
Cancer-associated fibroblasts mediate migration of myeloid-derived suppressor cells in pancreatic ductal adenocarcinoma through SDF-1/CXCR4 pathway
-
摘要:
目的 探讨胰腺导管腺癌(PDAC)中肿瘤相关成纤维细胞(CAF)调节CD13高表达的中性粒细胞样髓源性抑制细胞(CD13hi-nMDSC)迁移的机制,为PDAC的免疫治疗提供实验依据和潜在分子靶标。 方法 从5例PDAC患者的胰腺癌组织中分离纯化CAF,用细胞免疫荧光、流式细胞术鉴定CAF表型和纯度,用qPCR、ELISA技术检测CAF中相关细胞因子的表达。用Transwell系统构建CAF条件培养基及髓源性抑制细胞(MDSC)迁移系统,观察MDSC的迁移情况,研究上述细胞因子参与调节MDSC迁移的具体作用机制。 结果 分离的原代CAF表达活化标志物成纤维细胞激活蛋白(FAP)和α- 平滑肌肌动蛋白(α-SMA),对照人包皮成纤维细胞(HFF)不表达FAP和α-SMA。qPCR结果显示CAF中IL-6、单核细胞趋化蛋白1(MCP-1)、基质细胞衍生因子1(SDF-1)mRNA的表达高于HFF;CAF培养上清中IL-6、MCP-1、SDF-1的含量均高于HFF培养上清(均P<0.01),且随培养时间的延长而升高。与HFF条件培养基和普通培养基(RPMI 1640)相比,CAF条件培养基能招募更多的总MDSC和CD13hi-nMDSC(均P<0.01);在培养体系中单独加入SDF-1重组蛋白可以诱导总MDSC和CD13hi-nMDSC的迁移,并且加入SDF-1中和抗体或CXC趋化因子受体4(CXCR4)阻断抗体后能够减少CAF条件培养基诱导的CD13hi-nMDSC迁移(均P<0.01);虽然单独加入MCP-1也可以诱导总MDSC和CD13hi-nMDSC的迁移,但CD13hi-nMDSC迁移的数量明显少于SDF-1实验组;IL-6重组蛋白不能引起总MDSC和CD13hi-nMDSC的迁移。 结论 CAF可通过SDF-1/CXCR4通路介导PDAC中总MDSC及CD13hi-nMDSC的迁移。 Abstract:Objective To explore the mechanism by which cancer-associated fibroblasts (CAFs) regulate CD13-high expression neutrophil-like myeloid-derived suppressor cell (CD13hi-nMDSC) migration in pancreatic ductal adenocarcinoma (PDAC), so as to provide potential molecular targets and experimental evidences for immunotherapy in patients. Methods CAFs were isolated and purified from pancreatic cancer tissues of 5 PDAC patients. The phenotype and purity of CAFs were identified by immunofluorescence and flow cytometry. The expression of related factors in CAF was detected by quantitative polymerase chain reaction (qPCR) and enzyme-linked immunosorbent assay (ELISA). CAF conditioned medium and myeloid-derived suppressor cell (MDSC) migration system were constructed by Transwell to observe the migration of MDSCs and to study the specific mechanisms by which the aforementioned cytokines participate in regulating the migration of MDSCs. Results The isolated primary CAFs expressed activation biomarkers fibroblast activation protein (FAP) and α-smooth muscle actin (α-SMA), while the human foreskin fibroblasts (HFFs) of control cells did not express FAP and α-SMA. qPCR results showed that the mRNA expression levels of interleukin 6 (IL-6), monocyte chemotactic protein 1 (MCP-1), and stromal cell-derived factor 1 (SDF-1) in CAFs were higher than those in HFFs (all P < 0.01). The contents of IL-6, MCP-1, and SDF-1 in the CAF culture supernatant were significantly higher than those in the HFF culture supernatant (all P < 0.01), and the secretion content increased with the prolongation of culture time. Compared with HFF conditioned medium and regular medium (RPMI 1640), CAF conditioned medium could recruit more total MDSCs and CD13hi-nMDSCs (all P < 0.01). The addition of SDF-1 recombinant protein alone in the culture