 2019, Vol. 37
2019, Vol. 37扩展功能
文章信息
- 王龙, 李伟强, 王皓, 吴海, 辛浩
- WANG Long, LI WeiQiang, WANG Hao, WU Hai, XIN Hao
- 华北地台马沟剖面寒武系苗岭统微生物岩的组构特征、沉积环境和地质意义
- Fabric Characteristics, Depositional Environment and Geological Implications of Cambrian Miaolingian Microbialites at Magou Section, North China Platform
- 沉积学报, 2019, 37(6): 1193-1209
- ACTA SEDIMENTOLOGICA SINCA, 2019, 37(6): 1193-1209
- 10.14027/j.issn.1000-0550.2019.018
-
文章历史
- 收稿日期:2018-12-25
- 收修改稿日期: 2019-02-01



2. 中国石油长庆油田公司勘探开发研究院, 西安 710018;
3. 中国石油勘探开发研究院, 北京 100086;
4. 中国石油杭州地质研究院, 杭州 310023
2. Research Institute of Exploration and Development, PetroChina Changqing Oil field Company, Xi'an 710018, China;
3. PetroChina, Research Institute of Petroleum Exploration and Development, Beijing, 100086, China;
4. PetroChina Hangzhou Research Institute of Geology, Hangzhou 310023, China
寒武纪与地球生物学相关的碳酸盐沉积作用大致可以分为两个阶段:1)随着寒武纪生命大爆发和后生动物的辐射,古杯类等后生动物繁盛在寒武纪早期的生物礁中,古杯动物—钙化微生物礁(archaeocyath-calcimicrobe reefs)遍布于各种各样的低纬度(大多为浅海)环境[1-2];2)从寒武纪第二世末期到奥陶纪早期,后生动物生物礁变得越来越稀少[3],多样性也显著降低,直到早奥陶世才开始复苏[2, 4]。这段时间全球以大量繁盛的微生物岩(礁)为特征[4-7],Riding[8-13]称其为“显生宙早期第一幕蓝细菌钙化作用事件”的发生时期或微生物岩的“寒武纪—早奥陶世复苏期”,不仅为研究地质历史时期与微生物活动相关的生物/有机矿化作用过程提供了有用信息,也是了解地球表层环境演变的一个重要窗口[8-9, 14]。
华北地台大致从寒武纪第二世的晚期开始接受沉积,超覆在前寒武—寒武纪“巨型不整合面”之上[15],形成了一套二级海侵背景下的厚层陆表海硅质碎屑岩—碳酸盐岩混积序列[16-17]。这套特别的地层序列在苗岭统和芙蓉统中包含有多种多样的微生物岩。通常认为苗岭统的微生物岩(礁)以凝块石和树形石为特征,主要由附枝菌等钙化微生物所组成[2-3, 5-7];而芙蓉统的微生物岩主要由迷宫状微生物岩(礁)和柱状叠层石所组成,几乎没有附枝菌[5-7, 18]。本次研究率先发现和详细描述了华北地台中北部马沟剖面苗岭统迷宫状微生物岩、均一石和叠层石的层序位置、沉积组构和环境属性,并对其成因作了可能性的解释。这为认识寒武系微生物岩(礁)的多样化构成提供了一个典型实例,也为理解微生物岩复杂多样的沉积作用和形成机制提供了重要线索。
1 地质背景 1.1 剖面位置及概况马沟剖面位于河北省唐县县城西北部约30 km处的迷城乡至马沟村的公路旁(图 1),主要出露的地层包括馒头组至芙蓉统,与北京西山剖面相比较,缺失昌平组,馒头组直接平行不整合面覆盖在新元古界长龙山组之上。馒头组下部为紫红色和砖红色厚层块状白云质泥岩夹少量泥质白云岩透镜体,厚度为36 m左右;上部为厚度5 m左右的浅灰色、灰黄色泥晶白云岩和泥质白云岩。馒头组之上即为由毛庄组、徐庄组、张夏组和崮山组构成的苗岭统[19],在其高水位体系域和强迫型海退体系域的碳酸盐岩单元中,常常包含特别的由微生物岩主导的生物层、生物丘或生物丘复合体。

