 2014, Vol. 34
2014, Vol. 34



浮游植物休眠体是沉积物中特殊的生物类群,其种类组成和分布特征与海洋环境的营养盐含量、水温及盐度等密切相关,同时沉积物的类型、颗粒大小和沉积速率及人为因素等也均能对其分布产生显著影响(Genovesi et al., 2011; Prebble et al., 2013; Hessler et al., 2013).因此,沉积物中的浮游植物休眠体是一种有效的海洋环境变化的生物指示指标和信号载体(Matsuoka,1999; Zonneveld et al., 2012; Liu et al., 2013).
甲藻孢囊是甲藻有性生殖的产物,具有较厚的细胞壁,能抵御酸碱腐蚀和缺氧、黑暗环境.研究发现,甲藻孢囊的分布能够较为全面地反映海域富营养化的演变历史(Matsuoka,1999;Zonneveld et al., 2012; Liu et al., 2013).甲藻孢囊萌发可为水体提供营养细胞,被认为是引发赤潮的“种源”(Kremp et al., 2000);而孢囊的形成又将大量营养细胞从水体转移至沉积物中,是赤潮后期营养细胞大量减少、赤潮消亡的重要原因(Kremp et al., 1999;Wang et al., 2007).
柘林湾地处闽粤两省交界,是南海西北部的一个半封闭型海湾,同时也是广东最大的海产养殖基地之一.养殖业的发展与陆源排污排废的影响,致使该海域富营养化问题日益突出,浮游植物与浮游动物的生物多样性下降,群落组成呈小型化的趋势,成为赤潮多发区(黄长江等,1999;周凯等,2002).为了了解甲藻孢囊在柘林湾的分布规律及其萌发趋势,本研究采集柘林湾10个站位的表层沉积物样品,对沉积物中甲藻孢囊进行定性定量分析,并对沉积物中浮游植物休眠体进行萌发实验,对萌发的浮游植物群落结构进行分析,以了解甲藻孢囊的分布状况,揭示柘林湾海域赤潮发生潜势.
2 材料与方法(Materials and methods) 2.1 采样点的设置与样品的采集在柘林湾海域设置了10个采样点(图 1),于2008年11月用彼得逊采泥器采集表层沉积物,样品采集后,用透明PVC管垂直插入采集的沉积物中,得到深度约10 cm的柱状样品,取上表层0~2 cm沉积物,置于低温(4 ℃)、缺氧(充入氮气)、避光条件下保存待分析.
 |
| 图 1 本次调查中采样点的设置 Fig. 1 Sampling stations in this study |
称取沉积物样品3 g左右,在70 ℃烘箱中烘干(24 h),以测定沉积物中的含水率.另称取沉积物样品3~5 g,用盐酸和氢氟酸分别去除沉积物中的钙质、硅质,然后分别用125 μm和20 μm网筛过滤,收集20~125 μm之间的悬浮液至小塑料瓶中,定容至10 mL.
取1 mL处理后的样品于计数框内,在Leica DMIRB倒置显微镜下观察,对甲藻孢囊进行定性定量分析,分别计数含有内含物的活体孢囊和已经萌发的空孢囊.重复取样观察,使每个样品至少观察到100~200个孢囊.表层沉积物中孢囊的密度以每克干沉积物中的孢囊数(cysts · g-1)表示.
孢囊分类鉴定至种,在不能确定种名的情况下,则鉴定至属.由于甲藻孢囊最初是孢粉学家研究的对象,早期孢囊分类学研究是根据孢囊的孢粉学特征进行分类命名,随着孢囊研究的深入,发现现存甲藻也能形成孢囊,因此,孢囊分类学具有两套分类系统.孢粉学学名能更直观反映孢囊的形态特征,而生物学学名则能反映孢囊与营养细胞的对应关系,目前在孢囊研究中两套分类系统并存,因此,本文中也同时列出了甲藻孢囊的孢粉学学名、营养细胞学名和中文名称.孢囊分类鉴定主要参考文献(Matuoka et al., 2000;王朝晖,2007).
2.3 浮游植物休眠体的萌发与观察由于浮游植物休眠体均具有一定时间的强制性休眠期,只有度过了强制性休眠期的休眠体方具有萌发潜势.强制性休眠期的长短与种类及保存的环境状况有关,一般为2周至半年不等(Matsuoka et al., 2000).沉积物样品在4 ℃下缺氧避光保存1年后再进行萌发实验,以保证全部休眠体度过强制性休眠期,同时也可使表层沉积物中浮游植物营养细胞失去生长活性,确保观察到的浮游植物全部为休眠体萌发而得.