system could induce the migration of total MDSCs and CD13hi-nMDSCs, and the addition of SDF-1 neutralizing antibodies or C-X-C motif chemokine receptor 4 (CXCR4) blocking antibodies could significantly reduce the migration of CD13hi-nMDSCs induced by CAF conditioned medium (all P < 0.01). Although MCP-1 alone could also induce the migration of total MDSCs and CD13hi-nMDSCs, the number of CD13hi-nMDSCs migrating was significantly less than that of the SDF-1 experimental group. The IL-6 recombinant protein did not induce the migration of total MDSCs or CD13hi-nMDSCs. Conclusion CAFs can mediate the migration of total MDSCs and CD13hi-nMDSCs in PDAC through SDF-1/CXCR4 pathway. -
胰腺导管腺癌(pancreatic ductal adenocarcinoma,PDAC)是常见的消化系统恶性肿瘤病种,具有高侵袭性、预后差的特点,5年预期生存率不足10%[1]。近年来,PDAC的发病率以每年0.5%~1% 的速度增长,在中国2022年肿瘤死因中居第6位,在美国肿瘤相关死亡中居第3位,预计未来10年内PDAC的发病率将继续上升,到2030年将成为肿瘤第二大致死病种[2-3]。
PDAC是肿瘤间质最丰富的实体肿瘤之一,间质中存在丰富的肿瘤相关成纤维细胞(cancerassociated fibroblast,CAF)。CAF通过分泌多种细胞因子参与募集免疫抑制细胞,阻止免疫细胞杀伤肿瘤细胞,帮助胰腺癌细胞逃避宿主免疫杀伤[4-5]。髓源性抑制细胞(myeloid-derived suppressor cell,MDSC)是肿瘤间质微环境中最重要的免疫抑制细胞群,其可分泌过氧化物酶、一氧化氮合酶、TGF-β、IL-10和环氧合酶2,发挥免疫抑制作用[6]。有研究表明集落刺激因子1(colonystimulating factor 1,CSF-1)/ 集落刺激因子1受体(colony stimulating factor 1 receptor,CSF-1R)信号通路是肿瘤相关巨噬细胞分化和存活的关键调节因子,CSF-1R抑制剂可以耗竭肿瘤相关巨噬细胞从而发挥抗肿瘤作用,但在实体肿瘤中,单纯使用CSF-1R抑制剂疗效不明显[7]。Kumar等[8]发现CAF能通过促进中性粒细胞样MDSC(neutrophil-like MDSC,nMDSC)在肿瘤中的聚集来抑制CSF-1R抑制剂的抗肿瘤作用。成纤维细胞激活蛋白(fibroblast activation protein,FAP)是一种具有双酶活性的丝氨酸蛋白酶,Liu等[9]推测以FAP为靶点的嵌合抗原受体T细胞(chimericantigen receptor T cell,CAR-T)可以耗竭肿瘤中的CAF,进而抑制MDSC的募集,促进CD8+ T细胞和序贯治疗靶向密封蛋白18.2(claudin 18.2,CLDN18.2)的CAR-T在肿瘤组织中的浸润与存活,最终增强以CLDN18.2为靶点的CAR-T治疗PDAC的疗效。基质细胞衍生因子1(stromal cellderived factor 1,SDF-1)又称CXC基序趋化因子配体(C-X-C motif chemokine ligand,CXCL)12,Sun等[10]发现在乳腺癌中奥拉帕尼可以降低CAF释放SDF-1α,从而抑制MDSC的募集,增加CAR-T的抗肿瘤活性。这些研究成果均表明,CAF参与肿瘤免疫抑制微环境的形成,并与MDSC密切相关。
本课题组前期研究发现,CD13高表达(CD13hi)的MDSC是PDAC中发挥免疫抑制功能的主要细胞亚群,其在胰腺癌患者外周血及肿瘤组织中明显增多[11]。胰腺癌中CAF通过IL-6/STAT3通路促进外周血单个核细胞分化为CD13hi-nMDSC[12],但在PDAC中CAF招募并参与MDSC迁移的机制尚不明确。本研究旨在探讨PDAC癌组织中CAF调节CD13hi-nMDSC迁移的机制,为揭示CAF和MDSC通过重塑胰腺癌微环境促进胰腺癌进展提供理论基础和实验依据。
1 材料和方法
1.1 组织标本、细胞及试剂
5例PDAC组织及3例PDAC患者的脾脏组织均来自于海军军医大学第一附属医院手术切除标本,本研究经海军军医大学第一附属医院伦理委员会审批,所有患者均签署知情同意书。8例患者中男6例、女2例,年龄为(67.37±7.41)岁,术前均未接受放疗及化疗,均行胰腺癌根治性切除手术治疗,术后病理结果均为PDAC。人包皮成纤维细胞(human foreskinfibroblast,HFF)及成人皮肤成纤维细胞(humandermal fibroblast-adult,HDF-a)由上海富衡生物科技有限公司提供。
FBS、RPMI 1640培养基购自美国Gibco公司;透明质酸酶、胰蛋白酶、胶原蛋白酶及DNA酶Ⅰ购自美国Sigma公司;RNA提取试剂盒购自上海飞捷生物技术有限公司;qPCR仪购自日本TaKaRa公司,α- 平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)抗体、波形蛋白(vimentin)抗体购自武汉博士德生物工程有限公司;PerCP-Cy5.5标记的抗人CD33抗体、Pacific blue标记的抗人类白细胞抗原DR(human leukocyte antigen-DR,HLA-DR)抗体、PE-Cy7标记的抗人CD15抗体、FITC标记的抗人CD11b抗体、PE标记的抗人CD13抗体、APC标记的抗人CD14抗体、PE标记的抗人FAP抗体、人IL-6 ELISA检测试剂盒、人单核细胞趋化蛋白1(monocyte chemotactic protein 1,MCP-1)ELISA检测试剂盒购自美国BioLegend公司;人SDF-1 ELISA检测试剂盒购自杭州联科生物技术股份有限公司;SDF-1(CXCL12)中和抗体、CXC基序趋化因子受体(C-X-C motif chemokinereceptor,CXCR)4阻断抗体购自美国R & D公司。引物由生工生物工程(上海)股份有限公司合成。
