金属的可交换态(Exchangeable form)、碳酸盐结合态(Carbonate bounded form)、铁锰氧化态(Fe-Mn oxides bounded form)、有机态(Organic matter bounded form)和残渣态(Residual form).所有样品采用美国Thermo Fisher公司的SOLLAR AAM6原子吸收光谱仪测定Ni含量(中国环境监测总站,1997).
试验中采用20%平行样和GSB-6标准菠菜样及GSS-1标准土样进行质量监控,误差控制在5%之内.试验试剂均选用优级纯试剂,试验器皿在使用前均经过10%硝酸浸泡24 h以上.
2.4 数据分析为研究土壤中Ni的各形态组成,利用下式计算土壤中Ni的形态比例系数P:

为研究小麦中Ni的富集迁移特征,利用下式计算Ni的生物富集系数(Bioconcentration factor)(方如康,2003)和迁移系数(Translocation factor)(李晓晨等,2007).生物富集系数是衡量土壤重金属被植物吸收难易程度的指标,其值越大表示该重金属越易于植物吸收.迁移系数是反应重金属在植物体内迁移能力的指标,其值越大代表该重金属由植物地下部迁移到地上部的能力越强.
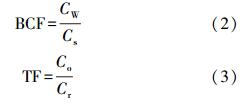
差异性显著检验(p<0.05)表明,外源Ni各添加质量分数下各器官的干重均与对照组(CK)达到显著性差异,且各质量分数之间达到显著性差异(图 1).这说明外源Ni对小麦生长产生了较大影响.
随着外源Ni添加质量分数的增加,小麦各器官的干重呈现出相似的变化趋势. Ni质量分数低于240 mg · kg-1(T4)时,各器官干重随着Ni添加质量分数的增加而增加;但高于 240 mg · kg-1(T4)后,各器官干重随着Ni添加质量分数的增加而减小. 这说明,在添加质量分数240 mg · kg-1前后,Ni对小麦的生长分别起到促进和抑制的作用.Ni添加质量分数为240 mg · kg-1时,茎叶、根、穗和籽粒的干重分别比对照组增加了21.9%、59.3%、34.1%和24.1%.Ni添加质量分数从240 mg · kg-1增加至360 mg · kg-1(T5)时,茎叶、根、穗和籽粒的干重分别减少12.2%、15.2%、12.7%和6.2%. Ni质量分数从360 mg · kg-1增加至600 mg · kg-1(T6)时,茎叶、根、穗和籽粒的干重分别减少73.5%、79.9%、62.6%和49.6%.随着Ni添加质量分数的增加,根干重的增加和减小的比例最大,籽粒干重的增加和减小比例最小.这说明,外源Ni的添加对根生长的影响大于对籽粒生长的影响.
 |
| 图 1 不同Ni添加质量分数下小麦各器官干重 Fig. 1 Dry weight of different organs under different treatments of Ni |
差异性显著检验(p<0.05)表明,各添加质量分数下各器官中Ni含量均与对照组(CK)达到显著性差异.添加质量分数越大,差异也越大.且各质量分数之间达到显著性差异(图 2).每个添加质量分数的3个平行样的各器官对Ni的生物富集系数和迁移系数进行差异显著性分析发现,各平行样之间差异性不显著;而不同添加质量分数下同一器官对Ni的生物富集系数和迁移系数差异显著(表 2).