称取1 g表层沉积物样品,加入到50 mL的三角瓶中,加入培养基40 mL.培养基用柘林湾海域陈旧的自然海水经GF/C滤膜过滤后配制而成,培养基中添加的氮、磷、硅浓度根据自然海水中营养盐水平配制,分别为500 μg · L-1氮、74 μg · L-1磷和1000 μg · L-1 硅,其余元素与1/2 f培养基相同.培养基和所有试验容器均经过高压蒸汽灭菌(121 ℃、15 min)后备用.萌发温度为25 ℃,在光照强度为100 μmol · m-2 · s-1、光暗比12 ∶ 12 h(L ∶ D)的恒温培养箱中进行萌发,每天摇动1次.每个沉积物样品设20 d和40 d萌发2组实验,每个实验组各设3个平行.在培养的第20 d和40 d,各取上清30 mL,合并3个平行样品,加入4%的甲醛溶液固定,静置24 h以上,逐步浓缩成2 mL待观察.
在Leica DMIRB显微镜下对萌发的浮游植物进行定性定量分析,萌发的浮游植物数量以每克干泥中萌发的浮游植物细胞数表示(cells · g-1).浮游植物分析鉴定根据文献(郭玉洁等,2003;Hallegraeff et al., 2003)进行,一些不确定种仅鉴定至属.其中一种小型薄壁甲藻的鉴定,则通过分离培养的细胞株进行rDNA序列测定,与GenBank上相关序列进行比对分析,结合扫描电子显微镜观察,确定为Biecheleriopsis adriatica.
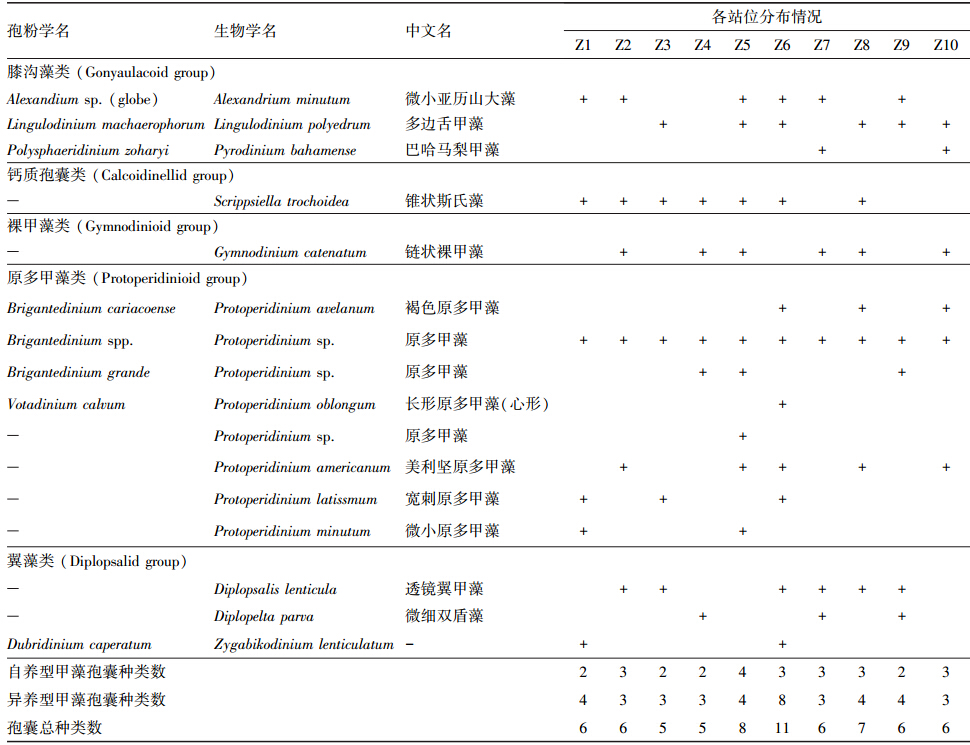
3 结果(Results) 3.1 柘林湾表层沉积物中甲藻孢囊的分布 3.1.1 甲藻孢囊的种类组成柘林湾海域表层沉积物中共分析鉴定出甲藻孢囊16种(表 1),其中,膝沟藻类3种、钙质孢囊类1种、裸甲藻类1种、原多甲藻类8种、翼甲藻类3种,包括自养型甲藻的孢囊5种,异养型甲藻的孢囊11种.大部分站位异养型孢囊种类较为丰富,原多甲藻类孢囊是柘林湾的优势类群.
