 2014, Vol. 34
2014, Vol. 34



2. 西北大学城市与环境学院, 西安 710127;
3. 陕西师范大学旅游与环境学院, 西安 710062
2. College of Urban and Environmental Sciences, Northwest University, Xi'an 710127;
3. Tourism and Environment College, Shaanxi Normal University, Xi'an 710062
因此,本研究基于垂向渗透系数室内模拟试验,在试验管中加入不同生物密度的颤蚓,利用回归分析及趋势分析来研究不同生物扰动作用强度下沉积物渗透性的变化特性,以正确认识生物扰动对河床沉积物渗透系数变化的影响机理.这不仅在理论上有助于人们深化对河床渗透性能的科学认识,而且对研究人为控制地表水对地下水污染,恢复河流水文生态功能,维持河流健康等具有重要的指导意义.
2 材料与方法(Materials and methods) 2.1 试验材料沉积物、颤蚓生物、上覆水均采集于渭河干流(眉县、咸阳、草滩、临潼和华县)潜流带,地点相同.沉积物是在做完原位渗透试验后,采集的有机玻璃管内沉积物,取样深度约45 cm,数量充足;颤蚓利用采泥器(20 cm×30 cm)、D型网、采样筛工具采集,采样深度约15 cm,数量充足;上覆水鉴于河水含沙量较大,试验使用自然澄清后的渭河河水,数量充足.此外,用到的实验仪器还有长40 cm、内径5.4 cm的透明有机玻璃管材,14孔的定制铁架台、定制水槽、秒表、分样筛、纱布、卡子等.
2.2 模拟试验试验分为1、2两组,每组7根模拟试验管.模拟试验管装有采集到的沉积物样品,利用分样筛把自然风干后的沉积物分为粗沙、细沙和浮泥,再按m(粗砂)∶ m(细砂)∶ m(浮泥)=4 ∶ 3 ∶ 3的比例充分混合,混合沉积物厚度约为35 cm;试验管中加入混合沉积物后,再在试验管中加入经过自然澄清后的原位采集到的渭河河水,静置不少于24 h,使沉积物中含水能够充分饱和;沉积物含水充分饱和后在7根试验管中加入不同生物密度的颤蚓(0、0.5、1.0、1.5、2.0、2.5和3.0 ind. · cm-2),试验用颤蚓采集于渭河干流,将试验装置放置不少于24 h后再进行渗透系数测定(孙刚等,2009);渗透系数每1 h测定1次,共测定3 d;根据试验时间,设定6:00—20:00为白天,20:00—6:00为夜晚.另外,由于7根试管中加入的颤蚓密度不同,根据不同密度标试验编号为MN0(对应密度为0 ind. · cm-2)、MN1(0.5 ind. · cm-2)、MN2(1.0 ind. · cm-2)等,其他类推.根据试验分组分为MN0-1、MN0-2、MN1-1、MN1-2等,其中,MN0-1、MN1-1等为第1组,MN0-2、MN1-2等为第2组.
2.3 渗透系数测定方法渗透系数的测定方法采用水头下降竖管渗透试验法(Chen,2000; Chen et al., 2010).试验采用的竖管是长80 cm、内径5.4 cm的透明有机玻璃管;距上端35 cm左右处固定试验管,下端浸入水中;管内沉积物厚度约35 cm,上部注满经过自然澄清后的原位采集到的渭河河水;准备工作完成后,记录水头下降过程中不同时段的水头高度,用来计算垂向渗透系数(图 1).
 |
| 图 1 室内垂向渗透系数测试示意 Fig.1 Measurement of vertical hydraulic conductivity in lab testing process |
模拟试验结束后,对不同模拟管沉积物垂向渗透系数(Kv)进行计算,其计算公式为(Chen,2000):
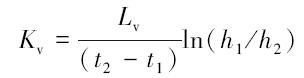
式中,Lv为有机玻璃管中沉积物的厚度(cm);h1和h2分别为t1和t2时刻测试管中的水位值(cm).
2.4 数据处理运用Excel和SPSS等软件对试验结果进行统计和分析,分析包括回归分析和趋势分析.