1.2 原代CAF的分离培养
收集5例PDAC患者手术后新鲜的肿瘤组织样本,使用眼科剪剪碎,用透明质酸酶(0.1 mg/mL)、DNA酶Ⅰ(0.1 mg/mL)、胶原蛋白酶(1 mg/mL)配制的消化酶体系消化1 h,加入适量含10% FBS的RPMI 1640培养基终止消化,用孔径为70 µm的细胞过滤器过滤,离心后去上清。将细胞沉淀重悬后放入培养箱中培养,当细胞融合度达到80%~90% 时,用0.25% 胰蛋白酶消化成单个细胞,按照1 ∶ 1比例传代,12 h后换液,利用成纤维细胞与其他细胞差速贴壁的原理,将贴壁慢的内皮细胞、胰腺癌细胞等杂细胞除去,传代培养至第3~4代时细胞形态趋于一致。
1.3 原代CAF、HDF-a和HFF鉴定
采用流式细胞术鉴定原代CAF、HDF-a和HFF的FAP表达情况。将CAF用0.25% 胰蛋白酶消化成单个细胞,离心、沉淀,使用FACS缓冲液(100 µL/106细胞)+1% FBS重悬细胞,然后加入FAP抗体(10 μL/106细胞),4 ℃环境下避光孵育30 min,上机检测FAP阳性细胞比例。采用细胞免疫荧光技术进行α-SMA和波形蛋白表型鉴定。将胰蛋白酶消化后的CAF用6 mL培养基重悬,在玻片中央滴加数滴细胞悬液,置于37 ℃、5% CO2培养箱培养3 h,待细胞贴壁后用4% 多聚甲醛溶液固定,破膜,滴加α-SMA抗体、波形蛋白抗体,置于湿盒内4 ℃孵育过夜,滴加二抗,室温下避光孵育50 min,滴加DAPI复染细胞核,封片后拍照。
1.4 CAF和HFF中相关细胞因子表达的检测
采用qPCR法检测相关细胞因子mRNA的表达。收集1×106个细胞,提取总RNA并检测RNA纯度,反转录合成cDNA。以cDNA为模板、β- 肌动蛋白为内参照基因进行qPCR扩增。反应条件:95 ℃预变性30 s;95 ℃ 10 s、60 ℃ 10 s、72 ℃10 s,40个循环。引物序列如下:IL-6正向引物为5'-CACTGGTCTTTTGGAGTTTGAG-3',反向引物为5'-GGACTTTTGTACTCATCTGCAC-3';SDF-1正向引物为5'-CCAACGTCAAGCATCTCAAAAT-3',反向引物为5'-CACACTTGTCTGTTGTTGTTCT-3';MCP-1正向引物为5'-AGAATCACCAGCAGCAAGTGTCC-3',反向引物为5'-TTGCTTGTCCAGGTGGTCCATG-3';FAP正向引物为5'-TGGTATAGCAGTGGCTCCAGTCTC-3',反向引物为5'-ATCTGCTGTTCCGTGGATGAGAAG-3';IL-1β正向引物为5'-GCCAGTGAAATGATGGCTTATT-3',反向引物为5'-AGGAGCACTTCATCTGTTTAGG-3';CC基序趋化因子配体(C-C motifchemokine ligand,CCL)5正向引物为5'-CCTCGCTGTCATCCTCATTGCTAC-3',反向引物为5'-CTTGACCTGTGGACGACTGCTG-3';CXCL1正向引物为5'-TCCTGCGAGTGGCACTGCTG-3',反向引物为5'-CTGGCAGCGCAGTTCAGTGG-3'。采用2- ΔΔCt法计算目的基因的相对表达量。
使用ELISA检测试剂盒检测CAF及HFF培养上清中IL-6、MCP-1、SDF-1含量。将原代分离的CAF及HFF以每孔2×105个细胞接种于6孔板,细胞贴壁后,加入1 mL含1% FBS的RPMI 1640培养基,分别培养24、48、72 h,用0.22 µm滤膜过滤去除杂质及细胞碎片,然后按照ELISA检测试剂盒说明书步骤进行检测。
1.5 分选人脾脏MDSC及CD13hi-nMDSC
将PDAC患者脾脏组织置于6 cm培养皿中,并加入适量的无菌PBS,用5 mL注射器橡胶塞末端磨碎,用孔径为70 µm的细胞过滤器过滤2遍,离心后弃上清;加入红细胞裂解液裂解红细胞,离心后重悬;加入HLA-DR、CD33、CD11b、CD14、CD13、CD15抗体(10 µL/106细胞),4 ℃避光孵育30 min,用流式细胞仪分选总MDSC(表型为HLA-DR-/CD33+/CD11b+)和CD13hi-nMDSC(表型为HLA-DR-/CD33+/CD11b+/CD14-/CD15+/CD13+),用于后续实验。
1.6 Transwell系统检测MDSC迁移
收集CAF或HFF培养上清作为条件培养基。对从PDAC患者脾脏组织分选的MDSC进行计数,将1×105个MDSC(分为总MDSC、CD13hi-nMDSC两部分实验)加入Transwell(孔径5 µm)上室。第1部分实验将总MDSC分为6组,分别将普通培养基(RPMI 1640)、CAF条件培养基(20%)、HFF条件培养基(20%)、重组IL-6蛋白(10 ng/mL)、重组SDF-1蛋白(10 ng/mL)、重组MCP-1蛋白(10 ng/mL)加入24孔板中(Transwell下室);第2部分实验将CD13hi-nMDSC分为8组,分别将普通培养基(RPMI 1640)、CAF条件培养基(20%)、HFF条件培养基(20%)、重组IL-6蛋白(10 ng/mL)、重组SDF-1蛋白(10 ng/mL)、重组MCP-1蛋白(10 ng/mL)、CAF条件培养基(20%)+SDF-1(CXCL12)中和抗体(5 µg/mL)、CAF条件培养基(20%)+ CXCR4阻断抗体(10 µg/mL)加入24孔板中(Transwell下室)。将Transwell系统放于细胞培养箱中培养3 h后,用细胞计数仪计数穿入下室的MDSC细胞数。
1.7 统计学处理
应用SPSS 22.0软件进行统计学分析。计量资料以 x±s表示,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析(事后多重比较采用最小显著性差异法)。检验水准(α)为0.05。
2 结果
2.1 PDAC肿瘤组织分离的原代CAF纯度及表型鉴定