本次调查中分析鉴定出的甲藻孢囊种类数不多,每个样品出现的孢囊物种数在5~11种之间,其中,Z6站位孢囊种类数最多,其余各站位孢囊种类数均相差不大.有毒的亚历山大藻孢囊和链状裸甲藻孢囊在柘林湾表层沉积物中广泛分布,而中国海域不常见的巴哈马梨甲藻孢囊(Pyrodinium bahamense)也有出现.
| 表1 甲藻孢囊在柘林湾海域表层沉积物中的分布 Table.1 Distribution of dinoflagellate cysts in surface sediments from Zhelin Bay |
柘林湾海域10个站位表层沉积物中甲藻孢囊密度为16.59~83.37 cysts · g-1,平均密度为39.52 cysts · g-1.从分布趋势来看,孢囊密度呈现出由湾外向湾内逐渐升高的趋势(图 2).孢囊密度较高的区域分布在柘林湾北部,北方有黄冈河流入,而且大量的海水养殖基地均分布在该区域,致使孢囊密度较高;而在靠近海山岛南及汛洲岛南的海域,海域养殖业较少,孢囊密度也较小.
 |
| 图 2 柘林湾海域甲藻孢囊密度的空间分布 Fig. 2 Spatial distribution of dinocyst in surface sediments from Zhelin Bay |
甲藻具有自养型和异养型两种营养类型,膝沟藻类、钙质孢囊类的全部物种及裸甲藻类中的链状裸甲藻、旋沟藻和哈曼褐多沟藻属于自养型甲藻,而全部原多甲藻类、翼藻类及裸甲藻类中的多沟藻属于异养型甲藻.异养型甲藻孢囊在数量上占总孢囊数量的百分比较高,平均为58.1%,多出现在养殖密集区域的Z1、Z5、Z6、Z8、Z9等站位(图 3).
 |
| 图 3 柘林湾海域各站位自养型、异养型甲藻孢囊及活体甲藻孢囊与空孢囊百分比 Fig. 3 Proportions of cysts of autotrophic,heterotrophic dinoflagellates,living and empty cysts in Zhelin Bay |
在表层沉积物中存在的孢囊集群中有未萌发的活体孢囊和已萌发的空孢囊,活体孢囊一般为水体中新形成的孢囊,内有细胞内含物,经历了强制性休眠期后能够萌发成营养细胞;空孢囊则是萌发后的孢囊,两者数量及其百分比则可反映孢囊萌发状况.本研究中空孢囊百分比均较高(图 3),平均百分比达到56.3%,说明大部分孢囊已经萌发.Z6站位的活体孢囊密度最高,为35.73 cysts · g-1,Z8站位的空孢囊密度最高,为58.24 cysts · g-1.
3.2 萌发的浮游植物群落结构 3.2.1 萌发的浮游植物种类组成柘林湾海域表层沉积物共萌发出浮游植物26属33种.其中,硅藻19种,占总种数的57.6%,以拟菱形藻(Pseudonitzschia spp.)、角毛藻(Chattoceros spp.)、中肋骨条藻(Skeletonema costatum)、圆筛藻(Coscinodiscus spp.)等为优势种;甲藻8种,有毒有害赤潮藻类种类较丰富,包括亚历山大藻、斯氏藻、裸甲藻等常见的有害生物,一种woloszynskioid类甲藻Biecheleriopsis adriatica是主要优势种;其他浮游植物种类如蓝细菌、绿藻、硅鞭藻等也有出现.20 d时萌发出的种类数较多,总种类数为26种,而且甲藻和硅藻种类数也较多;而40 d时萌发出的总种类数仅为18种.