苗岭统之上的芙蓉统长山组自下而上由陆棚相钙质泥页岩,深缓坡相中薄层钙质泥页岩、中薄层泥质条带泥晶灰岩和中厚层泥质条带泥晶灰岩构成,顶部局部发育10 m左右的块状泥晶生物丘。凤山组由两个三级层序构成,每个层序都从深灰绿色及深褐色钙质泥页岩陆棚相沉积,中薄层灰绿色及深灰色钙质泥页岩夹竹叶状砾屑灰岩透镜体、与灰色中厚层泥质条带泥晶灰岩组成的潮下米级旋回构成深缓坡至中缓坡相沉积,到中薄层泥质条带泥晶灰岩与厚层块状泥晶生物丘或生物潜穴灰岩组成的潮下型米级旋回构成中缓坡至浅缓坡相沉积。寒武系顶部被下奥陶统冶里组底部的条带状泥灰岩地层整合覆盖。
1.2 层序地层特征马沟剖面寒武系苗岭统发育较为完整,包括毛庄组、徐庄组、张夏组和崮山组,分别构成4个三级层序即SQ1至SQ4(图 2)。这些三级层序由米级旋回有序叠加形成的各具特色的沉积相序列所构成,反映了海平面的升降变化及沉积学响应[20]。其中,以在SQ2的高位体系域和强迫型海退体系域,以及SQ3和SQ4的高位体系域中发育的生物层、生物丘或生物丘复合体最为特别(图 2)。

|
| 图 2 马沟剖面寒武系苗岭统层序地层划分 Fig.2 Sequence stratigraphic division of the Cambrian Miaolingian at the Magou section |
毛庄组构成三级层序SQ1,下部为紫红色和砖红色厚层块状白云质泥岩和粉砂质泥岩夹少量泥质白云岩透镜体,厚度为49 m左右,构成层序的海侵体系域;上部的高水位体系域为浅灰色和灰黄色泥晶白云岩和泥质白云岩,中部夹有潮间坪下部的泥晶白云岩,总厚12 m左右(图 2)。徐庄组构成三级层序SQ2:1)下部由潮上坪块状紫红色泥岩和粉砂质泥岩所组成,向上潮间坪相的砂泥质白云岩和泥质粉砂岩夹层逐渐增多,总厚38 m左右;2)厚度为15 m左右的灰绿色粉砂质钙质泥页岩地层,夹泥晶灰岩和泥灰岩透镜体和透镜层,为陆棚相沉积,代表徐庄组构成的三级层序的凝缩段;3)灰色和灰白色块状生物丘(层)灰岩,夹少量钙质泥页岩地层,构成该三级层序的高位体系域和强迫型海退体系域,厚度为58 m左右,是该剖面最为特别之处(图 2)。
张夏组构成三级层序SQ3,包含3个四级亚层序。第一个亚层序下部为24 m厚的灰绿色及灰色陆棚相钙质泥页岩夹泥灰岩透镜体,局部夹有竹叶状灰岩透镜体,组成陆棚相地层;中部为中灰绿色厚层钙质泥页岩夹竹叶状砾屑灰岩透镜体、中薄层泥质条带泥晶灰岩组成的L-M型及厚基底类潮下型米级旋回,组成深缓坡相地层,厚度为7 m左右;上部为中灰绿色厚层钙质泥页岩夹竹叶状砾屑灰岩透镜体、中薄层泥质条带泥晶灰岩、厚层含生物碎屑鲕粒灰岩组成潮下型米级旋回,在鲕粒灰岩中见较多的竹叶状砾屑,组成中至浅缓坡相地层,厚度为5 m左右(图 2)。第二个亚层序的组成包括:1)25 m厚度的灰绿色及灰色陆棚相钙质泥页岩夹泥灰岩透镜体,局部夹有竹叶状灰岩透镜体,组成陆棚相地层;2)中灰绿色厚层钙质泥页岩夹竹叶状砾屑灰岩透镜体、中薄层泥质条带泥晶灰岩组成的L-M型及厚基底类潮下型米级旋回,组成深缓坡相地层,厚度为16 m左右;3)中薄层泥质条带泥晶灰岩,与厚层鲕粒灰岩或块状泥晶生物丘灰岩共同组成潮下型米级旋回,构成浅缓坡相地层,厚度为12 m左右。其中第3部分地层的下部以厚度为3~5 m的鲕粒灰岩为主,在鲕粒灰岩中见小型泥晶生物丘;上部为6~8 m的块状泥晶生物丘灰岩(图 2)。第三个亚层序组成包括:1)中灰绿色厚层钙质泥页岩夹竹叶状砾屑灰岩透镜体、中薄层泥质条带泥晶灰岩组成的L-M型及厚基底类潮下型米级旋回,组成陆棚相至深缓坡相沉积,厚度为15 m左右;2)中薄层泥质条带泥晶灰岩,与中厚层鲕粒灰岩共同组成潮下型米级旋回,构成浅缓坡相沉积,厚度为10 m左右;3)中薄层泥质条带泥晶灰岩(单层厚度为厘米级别,1~8 cm),与块状(单层厚度1~3 m)鲕粒滩相灰岩互层组成潮下型米级旋回,组成鲕粒滩相沉积,厚15 m左右(图 2)。
崮山组构成三级层序SQ4:1)深灰色及灰绿色钙质泥页岩,夹泥灰岩和泥晶灰岩透镜体和透镜层,发育异地型风暴竹叶状砾屑灰岩透镜体,厚度为24 m左右;2)中薄层钙质泥页岩与中厚层泥晶灰岩互层组成潮下型米级旋回,组成中至浅缓坡相沉积,夹一层厚度为0.3~1 m左右且呈波状起伏的泥晶生物丘灰岩,总厚度为12 m左右;3)中薄层泥质条带泥晶灰岩,与厚层块状鲕粒滩相灰岩互层组成潮下型米级旋回,组成鲕粒滩相地层,厚度10 m左右(图 2)。