CAF由PDAC患者手术标本分离获得,采用差速贴壁的原理纯化3~4代后,光学显微镜下观察可见细胞形态稳定,呈长梭状成纤维样结构,未见内皮细胞等杂细胞(图 1A)。采用流式细胞术检测细胞中活化标志物FAP的表达情况,结果显示分离的原代CAF细胞株中超过90% 的细胞为FAP+细胞,HDF-a也大多表达FAP,但HFF基本为FAP-细胞(图 1B)。采用细胞免疫荧光技术检测细胞中α-SMA和波形蛋白的表达情况,结果显示CAF细胞株均明显表达α-SMA和波形蛋白,HDF-a表达波形蛋白、少量表达α-SMA,HFF表达波形蛋白而α-SMA表达缺失(图 1C)。上述结果提示,分离的原代CAF纯度高,均表达成纤维活化标志物α-SMA、FAP和波形蛋白,符合实验要求;HFF缺失α-SMA和FAP表达,HDF-a表达FAP、少量表达α-SMA,因此选取HFF作为后续实验中的对照细胞。
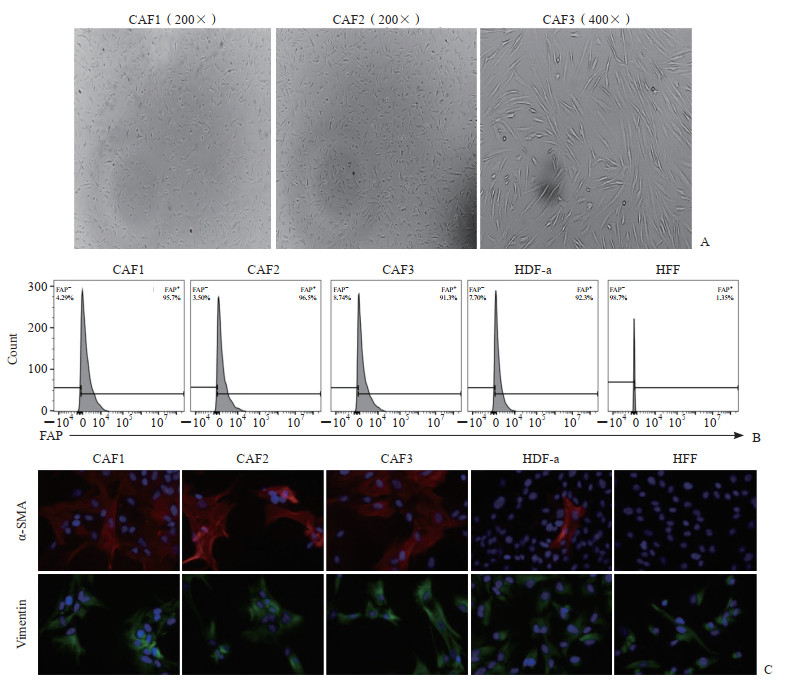 图 1 PDAC肿瘤组织分离的原代CAF纯度及表型鉴定Fig. 1 Purity and phenotypic identification of primary CAFs isolated from PDAC tumor tissuesCAF1, CAF2 and CAF3 were CAFs isolated from tumor tissues of distinct PDAC patients. A: CAFs under microscope; B: The expression levels of FAP on the surface of cells were analyzed by flow cytometry; C: The expression levels of α-SMA and vimentin in cells were analyzed by immunofluorescence staining (200×), and the slide was stained with DAPI (blue), vimentin antibody (green), and α-SMA antibody (red). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; FAP: Fibroblast activation protein; HDF-a: Human dermal fibroblast-adult; HFF: Human foreskin fibroblast; α-SMA: α-smooth muscle actin; DAPI: 4’, 6-diamidino-2-phenylindole.
图 1 PDAC肿瘤组织分离的原代CAF纯度及表型鉴定Fig. 1 Purity and phenotypic identification of primary CAFs isolated from PDAC tumor tissuesCAF1, CAF2 and CAF3 were CAFs isolated from tumor tissues of distinct PDAC patients. A: CAFs under microscope; B: The expression levels of FAP on the surface of cells were analyzed by flow cytometry; C: The expression levels of α-SMA and vimentin in cells were analyzed by immunofluorescence staining (200×), and the slide was stained with DAPI (blue), vimentin antibody (green), and α-SMA antibody (red). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; FAP: Fibroblast activation protein; HDF-a: Human dermal fibroblast-adult; HFF: Human foreskin fibroblast; α-SMA: α-smooth muscle actin; DAPI: 4’, 6-diamidino-2-phenylindole. 下载:
全尺寸图片
下载:
全尺寸图片
2.2 PDAC肿瘤组织分离的原代CAF细胞因子表达
使用qPCR技术检测5株CAF及对照细胞HFF中细胞因子在mRNA水平的表达情况,结果显示CAF中IL-6、MCP-1、SDF-1 mRNA表达量明显高于HFF(均P<0.01,图 2A)。用ELISA试剂盒检测培养24、48、72 h的细胞培养上清中IL-6、MCP-1、SDF-1的含量,结果显示IL-6、MCP-1、SDF-1在CAF培养上清中的含量随着培养时间的延长而升高,并且在各时间点的含量均高于HFF(均P<0.01,图 2B~2D)。
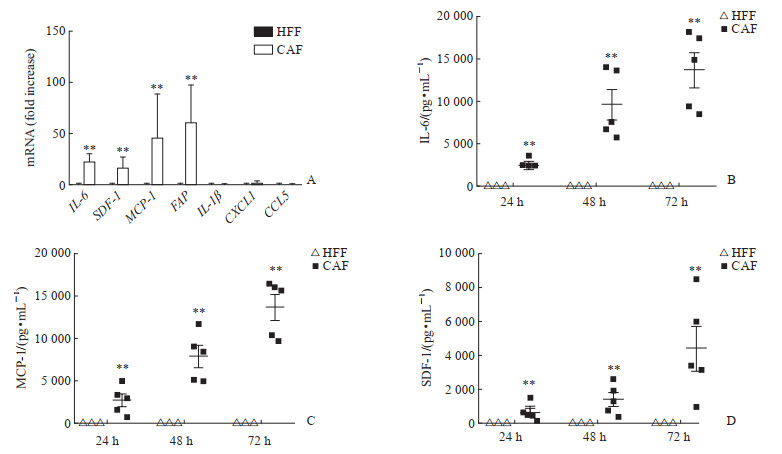 图 2 PDAC肿瘤组织分离的原代CAF中相关细胞因子的表达Fig. 2 Expression of related cytokines in primary CAFs isolated from PDAC tumor tissuesA: mRNA levels of cytokines in CAFs and HFFs were detected by quantitative polymerase chain reaction (**P<0.01 vs HFF. n= 5, x±s); B-D: Contents of IL-6, MCP-1, and SDF-1 in CAF and HFF culture supernatant detected at different time points by enzyme-like immunosorbent assay (**P<0.01 vs HFF. x±s). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; HFF: Human foreskin fibroblast; IL: Interleukin; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; FAP: Fibroblast activation protein; CXCL1: C-X-C motif chemokine ligand 1; CCL5: C-C motif chemokine ligand 5.下载:
