 2014, Vol. 34
2014, Vol. 34



藻类污染是国内外许多湖泊、水库面临的共性水质问题,尤其是深水型分层水源水库,原水中藻类浓度过高会直接影响饮用水安全,并增加水厂净水处理的难度和成本(黎雷等,2008;黄廷林等,2009; Hudnell et al., 2010;崔福义等,2011).分层水源水库的藻类控制途径主要有3种:降低水体氮、磷等营养盐含量,创造低温、低光照、低溶解氧等不利的生长环境,采用化学、生物、物理等方法直接去除已生长的藻类(孙昕等,2013).近年来,扬水曝气技术在众多水库水体富营养化原位控制技术中脱颖而出(Cong et al., 2009; Cong et al., 2011a;2011b;马越等2012).扬水曝气技术能同时混合表层与底部水体并向水体充氧,可有效抑制底泥中内源污染物的释放,能利用上、下层水体的循环流动驱使藻类从水库表层被动迁移至中、下部,进而控制藻类的过量繁殖.
研究发现,多数水源水库的优势藻类是铜绿微囊藻,该藻具有悬浮生长的生理特性,特征上浮速度为0.000275 m · s-1,形成不低于0.000275 m · s-1的向下流速是扬水曝气混合控藻的关键(丛海兵等2009;Cong et al., 2011b),为此需确定不同水库水文及扬水曝气条件下的流场特性.对于分层水源水库藻类控制研究,中试和现场试验均存在很大难度,采用计算流体力学(CFD)是研究此类大尺度水环境问题的重要有力手段,而FLUENT作为一个计算功能强大的CFD软件,已经开始应用于水处理领域(Li et al., 2006;韩晓婷等,2013).
因此,本研究针对西安金盆水库扬水曝气水质改善工程典型设计条件,采用商业FLUENT软件数值模拟不同水深条件下的扬水曝气器外围流场,旨在探明水深对扬水曝气抑藻范围及效果的影响,为扬水曝气器的工程应用提供技术依据和指导.
2 数值模拟方法(Numerical simulation methods) 2.1 计算区域模型研究对象为西安金盆水库扬水曝气器外围水体,将其简化为二维轴对称计算区域模型,本文仅研究模型的右半部分.模拟中将扬水曝气器简化为一个半径为0.375 m的等直径圆柱体(图 1a),水域半径为300 m,水深为50~110 m.扬水曝气器出水口顶部距水面0.5 m,扬水曝气器进水口距水库底部6 m.当扬水曝气器和外围水体构成一个系统时,二者通过扬水曝气器底部进水口和顶部出水口相互关联;扬水曝气器的控制效果与曝气器外围流场直接相关,在模拟时视扬水曝气器为黑体,只将曝气器外围水体作为模拟研究对象,扬水曝气器顶部出水口即为模拟区域顶部进水口,扬水曝气器底部进水口即为模拟区域底部出水口,故视扬水曝气器出水口为模拟区域进口,进水口为模拟区域出口.采用GAMBIT构建扬水曝气器右侧水体的四边形结构化网格,共有45595个节点、45072个单元,图 1b表示的是80 m水深算例的进、出水口附近水域网格,其中,横、纵坐标分别表示距离扬水曝气器轴心的长度(L)和水面以下深度(H),单位为m,后图中意义相同.
 |
| 图 1 实际水库物理模型简化图(a)及模拟区域计算网格(b) Fig. 1 Simplified diagram of the physical model of reservoir(a) and mesh of upper inlet and lower outlet areas outside the simulation domain(b) |
边界条件:当扬水曝气器曝气室内的气体释放进入中间上升管后,气弹开始加速上升导致水流速度逐渐增大,随着气弹的膨胀上升,其所受阻力也逐渐增加,当阻力增加至一定程度后,水流速度开始下降,表现为出现短暂的水流速度峰值,气弹释放的周期性引起曝气器出流速度的周期性(丛海兵等,2007;Cong et al., 2011a).故水体进口采用周期性速度入口(如图 2所示),进口速度与时间的变化关系采用解析法计算(Cong et al., 2011a),通过用户自定义函数(UDF)进行定义.每个周期内进口水温为上个周期内出口水温均值,采用UDF进行定义.水体出口采用压力出口.扬水曝气器及水体边壁均采用无滑移固体壁面.轴位于水体左边缘,在计算中不需给定任何参数.