2 迷宫状微生物岩(礁) 2.1 宏观特征微生物岩是指与微生物生长、新陈代谢、细胞表面性质,以及细胞外聚合物(Extracellular Polymeric Substances,EPS)相互作用有关的沉积物(岩)[2-4, 12],是微生物席或微生物膜的钙化作用产物[10, 14]。基于宏观形态学特征(中等尺度构造),常被划分为纹层状的叠层石、凝块状的凝块石、贫乏构造的均一石和树枝状的树形石等[1-4, 11, 14]。迷宫状微生物岩(maze-like microbialites)最早被描述为凝块石[21],后来因发育的普遍性而被Shapiro et al.[22]列为凝块石的一种特殊类别,并将厘米至分米级别的分叉迷宫状、“栅栏”式构造命名为迷宫体(maceriae)。Lee et al. [5],Chen et al.[6]则因其结构特殊而直接取名迷宫状微生物岩(礁),并进一步分类为板格状迷宫状微生物岩、柱状迷宫状微生物岩和柱状杂乱的微生物岩(表 1)。陈吉涛等[7]则根据迷宫和迷宫间沉积组分的不同,将迷宫状生物岩(礁)分为泥质迷宫状微生物岩(礁),颗粒状迷宫微生物岩(礁)和白云质迷宫微生物岩(礁)3类。
| 微生物岩类型 | 整体构造 | 内部结构 | 示意图 | 描述 | 沉积环境 |
| 板格状 | 板格状 | 迷宫状 | 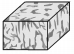
|
呈生物丘或者生物层样式产出;由迷宫体和迷宫体之间的沉积物两部分构成,之间界限模糊;迷宫体内部结构杂乱;迷宫体之间沉积物为灰泥和生肩;迷宫由泥晶、葛万菌等组成 | 低能正常浪基面以下 |
| 迷宫状微生物岩(类型Ⅰ) | 杂乱状 | ||||
| 柱状 | 柱状 | 迷宫状 | 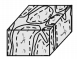
|
生物层内可见柱状构造(> 60cm); 柱体内呈现迷宫构造,边缘杂乱状;柱体间为粗粒沉积物;类型Ⅱ微生物岩向上转变为类型Ⅲ微生物岩 | 中能正常浪基面附近 |
| 迷宫状微生物岩(类型Ⅱ) | 杂乱状 | ||||
| 柱状 | 柱状 | 杂乱状 | 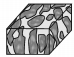
|
可见柱状构造(直径10~15cm); 柱体的伸长率从3:1到10:1;柱体边缘平滑清晰;镜下可见泥晶、亮晶、葛万菌肾形菌、生肩等;柱体之间的沉积物由球粒和葛万菌等微生物碎片组成 | 高能正常浪基面之上 |
| 杂乱微生物岩(类型Ⅲ) |
若按照微生物岩巨型、大型、中型和微型构造的命名原则[22-23],依据构成迷宫状微生物岩的迷宫体或柱体的宏观组构(中型构造),可将其归类到凝块石、叠层石或均一石等[8, 22-26]。但鉴于其分布的广泛性,结构的特殊性和描述的方便性,本文将采用迷宫状微生物岩这一命名,并根据迷宫体的形态、组构,迷宫和迷宫间沉积的组分,马沟剖面迷宫状微生物岩大致可分为3种类别(图 3),主要产出在SQ2高位体系域(图 2)。

|
| 图 3 马沟剖面苗岭统徐庄组迷宫状微生物岩的宏观特征 (a)类型Ⅰ迷宫状微生物岩,切面上呈现出复杂的迷宫状构造,结构杂乱;(b)是(a)的素描;(c)是(a)的局部特写,可见明显的凝块和树形结构;(d)类型Ⅱ迷宫状微生物岩,迷宫体与周围沉积物之间的边界模糊,而且不规则,被选择白云岩化作用加强;(e)是介于类型Ⅱ和类型Ⅲ之间的过渡类型;(f)是(e)的素描,包括纵切面和横切面两个方位;(g)类型Ⅲ迷宫状微生物岩,迷宫表现为分叉或会聚的柱状体,柱体之间被三叶虫、双壳类的生物碎屑、鲕粒和少量的内碎屑充填;(h)是(g)的局部特写,柱状体中可见一些生物碎屑 Fig.3 Megascopic features of maceriate microbialites of Miaolingian Series at the Magou section (a) TypeⅠ maceriate microbialites showing complex maze-like structures and random distributions; (b) is the sketch of (a); (c) is the enlargement of (a): note the meso-clots and dendrites; (d) Type Ⅱ maceriate microbialites. The boundaries between the maceriae and surrounding sediments are blurred and irregular, but are en-hanced by selected