 2014
2014



2 广州甘蔗糖业研究所 广东省甘蔗改良与生物炼制重点实验室, 广东广州 510316
2 Guangzhou Sugarcane Industry Research Institute/Guangdong Key Lab of Sugarcane Improvement & Biorefinery, Guangzhou 510316
甘蔗是生长在亚热带和热带地区的一种经济作 物,是我国最主要的糖料作物。我国盐碱地面积达 9913 万hm2[1],而且盐渍化趋势日益加剧,甘蔗种 植区特别是广东广西沿海的耕地盐渍化更加严重。 土壤中的钠离子含量过高会对作物产生不良影响, 盐胁迫使细胞膜的组分和透性等发生一系列变化, 导致细胞膜的功能受损进而使细胞的代谢及生理功 能受到破坏[2, 3]。由于全球气候变暖、人口增长、 工业污染和化肥使用不当等因素,土壤盐渍化日趋 严重,已成为阻碍作物生长发育及高产优质的主要 因素之一[4, 5],盐胁迫下非盐生植物体内Na+积累的 同时,常伴随着K+含量的下降[6]。钾是植物所必需 的3 大营养元素之一,是植物体内多种酶的活化 剂[7],钾对作物的盐胁迫具有一定的缓解作用,在 其他作物如玉米[8]、长春花[9]、小麦[10]等多种作物 中都有报道,但在甘蔗还未见报道。甘蔗是一种中 度耐盐作物,一般土壤含盐量在0.3%以下才能正 常生长[11]。本研究利用水培方法探讨了钾对盐胁迫 下甘蔗苗期生长的影响,有助于揭示钾对甘蔗盐胁 迫缓解的机理,并为盐渍地甘蔗生产提供科学依 据。
1 材料与方法 1.1 材料2012 年8 月10 日在广州甘蔗糖业研究所温室 内,用ROC22 单芽蔗茎置于育苗盘,用河沙覆盖 浇水待其出苗,于9 月10 日移栽到水培,用霍格 兰营养液培养2 周开始处理。
1.2 试验处理根据前期预实验结果设置盐胁迫浓度为质量分 数0.5%的氯化钠。营养液中钾离子浓度设置4 个 浓度,分别为K1 (0 mmol/L)、K2 (0.5 mmol/L)、K3 (3 mmol/L)、K4 (6 mmol/L),其中K1 为无钾对照。 每个处理重复4 次,每周更换营养液1 次。各处理 营养液大量元素配方见表1,微量元素统一为:FeSO4 13.9 mg/L,Na2EDTA 18.61 mg/L,MnSO4·4H2O 2.13 mg/L,H3BO3 2.86 mg/L,CuSO4·5H2O 0.08 mg/L, ZnSO4·7H2O 0.22 mg/L,(NH4)6Mo7O24·4H2O 0.02 mg/L。培养8 周后收获取样调查,分析各项指标。
| 表1 不同处理营养液大量元素配方(mg/L) |
植株株高和根系长度用直尺量取,根冠比即为 根系长度和株高长度的比值,植株钠、钾含量用硫 酸双氧水消煮-火焰光度计测定,地上部、地下部 干鲜重,根冠比,叶片SPAD 值用SPAD502 仪器 测定,保水力测定方法:取叶片称其鲜重,在恒温 20℃条件下,经24、48 h 后再对叶片称重,用2 次重量之差与第1 次重量之比(即失水量)表示[10]。
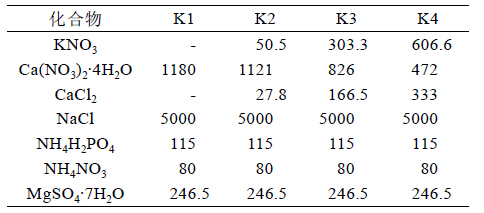
2 结果与分析 2.1 不同处理对甘蔗生物量的影响根据对甘蔗植株体不同部位的干鲜重统计,在 盐胁迫条件下,随着溶液中钾浓度的提高,植株体 叶片、茎、根系的干鲜重也随之增加,当溶液中钾 浓度上升到0.5 mmol/L(K2)时,植株体的叶片和茎 的生物量已显著高于对照(K1),而当钾浓度上升到 6 mmol/L 时,其植株生物量又显著高于其他各个 处理,说明在盐胁迫下,提供充足的钾能促进植株 的生长(见表2)。
| 表2 不同处理下甘蔗鲜重和干重 |
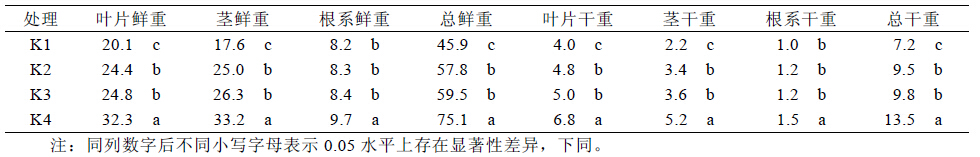