 |
| 图 2 模拟区域顶部入口速度变化图 Fig. 2 Velocities at the upper inlet of the simulation domain during a period of 270 seconds |
初始条件:根据西安金盆水库水温及水位的年际变化资料,坝前平均水深范围50~110 m,夏季分层最强,水深30 m以内变温层水温近似呈线性递减,水深30 m以下水温基本稳定不变,变温层温度梯度0.73 ℃ · m-1.水体的水温分层结构、水的密度及粘度与温度的变化关系均采用UDF定义.模拟水深分别取50、65、80、95、110 m.实际扬水曝气器运行情况与模拟情况相同,同样在入口速度峰值持续很短的时间.
本文中铜绿微囊藻的典型直径和上浮速度分别取0.5 mm和0.000275 m · s-1(丛海兵等,2009;Cong et al., 2011b).首先采用Stokes公式计算铜绿微囊藻的典型密度(许保玖,2000),然后根据扬水曝气器运行水库中的藻类浓度计算出藻类的质量比浓度,并用UDF给出初始状态下水深方向的藻类质量比浓度分布.
2.3 求解与数据分析方法求解方法:FLUENT通过有限体积法求解水动力学偏微分方程组,紊流模型采用适于高应变率及流线弯曲程度大的RNG κ-ε模型,水体传热考虑表面和水下传热(Hodges,1999)并采用UDF给定传热条件.采用一阶迎风格式(First Order Upwind Scheme)对方程进行离散,采用PISO(Pressure Implicit Splitting of Operator)算法及基于压力(Pressure Based)的隐式(Implicit)求解器,求解非稳定状态下的离散方程.
RNG κ-ε模型的控制方程如下(王福军,2004):
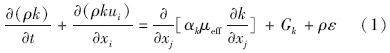
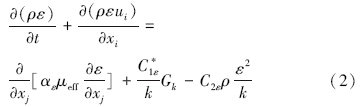


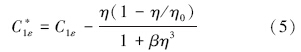
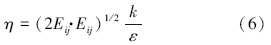
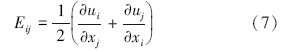
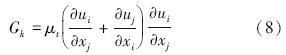
数据分析方法:根据铜绿微囊藻的悬浮生长特性,核心控藻区域定义为垂直向下速度值大于0.000275 m · s-1的区域,核心控藻区域的面积采用Photoshop软件进行计算,核心控藻区域的百分比定义为核心控藻区域与整体水域的百分比.为揭示扬水曝气过程中藻类的迁移特性,以藻类生长补偿点(水下3 m)处藻类为对象,利用FLUENT自带的迹线功能,绘制代表点处藻类的迁移轨迹.
扬水曝气器出流周期性变化导致其外围流场也周期性变化,首先根据80 m水深算例的模拟结果,一个周期内核心控藻区域的百分比从第15 s的29.56%减小到终点(270 s)的18. 84%,故确定一个周期的起点(终点)为最不利控藻时刻.
藻类迹线分析方法:为揭示扬水曝气过程中藻类的迁移特性,以藻类生长补偿点(水下3 m)处藻类为对象(丛海兵等,2009;Cong et al., 2011b),利用FLUENT自带的迹线功能,绘制代表点处藻类的迁移轨迹.记水流无质量质点的运动速度为U(x,y),由于微囊藻具有特征上浮速度(0.000275 m · s-1),故藻类实际运动横向运动速度与质点横向速度相同,而垂向运动速度为质点横向速度减去0.000275,即藻类运动速度U0(x,y)不同于质点速度U(x,y).为当流体相对稳定时,取水体中某点P0(x,y)处藻类质点为对象,绘制间隔取1 m,该质点先以点P0(x,y)的速度U0(x,y)沿U0(x,y)的矢量方向运动1 m后至点P1(x,y),再以点P1(x,y)处速度U1(x,y)运动1 m,类似运动到Pi(x,y)点,将藻类迁移的个点连线即可得到其轨迹.