dolomitization; (e) is the transitional type between Types Ⅱ and Ⅲ; (f) is the sketch of (e), including both vertical and transverse sections; (g) Type Ⅲ maceriate microbialites, showing branched or convergent columnar shapes, with biodebris such as trilobites and bivalves, oolitic grains and a small amount of internal debris fillings; and (h) is the enlargement of (g) expressing biodebris between the columns |
类型Ⅰ迷宫状微生物岩在切面上呈现出复杂的迷宫状构造,颜色为浅灰色至暗灰色,结构杂乱(图 3a,b)。迷宫体单独的形态可以呈现为短柱状、圆状、树形、新月形、迂曲状、环扣状和聚集状等形状,迷宫之间主要为灰泥所充填。这种类型的微生物岩类似于Lee et al. [5],Chen et al.[6]描述的板格状迷宫状微生物岩和Chen et al. [7]描述的泥质迷宫状生物岩,但最大的不同在于,迷宫体内部可见到清晰的中型尺度的凝块,甚至可见树形凝块(图 3c)。
类型Ⅱ和类型Ⅲ迷宫状微生物岩在垂向表现为分叉或会聚的柱状体,在横切面上则呈现出复杂的迷宫状构造(图 3d~g),颜色多为浅灰色(一般较周围沉积物颜色要浅)。两者的主要区别在于类型Ⅱ迷宫状微生物岩迷宫体之间的沉积物以灰泥为主,碳酸盐颗粒例如生物碎屑很少出现在类型Ⅱ迷宫状微生物岩的柱体内部,柱体往往表现为贫乏构造的均一石,或只含有少量生物碎屑(图 3h)。其次,在类型Ⅱ迷宫状微生物岩中,迷宫体与周围沉积物之间的边界除非被选择性白云石化作用所加强(图 3d),否则在很多情况下较难识别(图 3f),甚至难以识别[21]。而类型Ⅲ迷宫状微生物岩的迷宫体与周围沉积物之间的边界较为清晰(图 3e~h),柱体之间的沉积物以颗粒为主,并包含了三叶虫、双壳类的生物碎屑和少量的内碎屑,表明对于迷宫的形成灰泥的大量注入并非必须的[7]。
2.2 微观特征马沟剖面苗岭统的迷宫状微生物岩,在低倍显微镜下主要表现为非均匀分布的相对致密的泥晶、泥晶团块和斑斑点点不规则散布的微亮晶及亮晶;在高倍显微镜下可观察到较为典型的丝状蓝细菌鞘化石(图 4)。这些钙化蓝细菌丝状体直径约10~20 μm,稠密的泥晶方解石围绕菌丝体鞘选择性沉淀,形成明显的厚度均匀(1~5 μm)的暗色泥晶壁,其内部被亮晶或微亮晶方解石充填。钙化蓝细菌丝状体放大后可以观察到发育良好的管状构造(图 4b,e),菌丝体之间相互缠绕或不缠绕,可归为葛万菌及其组合(Girvanella Group)[8-10]。其中,蓝细菌丝状体和短柱状亮晶沉积物(很可能是钙化蓝细菌——胞网菌Bacinella的钙化残余物)在不规则泥晶团块和基质中的广泛出现支持了这些沉积物为蓝细菌微生物席的钙化产物的解释(图 4a,b),但单一的矿物相和泥晶为主的组构增加了进一步了解这一结构的难度,更何况要“穿越成岩作用过滤器”的影响[26]。

|
| 图 4 马沟剖面苗岭统徐庄组迷宫状微生物岩的微观特征 (a)(b)类型Ⅰ迷宫状微生物岩的镜下特征,放大后可见到清晰的缠绕状丝状蓝细菌鞘化石(葛万菌);(c)~(e)类型Ⅲ迷宫状微生物岩的镜下特征,可见到较多的生屑,以及普遍发育的泥晶构成的不规则状凝块,放大后可见到明显的钙化蓝细菌丝状体,小黑点为黄铁矿晶体残余物。所有照片都为单偏光镜照片 Fig.4 Photographs taken under plane polarized light showing microscopic characteristics and calcified cyanobacteria of maceriate microbialites from Miaolingian Xuzhuang Formation at the Magou section (a)(b) microscopic characteristics of TypeⅠ maceriate microbialites, consisting of cyanobacterial filaments. Note obvious intertwisted sheath fossils of the cyanobacterium Girvanella; (c)~(e) large number of bioclasts and irregular microclots of Type Ⅲ maceriate microbialites. The enlargement shows obvious calcified cyanobacteria filaments, and the black shapes are residual pyrite crystals |