 |
| 图 2 不同Ni添加质量分数下小麦各器官中Ni含量的变化 Fig. 2 Concentrations of Ni in different organs under different treatments |
| 表2 不同Ni添加质量分数下小麦各器官中Ni的生物富集系数和迁移系数 Table.2 BCF and TC of Ni in wheat under different treatments |
随着Ni添加质量分数的增加,各器官中Ni含量也呈增长趋势.各器官中Ni含量的大小关系为:根>茎叶>颖壳>籽粒.根中Ni含量平均为茎叶、颖壳和籽粒的349%、444%和645%.各添加质量分数之间,茎叶、根、颖壳和籽粒中Ni含量平均增长了49.1%、74.4%、67.6%和62.9%.根中Ni含量增长最快,茎叶中Ni含量增长最慢.尽管籽粒中Ni含量增长较快,但是籽粒中Ni含量是最低的,因此根是Ni的主要贮存器官.各器官对Ni的生物富集系数的大小关系为:根>颖壳>茎叶>籽粒,根的生物富集系数最大,籽粒的富集系数最小.无论Ni含量的变化还是生物富集系数的变化都说明,小麦从土壤中吸收的Ni大部分累积在根部,很小部分在籽粒累积.随着Ni添加质量分数的增加,各器官Ni的生物富集系数和迁移系数在Ni添加质量分数240 mg · kg-1(T4)之前呈增加趋势,这之后呈减少趋势.Ni在小麦中的迁移系数均小于1,说明Ni在小麦中的迁移能力较弱,小麦从土壤中吸收的Ni主要在根部累积.
3.3 小麦中Ni含量与土壤中Ni形态的关系差异性显著检验(p<0.05)表明,外源Ni各添加质量分数下土壤中Ni的各形态比例系数与对照组(CK)的达到显著性差异,且各质量分数之间达到显著性差异(图 3).这说明,外源Ni对土壤中Ni的存在形态产生了较大影响.
 |
| 图 3 不同Ni添加质量分数下土壤中Ni的形态比例系数 Fig. 3 Percentages of different forms of Ni under different treatments |
土壤中Ni的同一形态比例系数在Ni不同的添加质量分数下存在很大差异(图 3).在对照组(CK)中,土壤中Ni的主要赋存形态为残渣态.随着Ni质量分数的增加,可交换态Ni和碳酸盐结合态Ni所占比例逐渐增加,残渣态Ni所占比例逐渐减小;并且,碳酸盐结合态逐渐成为Ni的主要赋存形态.这说明,随着外源Ni添加量的增加,土壤中Ni的各存在形态比例系数均发生了显著变化.
对小麦各器官中Ni含量与各形态比例系数做相关分析(表 3)发现,各器官中Ni含量与残渣态Ni比例系数和有机结合态Ni比例系数呈负相关,与可交换态Ni比例系数、碳酸盐结合态Ni比例系数和铁锰氧化态Ni比例系数呈正相关,其中与可交换态Ni比例系数呈极显著正相关关系.这说明,可交换态Ni是影响小麦中Ni含量的主要因素,更容易被小麦吸收,而残渣态Ni和有机结合态Ni则不会被小麦吸收.对各器官中Ni含量与各形态比例系数进行逐步回归分析(表 4)发现,各器官中Ni含量与可交换态Ni比例系数之间存在很强的线性关系.逐步回归分析保留了可交换态Ni对小麦中Ni含量的影响,进一步说明小麦主要从土壤中吸收可交换态Ni.
| 表3 各器官中Ni含量与土壤中Ni各形态比例系数的相关分析 Table.3 Correlation between the concentrations of Ni in wheat and percentages of different forms in soil |
| 表4 各器官中Ni含量与土壤中Ni各形态比例系数的逐步回归分析 Table.4 Stepwise regression analysis between the concentrations of Ni in wheat and percentages of different forms in soil |
Ni是一些高等植物的必须营养元素(Eskew et al., 1983),也是一种极毒元素.有研究表明,低浓度的Ni能够促进小麦幼苗的生长,而高浓度的Ni起抑制作用,且对根的抑制作用大于其他器官(杨红超和王素平,2011).Ni对其他作物的生长也表现出低浓度刺激和高浓度阻碍的作用(杨定清等,1994).本研究也得出类似结果.Ni是植物生长的必需元素,是一些植物脲酶和一氧化碳脱氢酶、甲醛还原酶和氢酶的金属辅基(扶惠华等,1996;罗红艺和景红娟,2002).适量的Ni能够促进植物中这些酶的合成,有利于干物质的积累,促进植物生长.但是过量的Ni会取代叶绿素中心离子Mg,引起叶绿素含量降低,导致光合作用崩溃(Kupper et al., 1998;Ewais,1997).另外,过量的Ni也会破坏植物的膜结构,引起生理生化代谢紊乱,最终导致植物枯萎死亡(王光忠等,2003).目前,关于谷物中Ni的食品卫生标准尚未制定,一般以作物减产10%时的土壤总Ni含量作为Ni的临界值.本研究发现,Ni添加质量分数从240 mg · kg-1增加至360 mg · kg-1时,各器官干重平均减少11.6%,可以确定在本研究中Ni的临界值约为Ni添加质量分数240 mg · kg-1.张素霞和王宏康(1991)对小麦的盆栽实验得出小麦受害的Ni的投加量为150 mg · kg-1.土壤环境质量标准中保障农林业生产和植物正常生长的土壤Ni的临界值为200 mg · kg-1,而在本研究中Ni的添加质量分数为180 mg · kg-1时,土壤中Ni含量达到216.9 mg · kg-1,超过土壤环境质量标准中Ni临界值.土壤环境质量标准中Ni临界值是结合多数作物及不同土壤类型得出,本研究结果与之接近.不同种类的作物对Ni的耐受能力不尽相同,另外,干旱区绿洲土壤较高的pH和碳酸盐含量也可能是导致临界值出现差异的原因.因此,临界值的差异可能是由于选用的作物品种不同及供试土壤性质不同.