3 结果(Results) 3.1 生物扰动影响垂向渗透系数的实验模拟 3.1.1 不同时间垂向渗透系数变化从2013年7月9日20时至2013年7月12日20时,72 h共内记录了67次数据,第1个24 h记录了24次,第2个24 h记录了22次,第3个24 h记录了21次.通过对所测的数据进行渗透系数计算,并求算数平均数,计算出每个完整24 h内的垂向渗透系数和每个完整24 h内白天和晚上的垂向渗透系数(表 1).不同颤蚓密度下垂向渗透系数的值为第1、2组的均值,其中,MN0以MN0-1测定的垂向渗透系数表征(测定前MN0-2沉积物表层被人为破坏).对3个昼夜过程中计算的昼、夜6个垂向渗透系数进行相关分析,除MN5线性相关系数为0.63外,其余测定管线性相关系数均大于0.86(表 2),存在明显的线性相关.随着试验时间的增加,垂向渗透系数逐渐变大(表 1),说明在环境基本不变情况下的一段时间范围内,沉积物的渗透性是呈线性逐渐增加,颤蚓的扰动作用越来越明显.
| 表 1 不同时间垂向渗透系数 Table 1 Test value of vertical hydraulic conductivity within different times |
| 表 2 白天和晚上垂向渗透系数线性相关系数 Table 2 Linearly dependent coefficient of vertical hydraulic conductivity within daytime and nighttime |
底栖动物的生物扰动方式包括生物构造、生物冲灌、生物活动等(杨群慧等,2008).汪振华等(2011)在研究鱼类聚集时发现,鱼类在白天和晚上存在不同的集聚情况.由于昼夜变化,不同生物在白天和夜晚表现出不同的活动规律.颤蚓活动在昼夜变化下也应存在一定变化.鉴于在环境基本不变情况下,沉积物的渗透性呈线性增加,为更好地分析颤蚓生物扰动下沉积物白天和夜晚垂向渗透系数的大小,以1天24小时作为1个测试循环周期,分别计算白天-夜晚(先测白天渗透系数)2个循环均值和夜晚-白天(先测夜晚渗透系数)3个循环均值(表 3).结果表明:对于白天-夜晚循环,白天垂向渗透系数小于夜晚,而对于夜晚-白天循环,其结果恰好相反;白天渗透系数先测,白天测定值小,晚上渗透系数先测,夜晚测定值小;光照等因素对颤蚓扰动作用影响较弱.沉积物渗透性大小在一定时间范围内上呈线性增加(表 1、表 2),光照等因素对颤蚓扰动作用影响较弱,白天测定值大于前一个晚上测定值,小于后一个晚上测定值(表 1).说明在一定时间范围内,在颤蚓生物扰动作用影响下,渗透系数在白天和夜晚的变化程度小于渗透系数在整个研究时段内的变化程度,光照条件对颤蚓扰动作用的影响不明显.这可能是因为颤蚓的前端藏在潜流带中,尾部露在水中摆动,光照对藏于潜流带的前端影响很小;颤蚓的耐污性非常强,主要要栖息于水质较差的河流底部,光照在水质较差河水中的透过率较低,光照对颤蚓的影响也很小;颤蚓主要通过进食含有有机质的沉积物生活,对光照需求较少.这进一步在一定程度上导致光照条件对颤蚓生物扰动作用的影响不明显.
| 表 3 白天和夜晚垂向渗透系数 Table 3 Measured value of vertical hydraulic conductivity within daytime and nighttime |
 |
| 图 2 不同颤蚓密度条件下沉积物垂向渗透系数 Fig.2 Vertical hydraulic conductivity of different Tubificid densities |
通过对不同颤蚓密度下沉积物垂向渗透系数的计算分析,发现随着颤蚓密度的增加,沉积物渗透系数先变大再变小,并在MN2(1.0 ind. · cm-2)附近出现极大值(图 2).试管MN2垂向渗透系数测定值为9.63 cm · d-1,在7个不同颤蚓密度试验中为最大值(表 1).MN1、MN2、MN3和MN4的测定值均大于MN0,MN5和MN6的测定值均小于MN0(表 1、图 2).表明颤蚓密度在2.0 ind. · cm-2以下时,对沉积物渗透性能具有明显增强作用,反映出在一定颤蚓密度下,颤蚓生物扰动作用对沉积物渗透性呈增强作用.而当颤蚓密度进一步加大后,其沉积物渗透性呈减弱作用,甚至密度达到一定程度时,沉积物渗透性比没有颤蚓时还弱.研究表明,生物扰动作用在一定密度范围内随着生物密度的增加而增强(聂小保等,2010).MN6中颤蚓密度为3.0 ind. · cm-2,为所有模拟试验管中生物密度最大的,进而为生物扰动作用最强的,但其垂向渗透系数并不是所有模拟试验中最大的.由此可以看出,颤蚓生物扰动作用在达到一定程度后对加强沉积物渗透性起到负作用.这可能是由颤蚓生物扰动作用改变了沉积物物理性质引起的.
