 2011
2011



甘蔗是我国主要的糖料作物,在我国食糖供应 中具有举足轻重的地位和作用,甘蔗糖占我国食糖 供应的90%左右[1]。由于甘蔗属于无性繁殖作物, 生产上一般用蔗梢作种,连续种植多年后种苗容易 感染或者积累病毒,造成产量、糖分大幅降低[2,3]。 生产健康种苗是一种有效的防治甘蔗病害、提高产 量和糖分、延长宿根年限的方法。已有研究结果表 明,利用甘蔗茎尖培养与热水处理等技术,可以有 效去除蔗株携带的花叶病毒、宿根矮化病病菌等, 减少因病害发生造成的甘蔗减产、糖分降低等[4,5]。 目前,世界上蔗糖主要生产国巴西、澳大利亚、古 巴等都建立了甘蔗健康种苗工厂化生产技术体系[6]。
近年来,我国各甘蔗育种科研单位都非常重视 健康种苗技术研究[7,8],并且建立了健康种苗示范 和应用基地。但是,由于甘蔗遗传背景复杂[9],品 种间遗传差异较大,在组织培养过程中不同品种间 对激素浓度的要求差异较大,不同品种在同一种培 养基上的繁殖系数可以相差几倍。繁殖过程中如果 繁殖效率低,不仅降低生产效率和增加种苗生产成 本,而且会严重降低种苗质量,对田间生产造成不 良影响。因此,为了促进甘蔗健康种苗在生产上的 应用,需要对生产上的主栽品种进行培养基优化实 验,筛选合适的激素浓度和组合,提高健康种苗繁 殖效率和质量。
正交试验设计应用于优化培养条件的生物学研 究,主要优点是能够在多试验条件中通过较少试验 方案找出最优方案或最佳培养条件。薛志平等采用 正交设计法[10],考察不同因素对甘蔗愈伤组织分化 的影响。本研究通过正交试验分析不同激素浓度对 甘蔗丛生苗繁殖效率的影响,优化培养条件,提高 健康种苗生产效率和质量。
实验材料为ROC22、粤糖00-236。
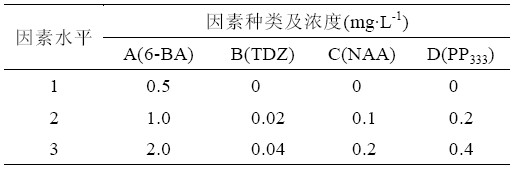
培养基采用MS 培养基,根据6-苄氨基嘌呤 (6-BA)、噻苯隆(TDZ)、萘乙酸(NAA)、多效唑(P333)4 种物质间的浓度设立正交组合,采用L9(34)正交试 验设计,配比4 因素3 水平(表1)。选取高度为株 高2~4 cm,生长势好,生长状态一致的同一批次 茎尖分化出的丛芽苗,分别接入到各个培养基中, 每种培养基接种18 瓶,每瓶接3 丛。培养条件为 每天光照培养16 h,温度28℃,光强2000 lx。10 天后统计每瓶苗的分化芽数。
表1 正交试验因素水平表L9(34)
| 表1 正交试验因素水平表L9(34) |
计算各个试验处理的平均值,用方差分析比较 各个处理间的差异显著性,统计分析采用SPSS 11.0 软件[11]。
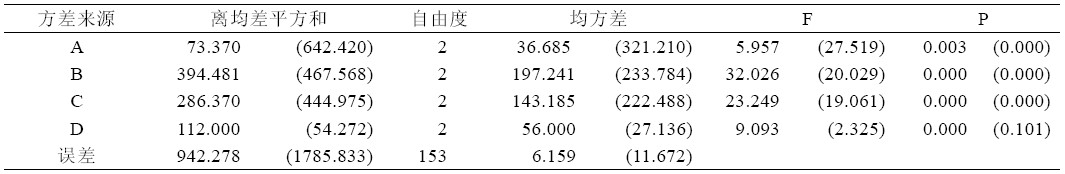
从试验结果的方差分析可以看出(表2),6-BA、 TDZ、NAA、P333 这4 个因素对ROC22 丛芽苗分 化的影响均存在极显著差异(P<0.01),但其影响程 度不同,对ROC22 丛芽苗分化影响的主次因素为 TDZ>NAA>P333>6-BA。而这4 个因素对粤糖 00-236 丛芽苗增殖的影响与ROC22 也有明显不 同,P333 对粤糖00-236 丛芽苗分化的影响不存在 显著差异(P>0.05),而其他3 个因素6-BA、TDZ、 NAA 则表现极显著差异(P<0.01),从方差分析表 可以看出,各因素对粤糖00-236 丛芽苗分化的影 响主次顺序为6-BA>TDZ>NAA>P333。