4.2 小麦中Ni的富集与迁移重金属在谷类植物体内分布的一般规律为根>茎叶>颖壳>籽实(戴树桂,2006),本研究结果与之相符.无论是Ni含量还是生物富集系数,都是根部的高于其他器官的,籽粒的小于其他器官的.李媛等(2008)对金昌市市郊农田-小麦系统中Ni行为特性研究也发现,在各器官中,根中Ni含量最高,籽粒中Ni含量最低.另外,Ni在小麦中的迁移系数均小于1,说明Ni在小麦中不易迁移.因此,小麦从土壤中吸收的Ni大部分在根累积,极少部分积累在籽粒中,且Ni在小麦中迁移能力很弱.但是,这并不意味着籽粒中的Ni对人体健康就没有危害.虽然关于Ni在谷物中的食品卫生标准尚未制定,但是参照油脂及其制品(中华人民共和国卫生部,2012)中Ni的限量1 mg · kg-1,仅对照组小麦籽粒中Ni含量(4.71 mg · kg-1)就远远超标.但是,小麦干重开始减少的临界值为240 mg · kg-1(T4),说明在研究区土壤中种植的小麦能够耐受较高浓度的Ni污染.同时小麦中Ni含量处于可能对人体健康产生危害的水平.因此,仅从Ni污染来看,研究区土壤不适宜种植小麦.
各器官的生物富集系数和迁移系数在临界值时最大,在这前后分别呈增长和减小趋势.这可能是由于临界值前后Ni对小麦的生长分别起到促进和抑制的作用.在临界值以前,Ni能够刺激小麦生长,一方面小麦大量吸收Ni合成各种酶,另一方面小麦干重的增加也能容纳更多的Ni.在临界值之后,小麦干重迅速减小,生理生化代谢紊乱,出现植株矮小、叶片发黄萎蔫等症状,导致小麦中Ni含量虽然增加但是富集能力减小.
4.3 Ni的生物有效性土壤-植物系统中重金属的积累能力和生物毒性,不但和其总量有关,很大程度上也是由其形态分布决定的,不同形态产生不同的环境效果(Ramos et al., 1994).土壤重金属之所以表现出不同的生物活性,是由于土壤中不同形态的重金属处于不同的能量状态,其在土壤中的迁移性不同(Xian et al., 1989).一般认为,可交换态和水溶态重金属是植物可吸收利用的主要形态(Wang et al., 1997).本研究发现可交换态Ni更易于被小麦吸收,是生物有效态,结果与之类似.对土壤中Ni的各形态比例系数分析发现,可交换态Ni和碳酸盐结合态Ni所占比例逐渐增加.虽然各器官中Ni含量与碳酸盐结合态Ni的相关性不高,但是在干旱区土壤中碳酸盐结合态重金属是生物的潜可利用态.赵转军等(2010)研究发现,在干旱区绿洲土壤中碳酸盐含量较高,因而碳酸盐结合态重金属也有较高的生物有效性.随着Ni添加质量分数的增大,土壤中碳酸盐结合态Ni的比例系数呈增大趋势.随着土壤中可交换态Ni被小麦逐渐吸收,碳酸盐结合态Ni就由生物潜可利用态变为生物可利用态.因此,外源Ni污染对研究区小麦的危害是持久的.