3.2 生物扰动影响沉积物变化的实验模拟 3.2.1 沉积物表层特征变化3个完整昼夜测定垂向渗透系数结束后,利用尼康D5000分别对试验管进行俯视和正视拍照(图 3),以分析沉积物的表层变化.通过观察与对比分析可以看出:MN0-1沉积物表面比较光滑,有起伏;MN1-1沉积物表面俯视图中能够看到明显的坑洞;MN2-1、MN3-1、MN4-1、MN5-1和MN6-1等试验管沉积物均存在坑洞,数量多但细小,且高颤蚓密度试验管中表层沉积物稳定性较差;沉积物表层随着颤蚓密度的增大而逐渐变得粗糙,对沉积物的影响作用越来越大,但MN6-1沉积物表面坑洞被细小沉积物覆盖,从而使其沉积物俯视图显得相对光滑.颤蚓的前段藏于沉积物中形成孔洞,后端在水中摇摆,摇摆作用使孔洞顶部的沉积物减少,在一定程度上加大了表层的坑洞,正如MN1-1中看到的情况一样;而随着颤蚓密度的增加,颤蚓的摆动作用也越来越明显,摆动作用加大坑洞的同时又被其他坑洞覆盖,使其不能形成相对稳定且明显的坑洞,也正如MN5-1和MN6-1所看到的.加入不同密度颤蚓的试验管沉积物表层均比未加入颤蚓试验管的沉积物表层粗糙,且粗糙程度越来越大,但MN5-1和MN6-1由于颤蚓密度相对较大,颤蚓生物扰动作用增强,大量颤蚓的扰动作用使沉积物稳定性较弱,使沉积物表层不能形成大量稳定的坑洞,使其沉积物表层相对光滑.颤蚓生物扰动减小沉积物细小颗粒物的阻塞作用呈现先增强后减弱的规律,进而使沉积物渗透性呈现先增强后变弱的趋势,渗透系数测定值也证明了这一结果(表 1、图 2).
 |
| 图 3 不同颤蚓密度条件下沉积物表层结构变化 Fig.3 Changes in surface sediments structure with different Tubificid densities |
开始测定渗透系数前和3 d后渗透系数测试结束时分别对各个试验管沉积物厚度进行了测量,并对试验前后沉积物厚度减少量进行计算和分析(图 4).沉积物厚度在室内模拟试验结束后均比试验前有所下降,且随着颤蚓密度的增加变化越来越大,厚度差的变化越来越明显(图 4).颤蚓主要通过进食含有有机质的沉积物和摇摆尾端完成其生命活动,在进食和摇摆尾部过程中能够改变原有沉积物颗粒大小,沉积物颗粒大小变化在一定程度上引起沉积物厚度差变化.随着颤蚓密度增大,试验前后沉积物厚度变化越来越大,也在一定程度上证明,颤蚓生物扰动作用随着颤蚓密度的增加而增大.
 |
| 图 4 不同颤蚓密度沉积物厚度变化 Fig.4 Sediment thickness variation with different Tubificid densities |
 |
| 图 5 不同颤蚓密度下沉积物成分变化趋势 Fig.5 Variation of weight percentage for particle composition of measured sediment in different Tubificid densities |
鉴于本次模拟试验中,颤蚓生物扰动深度集中在0~10 cm之间,重点对这一深度的沉积物进行了粒度分析,并计算了不同颤蚓密度条件下的沉积物成分百分比(表 4).并通过线性回归分析,得出沉积物中3种不同成分的趋势线(图 5).由表 4可知,7个不同颤蚓密度下沉积物组成成分百分数均值分别是:浮泥31.55%、细沙24.54%、粗沙43.91%,成分比与试验前沉积物测定基本一致.这在一定程度上反映颤蚓生物扰动能够改变沉积物粒度大小,进而影响沉积物成分比,成分比基本一致说明这一影响并非非常明显.浮泥百分比随着颤蚓密度的增大而不断增加,粗砂和细沙百分比不断减少(图 5).浮泥含量增加,粗沙和细沙含量减少,说明在颤蚓扰动作用下沉积物颗粒变小,这也在一定程度验证了沉积物厚度变化的原因.此外,也说明随着颤蚓密度的增加,颤蚓生物扰动作用逐渐加强,并破坏了原来的沉积物,使沉积物颗粒逐渐变小(图 5).不同颤蚓密度下,颤蚓生物扰动的影响效果不同,颤蚓密度越大,影响效果越大,沉积物成分变化越大.