表2 6-BA、TDZ、NAA、P333 4 个因素方差分析表
| 表2 6-BA、TDZ、NAA、P333 4 个因素方差分析表 |
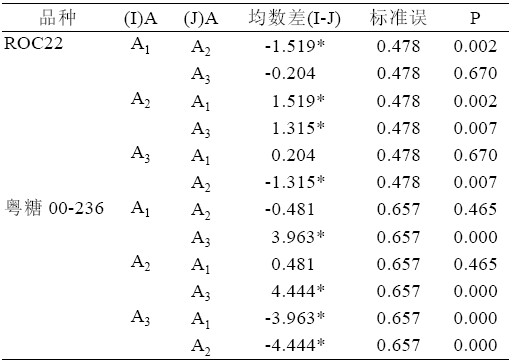
综合表3、表4 可以得出:ROC22 在6-BA 浓 度为A2 时,均数最大5.889,且A2 与A1、A3 之间 存在显著性差异(P<0.05),A1 与A3 之间不存在 显著性差异(P>0.05)。粤糖00-236 也是在6-BA 浓度为A2 时新芽均数最大12.833,与浓度为A1 时 的均数仅相差0.481,且2 者无显著差异(P>0.05), 但与6-BA 浓度A3 相比,2 者存在极显著差异(P< 0.01)。
表3 6-BA 单因素统计量表
| 表3 6-BA 单因素统计量表 |
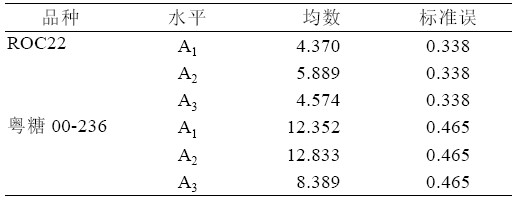
表4 6-BA 浓度水平配对比较表
| 表4 6-BA 浓度水平配对比较表 |
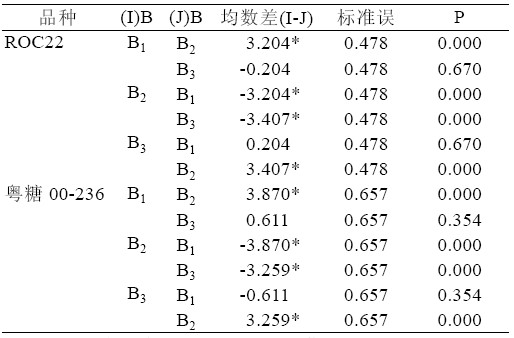
综合表5、表6,可以得出:ROC22 在TDZ 浓度B3 时,丛芽苗分化出新芽的均数最大为6.148, 与TDZ 浓度B1 均数5.944 仅差0.204,2 者无显著 差异(P>0.05)。粤糖00-236 在TDZ 浓度B1 时,新 芽均数已达到12.685,与浓度为B3 时均数12.074 相比,2 者无显著差异(P>0.05)。
表5 TDZ 单因素统计量表
| 表5 TDZ 单因素统计量表 |
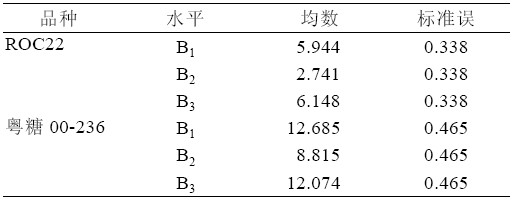
表6 TDZ 浓度水平配对比较表
| 表6 TDZ 浓度水平配对比较表 |
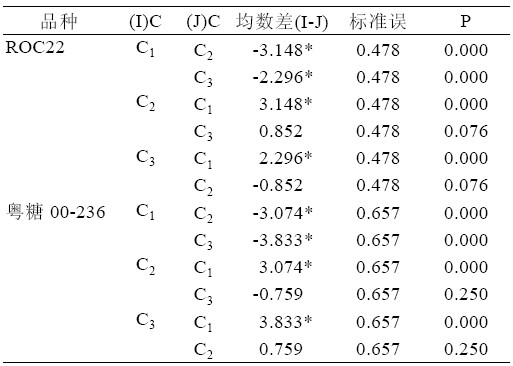
综合表7、表8,可以得出:ROC22 在NAA 浓度C2 时,丛芽苗分化新芽的均数达最大,与NAA 浓度为C3 新芽均数相比无显著性差异(P>0.05), 与未添加NAA 相比存在显著性差异。粤糖00-236 在NAA 浓度为C3 时新芽均数最大为12.722,与浓 度C2 无显著差异(P>0.05)。但是,C3、C2 与C1 之 间存在极显著差异(P>0.01)。
表7 NAA 单因素统计量表
| 表7 NAA 单因素统计量表 |
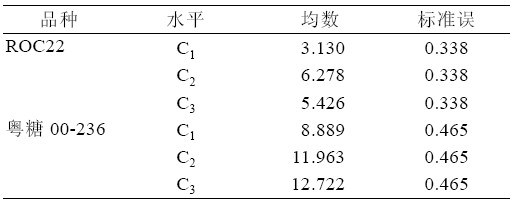
表8 NAA 浓度水平配对比较表
| 表8 NAA 浓度水平配对比较表 |
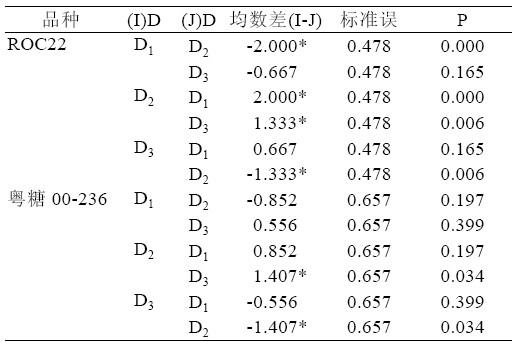