全尺寸图片
图 2 PDAC肿瘤组织分离的原代CAF中相关细胞因子的表达Fig. 2 Expression of related cytokines in primary CAFs isolated from PDAC tumor tissuesA: mRNA levels of cytokines in CAFs and HFFs were detected by quantitative polymerase chain reaction (**P<0.01 vs HFF. n= 5, x±s); B-D: Contents of IL-6, MCP-1, and SDF-1 in CAF and HFF culture supernatant detected at different time points by enzyme-like immunosorbent assay (**P<0.01 vs HFF. x±s). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; HFF: Human foreskin fibroblast; IL: Interleukin; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; FAP: Fibroblast activation protein; CXCL1: C-X-C motif chemokine ligand 1; CCL5: C-C motif chemokine ligand 5.下载:
全尺寸图片
2.3 CAF通过SDF-1/CXCR4通路促进CD13hinMDSC的迁移
由图 3可见,CAF条件培养基比普通培养基(RPMI 1640)和正常成纤维细胞HFF条件培养基招募更多的总MDSC和CD13hinMDSC(均P<0.01)。在Transwell迁移实验模型下室分别加入重组IL-6、SDF-1、MCP-1蛋白,结果显示重组IL-6蛋白不能引起总MDSC和CD13hi-nMDSC的迁移(图 3A、3B);虽然MCP-1也可以诱导总MDSC和CD13hi-nMDSC迁移,但CD13hi-nMDSC迁移的数量明显少于SDF-1(图 3B);重组SDF-1蛋白可以诱导总MDSC和CD13hi-nMDSC的迁移(均P<0.01)。因此,我们判断SDF-1、MCP-1参与了PDAC中MDSC的迁移,IL-6在MDSC迁移中并无作用。为了进一步证实SDF-1的作用,在CAF条件培养基中同时加入SDF-1中和抗体或CXCR4阻断抗体进行CD13hi-nMDSC迁移实验,结果显示加入SDF-1中和抗体或CXCR4阻断抗体后能够明显减少由CAF条件培养基诱导的CD13hi-nMDSC迁移(均P<0.01)。这提示CAF可通过SDF-1/CXCR4通路诱导CD13hi-nMDSC的迁移。
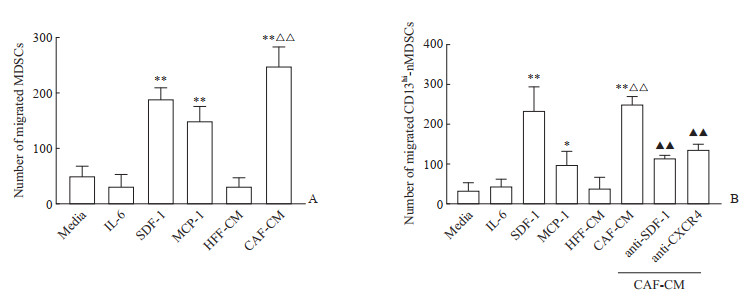 图 3 CAF-CM及相关细胞因子对MDSC迁移的影响Fig. 3 Effects of CAF-CM and related cytokines on MDSC migrationA : The average number of migrated MDSCs was detected by Transwell assay; B: The average number of migrated CD13hi-nMDSCs was detected by Transwell assay. *P<0.05, **P<0.01 vs HFF-CM; △△P<0.01 vs media; ▲▲P<0.01 vs CAF-CM. n= 5, x±s. CAF: Cancer-associated fibroblast; CM: Conditioned medium; MDSC: Myeloid-derived suppressor cell; IL-6: Interleukin 6; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; HFF: Human foreskin fibroblast; CD13hi: CD13 high expressed; nMDSC: Neutrophil-like MDSC; CXCR4: C-X-C motif chemokine receptor 4.下载:
全尺寸图片
图 3 CAF-CM及相关细胞因子对MDSC迁移的影响Fig. 3 Effects of CAF-CM and related cytokines on MDSC migrationA : The average number of migrated MDSCs was detected by Transwell assay; B: The average number of migrated CD13hi-nMDSCs was detected by Transwell assay. *P<0.05, **P<0.01 vs HFF-CM; △△P<0.01 vs media; ▲▲P<0.01 vs CAF-CM. n= 5, x±s. CAF: Cancer-associated fibroblast; CM: Conditioned medium; MDSC: Myeloid-derived suppressor cell; IL-6: Interleukin 6; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; HFF: Human foreskin fibroblast; CD13hi: CD13 high expressed; nMDSC: Neutrophil-like MDSC; CXCR4: C-X-C motif chemokine receptor 4.下载:
全尺寸图片
3 讨论
PDAC是胰腺癌最常见的病理类型,具有早期诊断困难、进展快、对放化疗不敏感、预后差等特点,尽管近年来医疗水平不断提高,但PDAC患者的5年预期生存率仍不足10%[1]。免疫治疗在多种恶性肿瘤治疗中取得实质性进展,但在PDAC的治疗中获益有限,这与PDAC独特的免疫抑制微环境密切相关[13]。