| 表 4 不同颤蚓密度下沉积物成分质量百分比 Table 4 Weight percentage of particle composition of measured sediment with different Tubificid densities |
颤蚓生物扰动能够改变沉积物的渗透系数,在渗透环境基本一致的情况下,沉积物中包含的颤蚓密度不同,其渗透系数表现出明显差异,且光照条件对颤蚓生物扰动作用的影响较小.随着颤蚓生物扰动时间的增加,渗透系数均呈现增大趋势,说明在一定时间范围内,随着试验时间增加,颤蚓生物扰动对渗透系数的影响效应也增加.但鉴于颤蚓的生活习性,光照难以到达河水底部甚至潜流带中,导致光照条件变化对颤蚓生物扰动作用的影响不是非常明显,而室内模拟实验的实测值也证明了这一点.此外,颤蚓密度小于一定量值时,沉积物渗透系数均大于控制管MN0测定值的情况;颤蚓密度大于一定量值时,沉积物渗透系数出现小于控制管MN0测定值;这一量值范围在2.0~2.5 ind. · cm-2之间;颤蚓密度在1.0 ind. · cm-2附近时渗透系数出现最大值.任朝亮等(2013)在研究底栖动物扰动对河床渗透性影响的现场试验中发现,在底栖动物密度小于1.0 ind. · cm-2的范围内,渗透系数随着底栖动物密度的增加而增加;聂小保等(2010)研究发现,颤蚓密度和颤蚓生物扰动之间呈现“N”型关系,而在密度小于4.0 ind. · cm-2的范围内两者的关系呈现“n”型;均和本次研究结果相似.从中可以看出,渗透系数在一定颤蚓密度下,随着颤蚓数量、密度的增加而逐渐变大,当超过这一密度后,渗透系数会变小.这是因为一方面随着密度增加,颤蚓首先通过增加迁移应对食物短缺,然后通过减小迁移避免能量消化应对食物短缺,进而影响沉积物渗透系数(聂小保等,2010);另一方面,当颤蚓密度达到一定程度后,其颤蚓的扰动作用破坏了沉积物的粒度分布,使沉积物成分发生变化,进而使测定的沉积物渗透系数发生变化,同时影响沉积物的渗透性(图 3、图 5).
4.2 颤蚓生物扰动对沉积物影响随着颤蚓密度的增大,颤蚓生物扰动对沉积物的影响越来越大,沉积物表面变得越来越粗糙,且沉积物稳定性逐渐变弱(图 3).沉积物的厚度变化随着颤蚓密度的增大而逐渐变大,投放颤蚓的MN1~MN6的沉积物厚度差比MN0的沉积物厚度差多50%~400%(图 4).并且沉积物成分随着颤蚓密度的增大变化逐渐明显,沉积物粒度有逐渐变小的趋势,其中,MN6的浮泥含量比MN0多4.7%,其他试验管均有不同变化(表 4、图 5).以上均证明了随着颤蚓密度的增大,沉积物变化越来越明显,也即颤蚓生物扰动作用随着颤蚓密度的增加而逐渐变大.一般认为,沉积物的渗透性主要由沉积物的颗粒大小决定,沉积物颗粒越大,渗透性也越好(宋进喜等,2009),反之,渗透性越弱.沉积物成分随着颤蚓密度的增大,变化逐渐明显,且沉积物粒度有逐渐变小的趋势(图 5).沉积物粒度变小,浮泥含量增加,粗沙、细沙含量变小,在一定程度上能够减小沉积物的渗透性.颤蚓前端深入沉积物中,后端在水中摇摆;深入沉积物的前端通过吞噬沉积物满足自身生命需要的同时也改变着沉积物结构和成分,使沉积物中形成交叉孔洞,浮泥含量增加;水中摇摆的后端则通过摇摆作用改变河水底部的水流环境,并影响沉积物表层的坑洞形式;沉积物在颤蚓前端和后端的不同作用下改变着沉积物的表层特性、厚度和成分.