3 模拟结果与分析(Results and analysis) 3.1 模拟结果的验证西安金盆水库是典型的分层水库,坝前平均水深80 m,总库容2.0亿m3,向西安市日均供水76万m3,占供水总量的70%.为控制金盆水库水体富营养化,2010年7月在坝前1.1 km范围的主库区建成扬水曝气水质改善工程,采用250~300 m的间距布置8台直径0.75 m的扬水曝气器(图 3)(马越等,2012).
 |
| 图 3 扬水曝气器分布图 Fig. 3 Layout of water-lifting aerators |
以受水库出流影响最小的4#曝气器作为模拟结果验证对象,测试断面沿4#至6#曝气器连线,该处地形较为平坦,与计算网格差别不大.曝气器运行期间,对于水温、叶绿素a等8项常规指标,采用HYDROLAB水质分析仪每日现场监测;对于水体藻类浓度,采用有机玻璃自动取样器对表层0.5 m、水下30 m和库底之上5 m的水样进行采集,保存后带回实验室进行分析,分析频率为每周1~2次,监测断面距曝气器50 m.
流速采用三维多普勒剖面流速仪(ADP,美国LAUREL公司WH600kHz)进行测量,根据模拟结果,仅在离曝气器50 m以外区域的流场不受曝气器出口的周期性出流影响.因控藻效果主要和垂向水流速度有关,故仅采用不依赖于坐标系的垂向水流速度加以验证,测试范围距离曝气器50~200 m.
图 4a表示水深80 m(7月)条件下模拟的核心控藻区域.根据从ADP导出的垂向水流速度数据,绘制实测的控藻区域如图 4b所示.对比4a和图 4b可知,在距离曝气器50~200 m的测试范围内,模拟的核心控藻区域和实测区域的形状和位置基本类似,说明模拟的垂向速度与实测速度吻合较好.
 |
| 图 4 核心控藻区域对比(H=80 m) Fig. 4 Comparison of core zone of algae inhibition(H=80 m) |
图 5表明,随着扬水曝气器的运行,表层藻类会随上、下层之间的循环水流而被输送至中、下部水体,表层混合比底部混合稍快,远处藻类比近处藻类混合稍慢.当曝气器运行至第14 d,水体藻类接近完全混合,表层藻类体积分率由峰值0.005陡降至0.001左右,至24 d后基本稳定在0.0006左右,底层藻类体积分率则由0逐渐上升至0.0006左右.使用有机玻璃自动取样器从水深30 m、80 m处取得水样,进而测得藻数量最后计算得出藻类体积分率,即实测值(Cong et al., 2011b).对比实测值可知,藻类浓度的预测比较理想,且预测误差随运行时间的延长逐渐降低,到第24 d降至10%左右.
 |
| 图 5 藻类体积分率的模拟值与实测值对比 Fig. 5 Comparison of simulated and measured algae concentration |
采用FLUENT模拟的扬水曝气器外围流场以进口顺时针环流和其他区域逆时针环流为特征(图 6).当扬水曝气器运行时,来自底部的低温水体从入口进入水温较高的表层水体后,利用其自身动能水平向右运动,同时因其密度低于表层水体而向下运动.在入流水体向下运动过程中,会因所受的浮阻力影响而改变运动方向并在入口水流卷吸作用下向上运动,在进口附近形成局部顺时针环流.而在扬水曝气右侧水体内则形成一个大的整体逆时针环流,此逆时针环流的中心基本位于水域中央.
 |
| 图 6 不同水深条件下扬水曝气器外围流场(a.H=50 m,b. H=80 m, c.H=110 m) Fig. 6 Flow outside the water-lifting aerator under different water depths(a.H=50 m,b. H=80 m, c.H=110 m) |
图 6还表明,当水深从50 m逐渐增加到110 m时,进口顺时针环流的范围和强度随水深条件的变化不明显.这是由于进口环流主要受温度梯度的影响,而不同水深条件下水深30 m范围内的温度梯度相同,入口低温水体向下所受到的浮阻力也相同.随着水深的增加,外围流场的垂向速度逐渐增加,主要是由于水体宽度一定的条件下,当水体深度增加时,水体的宽深比减小,壁面效应和水流断面结构导致的二次流的影响作用逐渐增强.同时,外围流场中的速度随扬水曝气器距离的增加而减小,这是因为外围水体的输入能量一定,进口水流在向右运动的过程中会受到水流阻力,导致其潜行动能逐渐减小.