根系是植物吸收养分的重要器官。盐胁迫下, 在营养液中无钾(K1)和低钾(K2)的情况下根系长度 和株高均显著低于K3 和K4,说明在盐胁迫下增 加溶液中钾的浓度有利于根系生长,同时促进地上 部的生长。作物在苗期通常有较大的根冠比,根系 生长良好有利于地上部的生长。钾肥有调节碳水化 合物转化和运输的作用,可促进光合产物向根和贮 藏器官的转移,通常能增加根冠比。表3 的结果表 明,当溶液中钾浓度增加到3 mmol/L(K3)时,植株 根冠比已显著大于K2 和K1 时的根冠比。
| 表3 不同处理下甘蔗根冠比 |
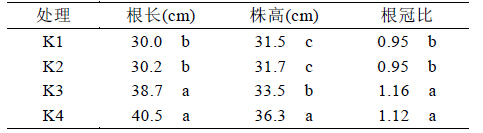
在盐胁迫下植株根系吸收水分难度增大,相应 地植株含水率会降低。通过测量植物叶片失水量可 知植物保水能力的大小。本试验甘蔗叶片的保水能 力用失水率来表示,失水率越大,其保水能力越差。 从表4 可以看出,甘蔗叶片在20℃恒温下的失水 很快,24 h 失水率达45%以上,48 h 失水率更达64%以上。不同处理之间存在显著差异,在K1 无钾条 件下,植株失水率要显著高于其余3 个有钾处理, 说明在盐胁迫下,给植株提供钾能降低叶片失水 率,提高植株保水能力。SPAD 值是反映植物叶片 叶绿素相对含量的指标,其值越大,植物叶片叶绿 素含量越高,叶绿素是植物进行光合作用的基础条 件之一。由表4 可知,在无钾条件下甘蔗叶片的 SPAD 值显著低于其他3 个处理,说明在盐胁迫下 提供钾有利于提高甘蔗叶片叶绿素含量。
| 表4 不同处理下甘蔗离体叶片失水率 |
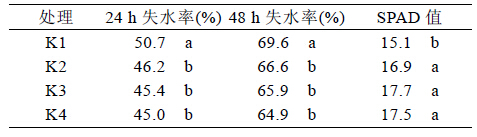
从表5 的结果可以看出在盐胁迫下,随着钾水 平的提高,甘蔗叶片中K+含量也随之增加,而Na+ 含量则随之降低,说明两者之间存在竞争和互相抑 制。在无钾条件下,甘蔗叶片中Na+的含量要高于 K+,而在加钾的3 个处理下甘蔗叶片K+含量要高 于Na+含量。研究表明植物体内K+/Na+值越高,其 耐盐性越好。由表5 的结果可知,随着溶液中钾浓 度的提高,K+/Na+值也随之升高,在无钾处理下, 其值小于1,加钾处理下其值均大于1。说明在盐 胁迫下,提供充足的钾可以提高甘蔗叶片的钾钠比 值,增强植株耐盐性,减少Na+的吸收。
| 表5 不同处理下甘蔗叶片钾含量、钠含量和钾钠比 |

盐胁迫条件对作物生长会产生不利影响,盐分 对植物生长的影响主要与渗透胁迫和离子毒害有 关,盐害最早的症状之一是植株茎部维管束系统钾 迁移的中断[12]。本研究在不同供钾条件下,甘蔗鲜 重增长和干重增长有显著差异,在盐胁迫条件下供 钾的生长效应高于对照,这可能是由于供钾能更好 地使甘蔗适应盐诱发的水分胁迫和离子毒害环境。 植物受到盐胁迫后Na+含量升高,K+/Na+比迅速下 降[13],本试验表明,随着根系环境钾浓度增加,植 株对钾的吸收也相应增加,从而获得较高的K+/Na+ 比值,提升植株的耐盐性。叶片是植物进行光合作 用的重要器官,叶绿素和水分是光合作用的重要因 子。林希昊等研究表明盐胁迫下甘蔗叶片质膜透性 增加,叶绿素降低[14]。本研究在盐胁迫下,甘蔗叶 片的失水率较大,而供钾后叶片失水率显著降低, 即叶片保水能力增加,同时叶片SPAD 值得到显著 提升,即叶绿素含量得到提高,因此可以说明,甘 蔗盐胁迫下供钾能通过提高叶片保水能力和叶绿素 含量从而增进光合作用,进而对甘蔗的盐胁迫起到 缓解作用。对于钾提升甘蔗耐盐性的深层次机理以 及甘蔗生育后期的表现则还需进一步的试验研究。
本研究结果表明,在盐胁迫下供钾能促进苗期 甘蔗生长,甘蔗生物量、根长、株高和根冠比在供 钾情况下得到显著提高,同时提高甘蔗叶片的保水 力、SPAD 值和钾钠比,从而提升甘蔗耐盐性。
| [1] | 叶铭. 科学家发明治理盐碱地新方法[N]. 人民日报, 2000-02-10(4).( 1) 1) |
| [2] | CHOWDHURY M A M, MOSEKI B, BOWLING D J F. A method for screening rice plants for salt tolerance [J]. Plant and Soil, 1995, 171: 317-322(1) |