5 结论(Conclusions)1)外源Ni在低添加质量分数下能促进小麦生长,但达到临界值240 mg · kg-1后,就会出现明显的毒害作用.其中,外源Ni对根生长的影响大于对籽粒的.
2)随着Ni添加质量分数的增加,各器官中Ni含量也呈增长趋势.其中根是Ni的主要贮存器官,而且根对Ni的富集能力最强,籽粒对Ni的富集能力最弱.总体来说,Ni在小麦中迁移能力较弱.
3)随着Ni添加质量分数的增加,土壤中Ni的存在形态发生了较大变化.可交换态Ni和碳酸盐结合态Ni所占比例呈增加趋势,残渣态Ni所占比例逐渐减少.Ni的主要赋存形态由残渣态变为碳酸盐结合态.
4)土壤中可交换态Ni是影响小麦中Ni含量的主要因素,更容易被小麦吸收,是生物有效态.
| [1] | 戴树桂. 2006. 环境化学[M]. 北京: 高等教出版社 |
| [2] | 丁竹红, 胡忻, 张宇峰. 2010. 螯合剂对小麦幼苗吸收金属以及土壤金属形态的效应[J]. 生态环境学报, 19(1): 97-101 |
| [3] | Eskew D L, Welch R M, Cary E E. 1983. Nickel: an essential micronutrient for legumes and possibly all higher plants[J]. Science, 222(4642): 621-623 |
| [4] | Ewais E A. 1997. Effects of cadmium, nickel and lead on growth, chlorophyll content and proteins of weeds[J]. Biologia Plantarum, 39(3): 403-410 |
| [5] | 方如康. 2003. 环境学词典[M]. 北京: 科学出版社. 127-128 |
| [6] | 扶惠华, 王煜, 田廷亮. 1996. 镍在植物生命活动中的作用[J].植物生理学通讯, 32(1): 45-49 |
| [7] | 国家环境保护局. 1995. GB 15618-1995 土壤环境质量标准[S]. 北京: 中国标准出版社 |
| [8] | Kovačević G, Kastori R, Merkulov L J. 1999. Dry matter and leaf structure in young wheat plant as affected by cadmium, lead and nickel[J]. Biologia Plantarum, 42(1): 119-123 |
| [9] | Kupper H, Kupper F, Spiller M. 1998. In situ detection of heavy metal substituted chlorophylls in water plants[J]. Photosynthesis Research, 58(2): 123-133 |
| [10] | 李晓晨, 赵丽, 赵星明. 2007. 南京市郊区蔬菜重金属污染特征的研究[J]. 安徽农业科学, 35(30): 9650-9651 |
| [11] | 李媛, 南忠仁, 刘晓文, 等. 2008. 金昌市市郊农田土壤-小麦系统Cu、Zn、Ni行为特性[J]. 西北农业学报, 17(6): 298-302 |
| [12] | 龙新宪, 杨肖娥. 2000. 植物镍营养[J]. 土壤通报, 31(1): 39-42 |
| [13] | 鲁如坤. 1999. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社 |
| [14] | 罗丹, 胡欣欣, 郑海锋, 等. 2010. 镍对蔬菜毒害临界值的研究[J]. 生态环境学报, 19(3): 584-589 |
| [15] | 罗红艺, 景红娟. 2002. 植物营养中新的必需元素——钠、镍、硅[J]. 高等函授学报(自然科学版), 15(3): 42-45 |
| [16] | Molas J. 1998. Changes in morphological and anatomical structure of cabbage (Brassica oleracea L.) outer leaves and in ultrastructure of their chloroplasts caused by an in vitro excess of nickel[J]. Photosynthetica, 34(4): 513-522 |
| [17] | 南忠仁, 刘晓文, 赵转军, 等. 2011. 干旱区绿洲土壤作物系统重金属化学行为与生态风险评估研究[M]. 北京: 中国环境科学出版社. 7-27 |