3.2.2 萌发的浮游植物密度萌发的浮游植物总细胞密度如图 4a所示.20 d萌发出的浮游植物细胞密度为49~11436 cells · g-1(以干重计,下同),平均为2015 cells · g-1;最高值出现在Z4站位,优势种主要为蓝细菌中的色球藻(Chroococcus spp.).40 d萌发出的浮游植物细胞密度明显高于20 d,为70~113538 cells · g-1,平均为31266 cells · g-1.Z1、Z4站位浮游植物密度较高,优势种分别为定鞭藻中的金色藻(Chrysochromulina spp.)和绿藻中的粘四集藻(Palmella mucosa).
 |
| 图 4 柘林湾海域表层沉积物萌发培养20 d和40 d后各浮游植物细胞密度(a.浮游植物总细胞,b.硅藻细胞,c.甲藻细胞,d.其他类别浮游植物) Fig. 4 Cell number of germinated phytoplankton(a),diatoms(b),dinoflagellates(c),phytoplankton other than diatoms and dinoflagellate(d)after 20 d and 40 d inoculation of surface sediments from Zhelin Bay |
硅藻是萌发浮游植物群落的主要优势类群,整体来说,20 d萌发的硅藻数量较低(图 4b).20 d萌发中Z2站位硅藻密度较高,为621 cells · g-1,优势种主要为拟菱形藻;40 d萌发中Z7站位硅藻细胞密度最高,为7630 cells · g-1,主要优势种也为拟菱形藻.Z3站位萌发的硅藻最少,20 d和40 d萌发后,硅藻密度均低于20 cells · g-1.
萌发的甲藻细胞密度远远低于硅藻,大部分站位的甲藻细胞密度小于100 cells · g-1(图 4c).20 d萌发后,Z6站位的甲藻细胞密度最高,为229 cells · g-1,Biecheleriopsis adriatica为优势甲藻,Z4站位中未见甲藻出现.40 d萌发后,甲藻细胞密度更低,最高值仅为64 cells · g-1,且有4个站位未见有甲藻出现.
其他类别的浮游植物在萌发的浮游植物中大量出现,特别是40 d萌发后密度更高(图 4d).优势种类主要是一些微微型(2~20 μm)种类,如蓝藻中的色球藻、定鞭藻中的金色藻、球形棕囊藻(Phaeocystis globosa)及绿藻中的粘四集藻等,如金色藻在Z1和Z4站位40 d萌发中大量出现,密度分别达50091和32603 cells · g-1;球形棕囊藻也在Z4和Z5站位40 d萌发中大量出现,细胞密度达17784和6962 cells · g-1;而粘四集藻则在各站位的40 d萌发中均大量出现.
4 讨论(Discussion)4.1 甲藻孢囊分布特征与海洋环境的关系
本研究中柘林湾海域10个站位仅观察到甲藻孢囊16种,孢囊密度均低于100 cysts · g-1,平均密度小于50 cysts · g-1,甲藻孢囊种类丰富程度和密度要明显低于2001年的调查结果(王朝晖等,2004).甲藻孢囊常在颗粒大小、沉降性质与之相近的粘土中积累,颗粒较细的粘性沉积物中甲藻孢囊密度较高(Yamaguchi et al., 1995).本调查孢囊种类和密度低的原因主要有:①本次调查区域位于黄冈河口、海山岛附近,沉积物中砂质较多,不利于孢囊的沉降和积累;而且黄冈河带来大量悬浮物质及养殖区的残饵沉积,加快了沉积物的沉降,使得孢囊被稀释,因而密度降低.而2001年调查的站位主要位于柘林镇附近养殖区,除了养殖污染外,生活污染也较为严重,初级生产力较高,因而孢囊产量也较高.②2001年的调查中,分析的是长度为20 cm左右的垂直沉积物样品,是近15~20年来孢囊沉降积累的结果,因而孢囊种类多样性也较高,而且2001年调查中每个样品中的种类数也不是很高.③本调查是表层沉积物样品,由于沉积物扰动和底栖动物的摄食,可能会造成某些孢囊的损失.
异养型甲藻孢囊的百分比是孢囊组成的重要部分.异养型甲藻属于消费者,需要摄食其他小型浮游植物(一般为小型硅藻)才能生长,根据生态系统的能量流动规律,较高营养级生物的生长速度和生物量均较初级生产者低,因此,异养型甲藻及其孢囊的产量应远低于自养型物种.在孢囊集群中,如果异养型甲藻孢囊百分比上升,说明其食物来源丰富,也就是说小型硅藻类数量丰富,从而能在一定程度上表示海域较高的富营养化程度(Matsuoka,1999).本研究中,柘林湾表层沉积物中异养型甲藻的孢囊百分比超过了50%,也在一定程度说明了该海域较高的富营养化程度和初级生产力.而实际上柘林湾水体富营养化较为严重,初级生产力高,硅藻为浮游植物主要类群(周凯等,2002).