3.3 水深对扬水曝气控藻区域的影响图 7为扬水曝气器外围水体的垂向速度等值线分布,其中,彩色区域被称为核心控藻区域.进入此区域的藻类会随水流下潜至低温、弱光、低溶解氧含量的中下部水体,藻类的生长会因不利环境而明显受抑.
 |
| 图 7 不同水深条件下的核心控藻区域(a.H=50 m,b.H=65 m,c.H=80 m,d. H=95 m,e. H=110 m) Fig. 7 Core zone of algae inhibition under different water depths(a.H=50 m,b.H=65 m,c.H=80 m,d. H=95 m,e. H=110 m) |
由图 7可见,当水深30 m内温度梯度均为0.73 ℃ · m-1时,虽然水深条件不相同,但扬水曝气核心控藻区域的总体形状大致相同.这是因为核心控藻区域直接取决于扬水曝气器外围流场,而扬水曝气器外围流场又主要取决于温度梯度.水深主要影响核心控藻区域的大小,当水深从50 m逐渐增加到110 m时,核心控藻区域的百分比约从12.5%增加到30.6%.
核心控藻区域的半径却随水深的增加而扩大.当水深从50 m逐渐增加到110 m时,核心控制区域的半径约从60 m增大到175 m.主要原因是在水体宽度不足以限制水流发展的条件下,当水体深度在一定范围内增加时,水深将成为水流发展的重要影响因素.在入流速度一定的情况下,当入流水体从扬水曝气器出口进入其外围水体后,入口水流在水体中的运动类似于平抛运动,模拟区域入口与出口的垂直距离将影响区域内水平射流的发展范围.随着扬水曝气器外围水体水深的增加,水体出口的影响逐渐变弱,入口水流向右的最大运动距离相应逐渐增加,表现为核心控藻区域的逐渐增大.
3.4 水深对藻类迁移的影响图 8为水深80 m条件下藻类被动迁移的迹线,垂直方向上,该迹线为水流速率与藻类上浮速率的差值,横、纵坐标分别表示离曝气器的距离和水深,3个代表点均距扬水曝气250 m,水深分别为3、20、40 m.藻类在水库中的循环可分为表层流动阶段、下降阶段、水底流动阶段和上升阶段.水深3 m处藻类首先水平向左迁移至进口环流区,然后下潜至中下部水体;之后不是沿库底一直潜行,而是悬浮并停留在水体左侧底部.水深3 m处藻类在藻类生长补偿点以下(即3 m以下)循环一周约需60 h,而铜绿微囊藻黑暗限气环境下的极限耐受时间为48 h(郭莉莎等,2012),说明该处藻类的生长会被完全抑制.水深20 m、40 m处藻类迁移轨迹与水深3 m处藻类大致相同,均在藻类生长补偿点以下.
 |
| 图 8 水深80 m条件下藻类被动迁移迹线 Fig. 8 Pathlines of algae transported passively under water depth of 80 m |
由图 8可见,在扬水曝气器运行时,藻类会随水流被动迁移,主要分为表层水平、垂直向下、底部水平和垂直向上运动4个阶段.藻类在生长补偿点以下的停留时间随水深增加而延长,当水深分别为50、65、80、95、110 m时,该停留时间分别为40、60、125、131、132 h.在各水深条件下,水体输入动能相同,单位水体获得的能量随水深增加而减小,导致水体平均速度随水深增加而减小,藻类迁移的速度随之减小,藻类在生长补偿点以下的运动时间也随水深增加而延长.
图 8说明,当水面下30 m内温度梯度均为0.73 ℃ · m-1、水深在50~110 m时,扬水曝气核心控藻区域的半径为60~175 m.图 5则说明,非核心除藻区域的藻类会被动迁移至核心区,再下潜至藻类的生长补偿点之下,从而藻类的生长受到完全抑制,亦即有效的控藻半径为300 m.理论而言,藻类在水体中的迁移包括移流传输、紊流扩散、反应和源(汇)项,本研究中,水域相对封闭,仅考虑对藻类迁移影响较大的扩散和移流项.图 2所示水体流速较小,紊动较弱,藻类的迁移主要受扬水曝气产生的循环水流的传输.模拟水域内藻类完全混合的时间随水深的增加而延长(图 9).
 |
| 图 9 不同水深条件下藻类完全混合的时间 Fig. 9 Time required for complete mixing of algae under different water depths |
1)水深30 m以内变温层水温结构相同的情况下,扬水曝气器外围稳定流场以进口顺时针环流和其他区域逆时针环流共存为特征;随着水深的增加,外围流场垂向速度逐渐增加;外围流场速度值沿径向递减.