MDSC是肿瘤微环境中最重要的免疫抑制细胞,其在肿瘤进展过程中大量扩增、募集,通过参与肿瘤免疫逃逸、促进肿瘤血管生成及增强肿瘤细胞的侵袭与迁移驱动肿瘤进展[6]。MDSC在食管癌、肝癌、乳腺癌、胰腺癌等肿瘤中明显增多,并且MDSC数量与肿瘤患者预后、免疫治疗疗效密切相关[11, 14-16]。有研究证实靶向参与招募MDSC的趋化因子受体能有效提高免疫治疗的抗肿瘤疗效[17]。临床前研究显示,在多种小鼠恶性肿瘤模型中,阻断CCL2/CC基序趋化因子受体(C-Cmotif chemokine receptor,CCR)2轴,可以减少肿瘤内MDSC的数量并提高抗肿瘤疗效[18-20];CXCR1/2抑制剂能减少MDSC募集并激活小鼠模型T细胞应答,提高免疫治疗效果[21]。CCR5及其配体CCL3、CCL4、CCL5参与调控MDSC的募集,阻断CCR5可降低乳腺癌、胰腺癌、结直肠癌和前列腺癌的肿瘤生长和转移潜能[17]。CSF-1R在乳腺癌和胰腺癌中表达上调,靶向CSF-1R可以抑制MDSC向肿瘤部位的募集并强化免疫检查点抑制剂、过继T细胞疗法的抗肿瘤应答[22-23]。靶向nMDSC表面的关键功能受体CD300ld能够抑制nMDSC的募集及其T细胞抑制功能,联合使用CD300ld阻断剂能提高免疫治疗效果[24]。因此,研究MDSC的募集机制对提高PDAC的免疫治疗疗效有重要意义。我们在前期研究中发现PDAC患者的外周血及肿瘤组织中nMDSC数量与健康受试者比较显著升高,并且发现了CD13hi-nMDSC和CD13低表达(CD13low)-nMDSC这2个新亚群,PDAC肿瘤组织中以CD13hi-nMDSC亚群增多为主;CD13hi-nMDSC分泌大量的精氨酸酶1抑制T细胞增殖,是发挥免疫抑制功能的主要细胞群[11]。但CD13hi-nMDSC在胰腺癌肿瘤组织中大量募集的具体机制尚不明确。
Caf是胰腺癌间质中的主要细胞成分,在肿瘤微环境中产生大量的胶原蛋白等细胞外基质,限制药物进入肿瘤核心部位,充当免疫细胞浸润的屏障,帮助胰腺癌细胞逃避宿主免疫监视[25]。已在肝癌、胰腺癌、胆管细胞癌等多种肿瘤的小鼠模型中证实Caf参与了Mdsc的扩增、募集[26]。本研究从5例Pdac患者的肿瘤组织中分离了原代Caf,通过Qpcr、Elisa技术筛选出Caf与对照细胞Hff表达差异较大的3种细胞因子Il-6、Sdf-1、Mcp-1,并发现Caf条件培养基能够促进cd13Hi-Nmdsc的迁移。为探讨Caf条件培养基促进Mdsc迁移的作用机制,在培养体系中分别加入重组Il-6、Sdf-1、Mcp-1蛋白,结果发现Sdf-1重组蛋白可以诱导Cd13Hi-Nmdsc的迁移,并且加入Sdf-1中和抗体后能够明显减少由Caf条件培养基诱导的Cd13Hi-Nmdsc迁移。以上结果说明,Caf主要通过sdf-1介导cd13Hi-Nmdsc的迁移。
SDF-1又称CXCL12,属于CXC家族趋化因子,能够激活2种具有不同下游信号通路的趋化因子受体:CXCR4和CXCR7。CXCR4在外周血淋巴细胞、单核细胞、中性粒细胞、树突状细胞等多种免疫细胞表面及肿瘤细胞中均有明显表达[27];在肝癌、乳腺癌、骨肉瘤中证实MDSC表面也表达CXCR4[10, 28],而且SDF-1是CXCR4的唯一配体[29]。Deng等[30]在体外细胞实验及小鼠肝癌模型中证实CXCL12能够促进MDSC向肿瘤组织迁移,并且CXCR4是肝癌患者预后不良的重要标志物之一。在胰腺癌中,有多项研究证实联合靶向CXCR4策略可以提高多种免疫治疗的效果[31-32]。因此,我们推断SDF-1的2个受体中,CXCR4可能起主要作用。为进一步研究SDF-1参与调节PDAC中CD13hi-nMDSC迁移的具体机制,在迁移体系中加入CXCR4阻断抗体,发现加入CXCR4阻断抗体后能够明显减少由CAF条件培养基诱导的MDSC迁移,说明CAF可以通过SDF-1/CXCR4通路促进CD13hi-nMDSC这一新亚群的迁移。
CAF是一群异质性较大的细胞群,在不同胰腺癌患者中分离的CAF表型特征及细胞因子的表达量有一定差异。本研究仅分析了5株原代CAF与CD13hi-nMDSC迁移的关系,SDF-1的另一配体CXCR7是否在CD13hi-nMDSC迁移中发挥作用尚未研究,具有一定的局限性。在后续实验中,我们会增加样本量进一步研究CAF与CD13hi-nMDSC迁移的相关机制。
综上所述,本研究对PDAC肿瘤微环境中的CAF与MDSC迁移的相关关系进行了探讨,发现CAF能够通过分泌的SDF-1促进MDSC的迁移,尤其是CAF可以通过SDF-1/CXCR4通路招募CD13hi-nMDSC这一发挥免疫抑制功能的主要细胞群,因此参与CD13hi-nMDSC亚群招募的关键因子SDF-1及其受体CXCR4可能是PDAC免疫治疗的重要潜在靶点。
-
图 1 PDAC肿瘤组织分离的原代CAF纯度及表型鉴定
Fig. 1 Purity and phenotypic identification of primary CAFs isolated from PDAC tumor tissues
CAF1, CAF2 and CAF3 were CAFs isolated from tumor tissues of distinct PDAC patients. A: CAFs under microscope; B: The expression levels of FAP on the surface of cells were analyzed by flow cytometry; C: The expression levels of α-SMA and vimentin in cells were analyzed by immunofluorescence staining (200×), and the slide was stained with DAPI (blue), vimentin antibody (green), and α-SMA antibody (red). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; FAP: Fibroblast activation protein; HDF-a: Human dermal fibroblast-adult; HFF: Human foreskin fibroblast; α-SMA: α-smooth muscle actin; DAPI: 4’, 6-diamidino-2-phenylindole.