孢囊一般需要溶解氧和适当的光强才能萌发(Anderson et al., 1987),因此,沉积物中的孢囊必需进入水体真光层才能萌发,而萌发后的孢囊壳(空孢囊)也会沉降至沉积物中.因此,空孢囊的比例还可作为孢囊萌发的依据(Kremp et al., 1999).在扰动较大的海域,空孢囊的百分比也较高.柘林湾位于亚热带海域,水温较高,营养盐含量丰富,有利于孢囊萌发,较高的空孢囊百分比说明该海域孢囊萌发率较高.同时也可能是由于研究海域处于人类活动比较剧烈的养殖海域与航道上,表层沉积物扰动剧烈,导致底层沉积物中已经萌发的空孢囊上浮至表层沉积物中.
4.2 萌发的浮游植物群落结构与赤潮发生潜势本研究中萌发的浮游植物种类多样性较低,种类丰富程度远低于同一海域浮游植物调查结果(周凯等,2002),而甲藻萌发的种类数也明显低于沉积物中孢囊的种类数.分析其原因,首先是由于并非所有浮游植物都能形成孢囊或休眠体,因此,休眠体种类丰富程度应较浮游植物低.其次,本研究中沉积物休眠体分别经过20、40 d的萌发培养,萌发出的营养细胞经历了一段时间的生长繁殖,生长速度快的细胞能够在短时期内迅速增长,成为营养细胞的优势种类,而其它竞争力差的种类在生长后期则慢慢消失,导致观察到的种类较少,并且优势种类均为一些生长速度较快的小型种类,特别是40 d萌发培养后.此外,影响萌发的因素很多,包括温度、溶解氧、营养盐、光照强度等环境条件(Kim et al., 2000)及生物内在节律(Matrai et al., 2005)等,可能导致沉积物中某些孢囊或休眠体不能在实验室设定的培养条件下萌发.
在本研究中一种woloszynskioid类甲藻Biecheleriopsis adriatica是萌发甲藻的优势种,但在沉积物中却未观察到该藻孢囊的存在.可能是由于对该藻生活史和孢囊形态的认识不足,不能辨别其孢囊,而在该藻培养过程中也未能观察到其孢囊的形成,但Moestrup等(2009)曾观察到了其疑似孢囊.Biecheleriopsis adriatic是一类薄壁甲藻类,细胞表面由大量小而薄的甲片覆盖(Moestrup et al., 2009),在光学显微镜下很难观察到其甲片,在形态上与裸甲藻非常相似,因此,通常被错误判断为裸甲藻类(Moestrup et al., 2009;Takahashi et al., 2014).最近有一些对这类薄壁甲藻进行深入的形态学和分子生物学研究的报道,并建立了若干个新属,Biecheleriopsis就是其中之一(Lindberg et al., 2005; Hansen et al., 2007; Moestrup et al., 2008,2009),而在以往的研究报道中将其定名为Gymnodinium corri或G. pygmaeum等(Moestrup et al., 2009).Biecheleriopsis adriatica是中国海域的新记录种,这可能与个体微小、细胞壁薄有关,在常规的浮游植物观察中常常被忽视或者被误定,估计在以往研究报道中与之大小相近的裸甲藻类可能都是该藻.而在本研究萌发浮游植物群落中出现了大量的Biecheleriopsis adriatica,说明其应该是柘林湾浮游植物常见种类,而在广东沿海其他沉积物萌发实验中该藻(鉴定为Gymnodinium corri)也为优势甲藻(康伟等,2009; Wang et al., 2013).因此,应密切关注其赤潮发生风险.
萌发的营养细胞中也出现了大量的其他类别的浮游植物,蓝藻、绿藻及定鞭藻在某些站位的细胞数量都相当丰富,而在以往的浮游植物调查中几乎未有这些种类的报道.它们在萌发的浮游植物中大量出现,说明它们理应是柘林湾海域浮游植物常见种类,但由于浮游植物调查采样时间间隔及样品采集量的局限性,有些浮游植物可能未在常规浮游植物调查中出现.而沉积物中休眠体是上层水体所有浮游植物休眠体长时间积累而成,通过沉积物的萌发,可以在一定程度上丰富浮游植物多样性.