2)在水深50~110 m条件下,扬水曝气器外围稳定核心控藻区域的形状基本不受水深条件的影响,核心控藻区域的面积和半径随着水深的增加而增大.当水深从50 m逐渐增加到110 m时,核心控藻区域的半径约从60 m增大到175 m,即从水深的1.2倍增加到1.6倍;核心控藻区域面积百分比约从12.5%增大到30.6%.
3)非核心控藻区域内的藻类会随环流被动迁移至核心区域,并最终悬浮并滞留在扬水曝气器底部附近;藻类在生长补偿点以下的停留时间随水深增加而延长,当水深从50 m增加到110 m时,该停留时间从40 h增加到132 h,藻类完全混合的时间约从16 d增加到30 d.
| [1] | 丛海兵,黄廷林,缪晶广,等. 2007. 扬水曝气器的水质改善功能及提水、充氧性能研究[J].环境工程学报,1(1): 7-13 |
| [2] | 丛海兵,黄廷林,周真明. 2009.于桥水库铜绿微囊藻上浮运动规律及其控制[J].给水排水, 35(11): 140-145 |
| [3] | Cong H B, Huang T L, Chai B B, et al. 2009. A new mixing-oxygenating technology for water quality improvement of urban water source and its implication in a reservoir [J]. Renewable Energy, 34(9): 2054-2060 |
| [4] | Cong H B, Huang T L, Chai B B. 2011a. Water-circulating aerator: optimizing structure and predicting water flow rate and oxygen transfer [J]. Journal of Hydraulic Engineering, 137(6): 659-667 |
| [5] | Cong H B, Huang T L, Chai B B. 2011b. Research on applying a water-lifting aerator to inhibit the growth of algae in a source-water reservoir[J]. International Journal of Environment and Pollution, 45(1): 166-175 |
| [6] | 崔福义,马华. 2011.水源水中藻的危害与饮用水除藻技术[J].给水排水, 37(6): 1, 103 |
| [7] | 郭莉莎,章军,吴娟,等.2012.黑暗限气条件下铜绿微囊藻细胞死亡的形态结构和生理生化变化[J].微生物学报,52(2): 228-235 |
| [8] | 韩晓婷,常青,毛玉红,等.2013. Taylor-Couette流场数值模拟及絮凝效果研究[J].环境科学学报,33(6): 1637-1641 |
| [9] | Hodges B R. 1999. Heat budget and thermodynamics at a free surface . ED 1300 BH. Centre for Water Research, Perth: The University of Western Australia |
| [10] | 黄廷林,丛海兵,柴蓓蓓. 2009.饮用水水源水质污染控制[M].北京: 中国建筑工业出版社 |
| [11] | Hudnell H K, Jones C, Labisi B, et al. 2010. Freshwater harmful algal bloom (FHAB) suppression with solar powered circulation (SPC) [J]. Harmful Algae, 9(2): 208-217 |
| [12] | Li J, Zhang J S, Miao J, et al. 2006. Application of computational fluid dynamics (CFD) to ozone contactor optimization[J]. Water Science and Technology: Water Supply, 6(4): 9-16 |
| [13] | 黎雷, 高乃云, 殷娣娣, 等. 2008. 控制饮用水原水中藻类、藻毒素的水厂处理工艺[J].中国给水排水, 24(6): 20-24 |
| [14] | 马越, 黄廷林,丛海兵, 等. 2012.扬水曝气技术在河道型深水水库水质原位修复中的应用[J]. 给水排水, 38(4): 7-13 |
| [15] | 孙昕, 黄廷林. 2013. 湖泊水库水体污染控制[M]. 武汉: 湖北科学技术出版社 |
| [16] | 王福军. 2004. 计算流体动力学分析-CFD软件原理与应用[M]. 北京: 清华大学出版社 |
| [17] | 许保玖. 2000. 给水处理理论[M]. 北京: 中国建筑工业出版社 |