综合表9、表10,可以得出ROC22 在P333 浓 度D2 时均数最大为6.065,且D2 与D1、D3 之间存 在极显著差异(P<0.01)。可见,ROC22 在P333 浓 度为D2 时,丛芽苗分化效果较好。粤糖00-236 也 是在P333 浓度为D2 时,新芽均数最大为11.944, D2 与D3 之间存在显著性的差异(P<0.05),但是D1 与D2、D3 之间不存在显著性的差异(P>0.05)。从 粤糖00-236 在D3 时的新芽数小于D1 时的新芽数, 说明在使用P333 时要筛选合适的使用量,P333 使 用过多对新芽生长存在不利影响。
表9 P333 单因素统计量表
| 表9 P333 单因素统计量表 |
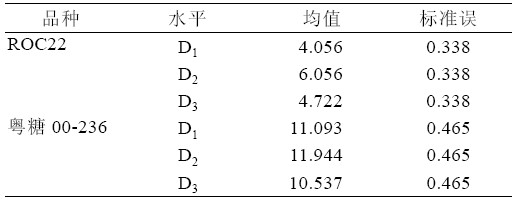
表10 P333 浓度水平配对比较表
| 表10 P333 浓度水平配对比较表 |
甘蔗不同基因型组织培养特性有显著差异,基 因型影响甘蔗愈伤组织诱导,增殖和分化[12,13]。本 研究以ROC22 与粤糖00-236 为供试材料,取茎尖 分化出的丛芽苗,在相同的培养条件下,使用相同 植物生长调节剂处理,ROC22 丛芽苗分化新芽数 明显低于粤糖00-236。可见,甘蔗基因型的自身特 性决定了该品种的丛芽苗分化能力,甘蔗组织培养 特性受遗传特性决定[14]。
外源激素的刺激使多酚氧化酶活性提高[15],若 降低外源激素含量可大大降低酚害影响[16],提高丛 芽苗诱导率, 而不同激素混合使用有利于降低分化 苗的变异率[8]。本研究在甘蔗丛芽苗分化增殖的阶 段,选用了2 种细胞分裂素类物质6-BA,TDZ。 粤糖00-236 的丛芽苗的分化在6-BA 浓度在0.5 mg/L 与1.0 mg/L 2 个水平,差异不显著。从降低 酚害影响,提高丛芽苗分化能力考虑,粤糖00-236 在繁殖阶段6-BA 浓度取0.5 mg/L 已达到较好繁殖 效果。岑秀芬等研究在甘蔗愈伤组织分化阶段[13], 使用浓度 0.04 mg/L 的TDZ,并与其他激素合理配 组,可提高愈伤组织分化效果。而本研究表明在甘 蔗丛芽苗繁殖阶段若培养基中使用6-BA,再添加 TDZ 对丛芽苗增殖影响并不显著。如添加浓度不 适,反而会影响丛芽苗的增殖效果。可见,在甘蔗 组织培养中,应根据其不同时期生长需要,选取适 宜的激素组合。
在细胞分裂素与生长素存在适当比例时,芽的 诱导能高效进行。建立适宜的激素比例,尤其是生 长素/分裂素之间的最佳组合,是提高甘蔗品种繁 殖效率的最重要因素之一。研究中发现,ROC22 和粤糖00-236 新芽数最大时培养基中6-BA 的浓度 相同,但是NAA 和TDZ 的浓度都不同,由此可见 不同品种对生长素/分裂素的比例要求不一致,在 健康种苗繁殖过程中要针对某个品种筛选适宜的生 长素/分裂素的比例。
早在上世纪90 年代,植物生长延缓剂P333 已 应用于植物组织培养领域,并取得良好效果[17]。多 效唑的添加使得甘蔗丛芽苗健壮生长,植株叶色浓 绿,叶片平展,芽苗质量提高,有壮苗作用,并且 可保持芽苗良好的生长状态[18]。P333 对ROC22 不 仅有壮苗作用,在P333 浓度达到0.2 mg/L 时,丛 芽苗的分化新芽数也有增加。而粤糖00-236 对P333 的反映与ROC22 明显不同,植株生长矮壮,但丛 芽苗的分化并未受到显著影响。通过添加适量P333 可以明显降低株高,减少继代时的工作量。在甘蔗 健康种苗生产中,可根据植物组织培养生产计划, 适当添加P333,调节丛芽苗生长速度,调控丛芽 苗继代时间。