4.3 颤蚓生物扰动对沉积物渗透性影响渗透系数随着颤蚓密度的增加呈现先增大后减小的趋势,且当颤蚓密度达到一定程度后,渗透系数反比没有颤蚓时还小(表 1、图 2).说明颤蚓生物扰动对沉积物渗透性影响是随着颤蚓密度的增大先变强后减弱,当颤蚓密度达到1.0 ind. · cm-2附近时(略大于1.0 ind. · cm-2),渗透系数出现最大值,说明此时颤蚓生物扰动对沉积物渗透性的加强作用达到最大.在底栖动物密度小于1.0 ind. · cm-2的范围内,底栖动物扰动能够加强沉积物的渗透性,且底栖动物密度和渗透系数之间存在强相关性(任朝亮等,2013).随着颤蚓密度进一步加大,颤蚓生物扰动作用逐步加强的同时,也逐渐破坏沉积物原有的稳定性,并使沉积物中浮泥含量增加,粗沙、细沙含量变小,在一定程度上减小沉积物渗透系数,进一步减弱了沉积物的渗透性.颤蚓的生物迁移和颤蚓密度之间呈现“N”型,而在小于4.0 ind. · cm-2的范围内,两者呈现“n”型.颤蚓首先通过生物迁移满足自身需求,避免食物短缺,而后通过减少生物迁移减少能量消耗来避免食物短缺(聂小保等,2010).此外,颤蚓密度达到一定程度后,其颤蚓的扰动作用破坏了沉积物的粒度分布,使沉积物成分发生变化,进而使测定的沉积物渗透系数发生变化,同时影响沉积物的渗透性.这说明颤蚓生物扰动作用在颤蚓密度小于一定程度时对沉积物渗透性有加强作用,而颤蚓密度大于一定程度后,对沉积物渗透性有减弱作用,这一程度大致范围在2.0~2.5 ind. · cm-2之间.
5 结论(Conclusions)1)河流潜流带渗透系数,在研究时间范围内,随着颤蚓生物扰动时间的增加,其值呈线性增加趋势,且渗透系数在白天和晚上的变化程度小于在整个试验时段上的变化程度.光照条件对颤蚓扰动影响不明显,沉积物的渗透系数在颤蚓密度约为1.0 ind. · cm-2时达到最大值,且颤蚓密度在2.0~2.5 ind. · cm-2之间某密度时,其沉积物渗透系数和无颤蚓生物时相同.
2)颤蚓密度越大,颤蚓生物扰动程度越大,沉积物表层、厚度、成分变化越大,沉积物颗粒有变小趋势,浮泥含量增加.
3)颤蚓生物扰动作用能够影响沉积物的渗透性能,当颤蚓密度小于一定量值时,颤蚓生物扰动对沉积物渗透性有加强作用;而颤蚓密度大于一定量值后,颤蚓生物扰动对沉积物渗透性有减弱作用;这一量值范围在2.0~2.5 ind. · cm-2之间.
4)颤蚓通过前端在沉积物中挖掘孔洞,后端在水中摇摆吸氧,并为满足自身生命需要吞食沉积物或向更深的沉积物迁移等生物扰动形式,来改变沉积物的内部和表层结构、沉积物成分组成等,以此缓解或加强沉积物中细小颗粒物的阻塞,并使沉积物的渗透性增强或减弱.