| [3] | KATSUHARA M, KUCHITSU K, TAKESHIGE K, et al. Salt Stress-Induced Cytoplasmic Acidification and Vacuolar Alkalization in Nitellopsis obtusa Cells: In VivoP-Nuclear Magnetic Resonance Study [J]. Plant Physiol, 1989, 90(3): 1102-1107(1) |
| [4] | 耿玉珂, 周宜君, 丁宁, 等. 植物耐盐突变体筛选与 耐盐转基因研究[J]. 中央民族大学学报:自然科学版, 2009, 18(4):10-17(1) |
| [5] | CHRISTOPHE B G, TOMADER E, JAMAL A, et al. Selection of callus cultures of sugarcane (Saccharum sp.) tolerant to NaCl and their response to salt stress [J]. Plant Cell Tiss Organ Cult, 2006, 87:9-16.(1) |
| [6] | NAKAMURA Y, TANAKA K, Ohta E. Protective effect of external Ca2+ one longation and intracellular concentration of K+ in intactmung bean roots under high NaCl stress [J]. Plant and Cell Physiology, 1990, 31: 815-821.(1) |
| [7] | MAATHUIS F J. Physiological functions of mineral macronutrients [J]. Curr Opin Plant Biol, 2009, 12(3): 250-258(1) |
| [8] | 赵洪彬, 刘厚忠, 张道园, 等. 钾对盐胁迫不同基因 型玉米幼苗光合特性的影响[J]. 吉林农业, 2011(5): 141-144(1) |
| [9] | 宋姗姗, 隆小华, 刘玲, 等. 钠钾比对盐胁迫下盛花 期长春花离子分布和光合作用的影响[J]. 土壤学报, 2011, 48(4):884-887(1) |
| [10] | 吕金岭, 董永. 盐逆境胁迫下施钾对降低小麦盐害的 生理效应研究[J]. 江西农业学报, 2007, 19(3):39-40(2) |
| [11] | 何启钧, 林彦栓, 陈春花, 等. 甘蔗的抗盐育种技术-盐分胁迫下甘蔗无性系的产量和品质性状表现[J]. 福 建农学院学报:自然科学版, 1993, 22(4):398-403(1) |
| [12] | MARCUMK B. Salinity tolerance mechanisms of grasses in the subfamily chloridoideae [J]. Crop Science, 1999(39):1153-1160.(1) |
| [13] | HASSAN I A. Interactive effects of salinity and ozone pollution on photosynthesis, stomatal conductance, growth, and assimilate partitioning of wheat (Triticuma estivum L.) [J]. Photosynthetica, 2004, 42(1):111-116(1) |
| [14] | 林希昊, 苏俊波, 刘洋, 等. 盐胁迫对甘蔗苗期生理 的影响[J]. 西南农业学报, 2011, 24(3):911-914(1) |