下载:
全尺寸图片
图 2 PDAC肿瘤组织分离的原代CAF中相关细胞因子的表达
Fig. 2 Expression of related cytokines in primary CAFs isolated from PDAC tumor tissues
A: mRNA levels of cytokines in CAFs and HFFs were detected by quantitative polymerase chain reaction (**P<0.01 vs HFF. n= 5, x±s); B-D: Contents of IL-6, MCP-1, and SDF-1 in CAF and HFF culture supernatant detected at different time points by enzyme-like immunosorbent assay (**P<0.01 vs HFF. x±s). PDAC: Pancreatic ductal adenocarcinoma; CAF: Cancer-associated fibroblast; HFF: Human foreskin fibroblast; IL: Interleukin; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; FAP: Fibroblast activation protein; CXCL1: C-X-C motif chemokine ligand 1; CCL5: C-C motif chemokine ligand 5.
下载:
全尺寸图片
图 3 CAF-CM及相关细胞因子对MDSC迁移的影响
Fig. 3 Effects of CAF-CM and related cytokines on MDSC migration
A : The average number of migrated MDSCs was detected by Transwell assay; B: The average number of migrated CD13hi-nMDSCs was detected by Transwell assay. *P<0.05, **P<0.01 vs HFF-CM; △△P<0.01 vs media; ▲▲P<0.01 vs CAF-CM. n= 5, x±s. CAF: Cancer-associated fibroblast; CM: Conditioned medium; MDSC: Myeloid-derived suppressor cell; IL-6: Interleukin 6; SDF-1: Stromal cell-derived factor 1; MCP-1: Monocyte chemotactic protein 1; HFF: Human foreskin fibroblast; CD13hi: CD13 high expressed; nMDSC: Neutrophil-like MDSC; CXCR4: C-X-C motif chemokine receptor 4.
下载:
全尺寸图片
-
[1] SARFATY E, KHAJOUEINEJAD N, ZEWDE M G, et al. Surgical management of pancreatic ductal adenocarcinoma: a narrative review[J]. Transl Gastroenterol Hepatol, 2023, 8: 39. DOI: 10.21037/tgh-23-27. [2] XIA C, DONG X, LI H, et al. Cancer statistics in China and United States, 2022: profiles, trends, and determinants[J]. Chin Med J (Engl), 2022, 135(5): 584- 590. DOI: 10.1097/CM9.0000000000002108. [3] SIEGEL R L, MILLER K D, WAGLE N S, et al. Cancer statistics, 2023[J]. CA A Cancer J Clin, 2023, 73(1): 17- 48. DOI: 10.3322/caac.21763. [4] VON AHRENS D, BHAGAT T D, NAGRATH D, et al. The role of stromal cancer-associated fibroblasts in pancreatic cancer[J]. J Hematol Oncol, 2017, 10(1): 76. DOI: 10.1186/s13045-017-0448-5. [5] HO W J, JAFFEE E M, ZHENG L. The tumour microenvironment in pancreatic cancer: clinical challenges and opportunities[J]. Nat Rev Clin Oncol, 2020, 17(9): 527-540. DOI: 10.1038/s41571-020-0363-5. [6] GABRILOVICH D I. Myeloid-derived suppressor cells[J]. Cancer Immunol Res, 2017, 5(1): 3-8. DOI: 10.1158/2326-6066.CIR-16-0297. [7] RIES C H, HOVES S, CANNARILE M A, et al. CSF-1/CSF-1R targeting agents in clinical development for cancer therapy[J]. Curr Opin Pharmacol, 2015, 23: 45- 51. DOI: 10.1016/j.coph.2015.05.008. [8] KUMAR V, DONTHIREDDY L, MARVEL D, et al. Cancer-associated fibroblasts neutralize the anti-tumor effect of CSF1 receptor blockade by inducing PMNMDSC infiltration of tumors[J]. Cancer Cell, 2017, 32(5): 654-668. e5. DOI: 10.1016/j.ccell.2017.10.005. [9] LIU Y, SUN Y, WANG P, et al. FAP-targeted CAR-T suppresses MDSCs recruitment to improve the antitumor efficacy of claudin18.2-targeted CAR-T against pancreatic cancer[J]. J Transl Med, 2023, 21(1): 255. DOI: 10.1186/s12967-023-04080-z. [10] SUN R, LUO H, SU J, et al. Olaparib suppresses MDSC recruitment via SDF1α/CXCR4 axis to improve the anti-tumor efficacy of CAR-T cells on breast cancer in mice[J]. Mol Ther, 2021, 29(1): 60-74. DOI: 10.1016/j.ymthe.2020.09.034. [11] ZHANG J, XU X, SHI M, etal. CD13 hi neutrophil-like myeloid-derived suppressor cells exert immune suppression through arginase 1 expression in pancreatic ductal adenocarcinoma[J]. Oncoimmunology, 2017, 6(2): e1258504. DOI: 10.1080/2162402X.2016.1258504. [12] 张冰冰, 唐海双, 张晶, 等. 肿瘤相关成纤维细胞通过IL-6/STAT3通路介导胰腺癌中髓源性抑制细胞的分化[J]. 第二军医大学学报, 2019, 40(5): 520-527. DOI: 10.16781/j.0258-879x.2019.05.0520. ZHANG B B, TANG H S, ZHANG J, et al. Cancerassociated fibroblasts mediate differentiation of myeloid-derived suppressor cells in pancreatic cancer through IL-6/STAT3 pathway[J]. Acad J Sec Mil Med Univ, 2019, 40(5): 520-527. DOI: 10.16781/j.0258-879x.2019.05.0520. [13] ULLMAN N A, BURCHARD P R, DUNNE R F, et al. Immunologic strategies in pancreatic cancer: making Cold tumors Hot[J]. J Clin Oncol, 2022, 40(24): 2789- 2805. DOI: 10.1200/JCO.21.02616. [14] OHKI S, SHIBATA M, GONDA K, et al. Circulating myeloid-derived suppressor cells are increased and correlate to immune suppression, inflammation and hypoproteinemia in patients with cancer[J]. Oncol Rep, 2012, 28(2): 453-458. DOI: 10.3892/or.2012.1812. [15] GABITASS R F, ANNELS N E, STOCKEN D D, et al. Elevated myeloid-derived suppressor cells in pancreatic, esophageal and gastric cancer are an independent prognostic factor and are associated with significant elevation of the Th2 cytokine interleukin-13[J]. Cancer Immunol Immunother, 2011, 60(10): 1419-1430. DOI: 10.1007/s00262-011-1028-0. [16] DIAZ-MONTERO C M, SALEM M L, NISHIMURA M I, et al. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy[J]. Cancer Immunol Immunother, 2009, 58(1): 49-59. DOI: 10.1007/s00262-008-0523-4. [17] JOSHI S, SHARABI A. Targeting myeloid-derived suppressor cells to enhance natural killer cell-based immunotherapy[J]. Pharmacol Ther, 2022, 235: 108114. DOI: 10.1016/j.pharmthera.2022.108114. [18] WANG Y, ZHANG X, YANG L, et al. Blockade of CCL2 enhances immunotherapeutic effect of anti-PD1 in lung cancer[J]. J Bone Oncol, 2018, 11: 27-32. DOI: 10.1016/j.jbo.2018.01.002. [19] FLORES-TORO J A, LUO D, GOPINATH A, et al. CCR2 inhibition reduces tumor myeloid cells and unmasks a checkpoint inhibitor effect to slow progression of resistant murine gliomas[J]. Proc Natl Acad Sci USA, 2020, 117(2): 1129-1138. DOI: 10.1073/pnas.1910856117. [20] LESOKHIN A M, HOHL T M, KITANO S, et al. Monocytic CCR2+ myeloid-derived suppressor cells promote immune escape by limiting activated CD8 T-cell infiltration into the tumor microenvironment[J]. Cancer Res, 2012, 72(4): 876-886. DOI: 10.1158/0008-5472.CAN-11-1792. [21] GREENE S, ROBBINS Y, MYDLARZ W K, et al. Inhibition of MDSC trafficking with SX-682, a CXCR1/2 inhibitor, enhances NK-cell immunotherapy in head and neck cancer models[J]. Clin Cancer Res, 2020, 26(6): 1420-1431. DOI: 10.1158/1078-0432.CCR-19-2625. [22] MOK S, KOYA R C, TSUI C, et al. Inhibition of CSF-1 receptor improves the antitumor efficacy of adoptive cell transfer immunotherapy[J]. Cancer Res, 2014, 74(1): 153-161. DOI: 10.1158/0008-5472.CAN-13-1816. [23] HOLMGAARD R B, ZAMARIN D, LESOKHIN A, et al. Targeting myeloid-derived suppressor cells with colony stimulating factor-1 receptor blockade can reverse immune resistance to immunotherapy in indoleamine 2, 3-dioxygenase-expressing tumors[J]. EBioMedicine, 2016, 6: 50-58. DOI: 10.1016/j.ebiom.2016.02.024. [24] WANG C, ZHENG X, ZHANG J, et al. CD300ld on neutrophils is required for tumour-driven immune suppression[J]. Nature, 2023, 621(7980): 830-839. DOI: 10.1038/s41586-023-06511-9. [25] YUAN S, MU W, LIU S, et al. Transforming cancerassociated fibroblast barrier into drug depots to boost chemo-immunotherapy in "shooting fish in a barrel" pattern[J]. ACS Nano, 2023, 17(14): 13611-13626. DOI: 10.1021/acsnano.3c02272. [26] KOMOHARA Y, TAKEYA M. CAFs and TAMs: maestros of the tumour microenvironment[J]. J Pathol, 2017, 241(3): 313-315. DOI: 10.1002/path.4824. [27] CHEN F H, FU S Y, YANG Y C, et al. Combination of vessel-targeting agents and fractionated radiation therapy: the role of the SDF-1/CXCR4 pathway[J]. Int J Radiat Oncol Biol Phys, 2013, 86(4): 777-784. DOI: 10.1016/j.ijrobp.2013.02.036. [28] JIANG K, LI J, ZHANG J, et al. SDF-1/CXCR4 axis facilitates myeloid-derived suppressor cells accumulation in osteosarcoma microenvironment and blunts the response to anti-PD-1 therapy[J]. Int Immunopharmacol, 2019, 75: 105818. DOI: 10.1016/j.intimp.2019.105818. [29] SCHIMANSKI C C, BAHRE R, GOCKEL I, et al. Dissemination of hepatocellular carcinoma is mediated via chemokine receptor CXCR4[J]. Br J Cancer, 2006, 95(2): 210-217. DOI: 10.1038/sj.bjc.6603251. [30] DENG Y, CHENG J, FU B, et al. Hepatic carcinomaassociated fibroblasts enhance immune suppression by facilitating the generation of myeloid-derived suppressor cells[J]. Oncogene, 2017, 36(8): 1090-1101. DOI: 10.1038/onc.2016.273. [31] BOCKORNY B, SEMENISTY V, MACARULLA T, et al. BL-8040, a CXCR4 antagonist, in combination with pembrolizumab and chemotherapy for pancreatic cancer: the COMBAT trial[J]. Nat Med, 2020, 26(6): 878-885. DOI: 10.1038/s41591-020-0880-x. [32] SEO Y D, JIANG X, SULLIVAN K M, et al. Mobilization of CD8+ T cells via CXCR4 blockade facilitates PD-1 checkpoint therapy in human pancreatic cancer[J]. Clin Cancer Res, 2019, 25(13): 3934-3945. DOI: 10.1158/1078-0432.CCR-19-0081.