虽然硅藻也是萌发浮游植物的优势类群,但萌发的浮游植物群落中硅藻的百分比远远低于浮游植物调查结果(周凯等,2002).萌发浮游植物组成中硅藻含量的降低首先与柘林湾的气候和环境条件有关,柘林湾气候温和,营养盐含量丰富,硅藻可以以营养细胞形式越冬或度过短暂的营养盐缺乏时期,故而硅藻孢子形成率较低.此外,在萌发培养前期,硅藻的大量生长能致使培养基中硅缺乏,导致一些非硅藻类细胞在萌发培养40 d后大量出现.而其中的优势种类球形棕囊藻也是柘林湾的常见赤潮藻类,其赤潮曾在1997年10月至1998年2月发生(陈菊芳等,1999),而1999年和2002年又发生了该藻赤潮(周凯等,2002;徐宁等,2003),球形棕囊藻赤潮的循环发生说明了休眠体萌发对该藻赤潮贡献.
5 结论(Conclusions)1)柘林湾海域甲藻孢囊种类多样性较低,孢囊密度也较低,但有毒有害赤潮藻类亚历山大藻和链状裸甲藻的孢囊分布广泛.
2)孢囊组成以异养型甲藻的孢囊占优势,显示了该海域较高的富营养化程度;空孢囊百分比含量较高,说明大部分孢囊已经萌发.
3)萌发的浮游植物种类数较少,以小型硅藻占据优势,同时还出现了大量金色藻及球形棕囊藻等其他类别的潜在有毒种类以及我国海域新记录种Biecheleriopsis adriatica,应密切关注这些藻类赤潮的潜在风险.
| [1] | Anderson D M, Taylor C D, Armbrust E V. 1987. The effects of darkness and anaerobiosis on dinoflagellate cyst germination [J]. Limnology and Oceanography, 32(2): 340-351 |
| [2] | 陈菊芳, 王艳, 徐宁, 等. 1999. 中国赤潮新记录种—球形棕囊藻(Phaeocystis globosa)[J].暨南大学学报, 20(3): 124-129 |
| [3] | Genovesi L, de Vernal A, Thibodeau B, et al. 2011. Recent changes in bottom water oxygenation and temperature in the Gulf of St Lawrence: Micropaleontological and geochemical evidence [J]. Limnology and Oceanography, 56(4): 1319-1329 |
| [4] | 郭玉洁, 钱树本. 2003. 中国海藻志(第五卷)硅藻门,第一册中心纲[M].北京: 科学出版社 |
| [5] | Hallegraeff G M,Anderson D M,Cembella A D.2003.Manual on Harmful Marine Microalgae[M].Paris: UNESCO Publishing |
| [6] | Hansen G, Daugbjerg N, Henriksen P. 2007. Baldinia anauniensis gen. et sp. nov.: a 'new’ dinoflagellate from Lake Tovel, N. Italy [J]. Phycologia, 46: 86-108 |
| [7] | Hessler I, Young M, Holzwarth U, et al. 2013. Imprint of eastern Indian Ocean surface oceanography on modern organic-walled dinoflagellate cyst assemblages [J]. Marine Micropaleontology, 101: 89-105 |
| [8] | 黄长江,董巧香,郑磊.1999.1997年底中国东南沿海大规模赤潮原因生物的形态分类与生态学特征[J].海洋与湖沼, 30: 581-590 |
| [9] | 康伟, 王朝晖, 付永虎,等. 2009. 大亚湾沉积物中浮游植物休眠体萌发研究[J].中国环境科学, 29(12): 1121-1127 |
| [10] | Kim Y O, Han M S. 2000. Seasonal relationships between cyst germination and vegetative population of Scrippsiella trochoidea (Dinophyceae) [J]. Marine Ecology Progress Series, 204: 111-118 |
| [11] | Kremp A, Heiskanen S.1999. Sexuality and cyst formation of the spring-bloom dinoflagellate Scrippsiella hangoei in the coastal northern Baltic Sea [J]. Marine Biology, 134(4): 771-777 |
| [12] | Kremp A, Anderson D M. 2000. Factors regulation germination of calcified cysts of the spring bloom dinoflagellate Scrippsiella hangoei from the northern Baltic Sea [J]. Journal of Plankton Research, 22(7): 1311-1327 |
| [13] | Lindberg K, Moestrup , Daugbjerg N. 2005. Studies on woloszynskioid dinoflagellates I: Woloszynskia coronata re-examined using light and electron microscopy and partial LSU rDNA sequences, with description of Tovellia gen. nov. and Jadwigia gen. nov. (Tovelliaceae fam. nov.) [J]. Phycologia, 44: 416-440 |
| [14] | Liu D Y, Shen X H, Di B P, et al.2013. Palaeoecological analysis of phytoplankton regime shifts in response to coastal eutrophication [J]. Marine Ecology Progress Series, 475: 1-14 |