| [1] | 李奇伟.现代甘蔗改良技术[M]. 广州:华南理工大学出版社,2000. ( 1) 1)
|
| [2] | Lee.T.S.G.Micropagation of sugarcane[J]. Plant Cell,Tissue Organ Culture,1987(10):45-47. (1)
|
| [3] | 游建华,何为中,曾慧,等.谈脱毒健康种苗在广西甘蔗生产的应用及效益展望[J]. 甘蔗糖业,2001(1):13-17. (1)
|
| [4] | 杨本鹏,张树珍,杨学,等.甘蔗健康种苗培育体系的建立[J]. 热带作物学报,2006(27):74-77. (1)
|
| [5] | 陈仲华,沈万宽.利用热水处理结合心叶愈伤组织培养脱除甘蔗宿根矮化病菌研究[J]. 甘蔗糖业,2007(5):16-20. (1)
|
| [6] | 王伦旺,何红,谭裕模,等.赴澳大利亚甘蔗糖业科技学习考察报告[J]. 中国糖料,2009(2):75-80. (1)
|
| [7] | 许莉萍,傅华英,潘大仁.甘蔗腋芽快速繁殖培养基及激素配方的筛选[J]. 福建农业大学学报,2001,29(4):401-404. (1)
|
| [8] | 陈彪,梁钾贤,陈伟栋,等.甘蔗组织培养配方中不同激素效应的研究[J]. 华南农业大学学报,2000,21(1):60-62. (2)
|
| [9] | 张华,林彦铨.甘蔗细胞与分子遗传学研究进展[J]. 甘蔗,2001(3):1-6. (1)
|
| [10] | 薛志平,林美娟,陈平华,等.正交试验设计在优化甘蔗再生体系中的应用[J]. 江西农业大学学报,2005(3):389-393. (1)
|
| [11] | 夏传涛,袁秉祥.无空列正交试验的设计及SPSS软件的数据处理[J]. 数理医药学杂志,2006,19(1):91-92. (1)
|
| [12] | 梁计南,谭中文,谭志勇,等.甘蔗不同基因型组织培养特性的研究[J]. 华南农业大学学报(自然科学版),2002,10(4):37-40. (1)
|
| [13] | 岑秀芬,杨燕妮,韦鹏霄,等.激素因子对甘蔗愈伤组织诱导增殖与分化的效应[J]. 广西农业科学,2010,41(9):889-892. (2)
|
| [14] | 谭志勇,谭中文,张志胜.不同基因型甘蔗组织培养特性的研究[J]. 广东农业科学,2005,1:34-36. (1)
|
| [15] | 秦廷豪,邹宗兰,吴才文.浅析甘蔗组织培养中的酚害[J]. 甘蔗,1997(2):12-14. (1)
|
| [16] | 贤武,王伦旺,王天算,等.甘蔗茎尖脱毒培养研究初报[J]. 广西蔗糖,2000(4):3-5. (1)
|
| [17] | 高凤菊,汤忠梅,王晓理,等.PP333在植物组织培养上的应用进展[J]. 农业与技术,2002(2):66-69. (1)
|
| [18] | 李松,何新民.烯效唑在甘蔗组织培养中的应用研究[J]. 甘蔗糖业,2001(6):19-22. (1)
|