| [18] | Ramos L, Hernandez L M, Gonzalez M J. 1994. Sequential fractionation of copper, lead, cadmium and zinc in soils from or near donana National park[J]. Journal of Environmental Quality, 23(1): 50-57 |
| [19] | Simon T. 2000. The effect of nickel and arsenic on the occurrence and symbiotic abilities of native rhizobia[J]. Rostlinná V ý roba, 46(2): 63-68 |
| [20] | 唐荣莉, 马克明, 张育新, 等. 2012. 北京城市道路灰尘重金属污染的健康风险评价[J]. 环境科学学报, 32(8): 2006-2015 |
| [21] | Tessier A, Campbell P G C, Bisson M. 1979. Sequential extraction procedure for the speciation of particulate trace metals[J]. Analytical Chemistry, 51(7): 844-851 |
| [22] | 王光忠, 张芬琴, 马爱山, 等. 2003. Ni2+对小麦幼苗生长的影响[J]. 河西学院学报, 19(5): 87-89 |
| [23] | Wang P X, Qu E F, Li Z B, et al. 1997. Fractions and availability of Nickel in loessial soil amended with sewage sludge[J]. Journal of Environmental Quality, 26(3): 795-801 |
| [24] | 王启明, 王素平. 2010. 镍对小麦幼苗POD活性和同工酶的影响[J].种子, 29(8): 42-44 |
| [25] | 王晓瑞, 周生路, 吴绍华. 2011. 长江三角洲地区小麦植株的重金属分布及其相关性——以昆山市为例[J]. 地理科学, 31(2): 226-231 |
| [26] | Xian X. 1989. Effect of chemical forms of cadmium, zinc and lead in polluted soils on their uptake by cabbage plants[J]. Plant and Soil, 113(2): 257-264 |
| [27] | 杨定清, 傅绍清, 青长乐. 1994. 镍的作物效应及临界值研究[J]. 四川环境, 13(1): 19-23 |
| [28] | 杨红超, 王素平. 2011. 重金属镍胁迫对小麦种子萌发及幼苗生长的影响[J]. 种子, 30(12): 18-20 |
| [29] | 杨军, 陈同斌, 郑袁明, 等. 2005. 北京市凉凤灌区小麦重金属含量的动态变化及健康风险分析[J]. 环境科学学报, 25(12): 1661-1668 |
| [30] | 张素霞, 王宏康. 1991. 污泥施肥时重金属镍对农作物的毒害研究[J]. 环境科学学报, 11(1): 71-78 |
| [31] | 张秀玲, 宋光煜. 2004. 植物的镍素营养[J]. 土壤肥料, (6): 33-36 |
| [32] | 张璇, 华珞, 王学东, 等. 2008. 不同pH值条件下镍对大麦的急性毒性[J]. 中国环境科学, 28(7): 640-645 |
| [33] | 赵翠翠, 南忠仁, 刘晓文, 等. 2010. 绿洲农田土壤主要微量元素的影响因素及分布特征研究——以张掖甘州区和临泽县为例[J]. 干旱区资源与环境, 24(10): 127-132 |
| [34] | 赵转军, 南忠仁, 王兆炜, 等. 2010. Cd、Zn复合污染菜地土壤中重金属形态分布与植物有效性[J]. 兰州大学学报(自然科学版), 46(2): 1-5 |
| [35] | 中国环境监测总站. 1990. 中国土壤环境背景值[M]. 北京: 中国环境科学出版社 |
| [36] | 中国环境监测总站. 1997. GB/T 17139-1997土壤质量镍的测定火焰原子吸收分光光度法[S]. 北京: 中国标准出版社 |
| [37] | 中国环境检测总站. 2004. HJ/T 166-2004土壤环境监测技术规范[S]. 北京: 中国环境科学出版社 |
| [38] | 中华人民共和国卫生部. 2012. GB 2762-2012 食品安全国家标准 食品中污染物限量[S]. 北京: 中国标准出版社 |