| [15] | Matrai P, Thompson B, Keller M. 2005. Circannual excystment of resting cysts of Alexandrium spp from eastern Gulf of Maine populations [J]. Deep-Sea Research II, 52(19/21): 2560-2568 |
| [16] | Matsuoka K. 1999. Eutrophication process recorded in dinoflagellate cyst assemblages-a case of Yokohama Port, Tokyo Bay, Japan [J]. The Science of the Total Environment, 231(1): 17-35 |
| [17] | Matsuoka K, Fukuyo Y. 2000. Technical guide for modern dinoflagellate cyst study[M].Tokyo: Asian Natural Environmental Science Center |
| [18] | Moestrup, Hansen G., Daugbjerg N. 2008. Studies on woloszynskioid dinoflagellates III: on the ultrastructure and phylogeny of Borghiella dodgei gen. et sp. nov., a cold-water species from Lake Tovel, N. Italy, and on B. tenuissima comb. nov. (syn. Woloszynskia tenuissima) [J]. Phycologia 47: 54-78 |
| [19] | Moestrup, Lindberg K, Daugbjerg N. 2009. Studies on woloszynskioid dinoflagellates V. Ultrastructure of Biecheleriopsis gen. nov. Phycological Research, 57: 221-237 |
| [20] | Prebble J G,Crouch E M,Carter L, et al.2013. An expanded modern dinoflagellate cyst dataset for the Southwest Pacific and Southern Hemisphere with environmental associations [J]. Marine Micropaleontology, 101: 33-48 |
| [21] | Takahashi K, Sarai C, Iwataki M. 2014. Morphology of two marine woloszynskioid dinoflagellates, Biecheleria brevisulcata sp. nov. and Biecheleriopsis adriatica (Suessiaceae, Dinophyceae), from Japanese coasts [J]. Phycologia, 53: 52-65 |
| [22] | 王朝晖,陈菊芳,徐宁,等. 2001.大亚湾裸甲藻种群的季节变化与环境条件的关系[J].生态学报, 21(11): 1825-1832 |
| [23] | 王朝晖. 2007.中国沿海甲藻孢囊与赤潮研究[M].北京:海洋出版社 |
| [24] | Wang Z H,Qi Y Z,Yang Y F.2007.Cyst formation: an important mechanism for the termination of Scrippsiella trochoidea (Dinophyceae) bloom [J].Journal of Plankton Research, 29(2): 209-218 |
| [25] | Wang Z H,Fu Y,Kang W,et al. 2013.Germination of phytoplankton resting cells from surface sediments in two areas of the southern Chinese coastal waters [J].Marine Ecology, 34(2): 218-232 |
| [26] | 徐宁,齐雨藻,陈菊芳,等. 2003.球形棕囊藻(Phaeocystis globosa Scherffel)赤潮成因分析[J].环境科学学报, 23(1): 113-118 |
| [27] | Yamaguchi M, Itakura S, Imai I. 1995. Vertical and horizontal distribution and abundance of resting cysts of the toxic dinoflagellate Alexandrium tamarense and Alexandrium catenella in sediments of Hiroshima Bay, the Seto Inland Sea, Japan [J]. Nippon Suisan Gakkaishi, 61(5): 700-706 |
| [28] | 周凯,黄长江,姜胜,等.2002.2000—2001年柘林湾浮游植物群落结构及数量变动的周年调查[J].生态学报, 22(5): 688-698 |
| [29] | Zonneveld K A F, Chen L, Elshanawany R, et al. 2012. The use of dinoflagellate cysts to separate human-induced from natural variability in the trophic state of the Po River discharge plume over the last two centuries [J]. Marine Pollution Bulletin, 64(1): 114-132 |