| [1] | Alex ander M D, Caissie D. 2003. Variability and comparison of hyporheic water temperatures and seepage fluxes in a small Atlantic salmon stream[J]. Groundwater, 41(1): 72-82 |
| [2] | 白秀玲,周云凯,张雷.2012.水丝蚓对太湖沉积物有机磷组成及垂向分布的影响[J].生态学报, 32(17): 5581-5588 |
| [3] | Battin T J, Kaplan L A, Newbold J D, et al. 2003. A mixing model analysis of stream solute dynamics and the contribution of a hyporheic zone to ecosystem function [J]. Freshwater Biology, 48(6): 995-1014 |
| [4] | Blaschke A P, Steiner K H, Schmalfuss R, et al. 2003. Clogging processes in hyporheic interstices of an impounded river, the danube at Vienna, Austria [J]. International Review of Hydrobiology, 88(3/4): 397-413 |
| [5] | Brunke M,Gonser T.1997.The ecological significance of exchange processes between river and groundwater[J].Freshwater Biology, 37(1): 1-33 |
| [6] | Chen X H.2000.Measurement of streambed hydraulic conductivity and its anisotropy [J]. Environmental Geology, 39(12): 1317-1324 |
| [7] | Battin T J, Kaplan L A, Newbold J D, et al. 2003. A mixing model analysis of stream solute dynamics and the contribution of a hyporheic zone to ecosystem function [J]. Freshwater Biology, 48(6): 995-1014 |
| [8] | 陈友媛. 2006. 生物活动对黄河口底土渗流特性的影响研究.青岛:中国海洋大学. 76-97 |
| [9] | Gardenr L R, Sharma P, Moore W S. 1987. A regeneration model for the effect of bioturbation by fiddler crabs on 210Pb profiles in salt marsh sediments [J]. Journal of Environmental Radioactivity, 5(1): 25-36 |
| [10] | 路永正, 阎百兴. 2010. 颤蚓扰动作用对铅镉在沉积物-水相中迁移的影响[J].中国环境科学, 30(2): 251-255 |
| [11] | Mauclaire L, Schurmann A, Mermillod-Blondin F. 2006. Influence of hydraulic conductivity on communities of microorganisms and invertebrates in porous media: a case study in drinking water slow sand filters [J]. Aquatic Sciences, 68(1): 100-108 |
| [12] | Mermillod-Blondin F. 2011. The functional significance of bioturbation and biodeposition on biogeochemical processes at the water-sediment interface in freshwater and marine ecosystems [J]. Journal of the North American Benthological Society, 30(3): 770-778 |
| [13] | 聂小保,黄廷林,张金松,等. 2010.底泥性质和分布密度对颤蚓迁移运动的影响[J].环境科学学报, 30(1): 103-110 |
| [14] | 聂小保,吴淑娟,吴方同,等. 2011.颤蚓生物扰动对沉积物氮释放的影[J].环境科学学报, 31(1): 107-113 |
| [15] | Nogaro G, Mermillod-Blondin F, Francois-Carcaillet F, et al. 2006. Invertebrate bioturbation can reduce the clogging of sediment: an experimental study using infiltration sediment columns [J]. Freshwater Biology, 51(8): 1458-1473 |
| [16] | 任朝亮,宋进喜,杨小刚,等. 2013.底栖动物扰动对河床渗透性的影响研究[J].环境科学, 34(11): 4275-4281 |
| [17] | 宋进喜,Chen X H,Cheng C,等.2009.美国内布拉斯加州埃尔克霍恩河河床沉积物渗透系数深度变化特征[J].科学通报,54(24): 3892-3899 |
| [18] | Song J X, Chen X H,Cheng C,et al.2010.Observation of bioturbation and hyporheic flux in streambed[J]. Frontiers of Environmental Science & Engineering in China, 4(3): 340-348 |
| [19] | 孙刚,房岩,安永辉,等. 2009. 泥鳅对水田上覆水中氮素动态的生物扰动效应[J].生态与农村环境学报, 25(2): 39-43 |
| [20] | Sun G, Fang Y, Wang P, et al. 2010. Bioturbation effects of branchiura sowerbyi (Tubificidae) on the vertical transport of sedimentary particles in paddy field [J]. Agricultural Science & Technology, 11(8): 117-119 |
| [21] | 汪振华,王凯,章守宇.2011.马鞍列岛多种生境中鱼类群聚的昼夜变化[J].生态学报, 31(22): 6912-6925 |
| [22] | Wood P J,Armitage P D.1997.Biological effects of fine sediment in the lotic environment[J].Environmental Management, 21(2): 203-217 |
| [23] | 杨群慧, 周怀阳, 季福武, 等. 2008. 海底生物扰动作用及其对沉积过程和记录的影响[J].地球科学进展, 23(9): 932-941 |
| [24] | 张弛, 王树功, 郑耀辉, 等. 2010. 生物扰动对红树林沉积物中AVS和重金属迁移转化的影响[J].生态学报, 30(11): 3037-3045 |
| [25] | 覃雪波, 孙红文, 吴济舟, 等. 2010. 大型底栖动物对河口沉积物的扰动作用[J].应用生态学报, 21(2): 458-463 |
| [26] | 滕彦国, 左锐, 王金生. 2007. 地表水-地下水的交错带及其生态功能[J].地球与环境, 35(1): 1-8 |