迷宫状微生物岩中还普遍见到黄铁矿颗粒,大小为5~30 μm,多呈稀疏分布,局部集中出现,成为致密碳酸盐泥晶构成中的特别的矿物组成(图 4e)。尽管黄铁矿形成模式不尽相同,但主要都来源于硫酸盐还原细菌(SRB),且与硫酸盐还原细菌造成的硫酸盐还原反应存在着直接的成因联系[27-29]:硫化物主要来自于沉积硫化物(约占地球表面硫化物的95%以上),而由于Fe元素的大量富集,致使沉积物中的硫化物几乎完全以黄铁矿的形式产出[30]。弥散状分布的黄铁矿与纹层状、凝块状和钙化微生物及其残余物一起,反映了形成微生物碳酸盐岩过程中的微生物沉淀作用和早期石化作用[10-13, 28]。
3 均一石生物丘(层) 3.1 宏观特征均一石指那些相对贫乏构造,宏观组构(中型构造)呈现隐晶状态的微生物岩[12, 14, 25]。但自从定义以来,迄今为止还没有可以类比的现代均一石的典型实例[26],即便在古老的地层记录中也很少被描述。这或许跟均一石相对简单的宏观组构有关,但实际上,块状致密泥晶构成本身,就是一种特别的沉积构造[31](图 5)。

|
| 图 5 马沟剖面苗岭统徐庄组均一石生物丘和生物层 (a)均一石生物丘,徐庄组上部;(b)是(a)的局部特写,由致密泥晶构成;(c)远照,类型Ⅲ迷宫状微生物岩的地层向上过渡均一石生物层,徐庄组上部;(d)近照,类型Ⅲ迷宫状微生物岩的地层向上过渡均一石生物层,徐庄组上部;(e)是(d)中类型Ⅲ迷宫状微生物岩的特写;(f)是(d)中迷宫状微生物岩的特写,柱状体中可见一些生物碎屑,柱体之间为粗粒沉积物和生屑沉积 Fig.5 Leiolitic bioherm and biostromes of Miaolingian Xuzhuang Formation at Magou section (a) leiolitic bioherm in the upper Xuzhuang Formation; (b) enlargement of (a) showing a leiolite composed of dense micrites; (c) Distant view of part of the upper Xu-zhuang Formation showing Type Ⅲ maceriate microbialites transiting to leiolitic biostromes; (d) enlargement of part of upper Xuzhuang Formation showing Type Ⅲ maceriate microbialites transiting to leiolitic biostromes; (e) enlargement of Type Ⅲ maceriate microbialites in (d) showing biodebris and grains infilling between columns; (f) enlargement of leiolitic biostromes in (d) |
马沟剖面苗岭统均一石主要以生物丘(图 5a,b)或者生物层的形式产出(图 5c,d,e),主要位于SQ2高位体系域和强迫型海退体系域的下部,以及SQ3的高位体系域(SQ3第二个亚层序的顶部;图 2)。从SQ2高位体系域下部的类型Ⅰ迷宫状微生物岩,SQ2高位体系域上部的类型Ⅱ或类型Ⅲ迷宫状微生物岩,到SQ2强迫型海退体系域的均一石生物丘(层),再向上自然过渡为叠层石生物丘,构成了一个罕见的微生物岩序列(图 5c,d)。这一序列可能代表了从较厚的微生物席到较薄的微生物席,乃至微生物膜的变化[26-28]。
事实上,如前文所言,类型Ⅱ或类型Ⅲ迷宫状微生物岩中的柱状体大多也由不显构造的均一石构成。这些迷宫状微生物岩柱体跟柱体间填充物之间的界线清晰,反映了微生物席较强的构筑作用,也反映了微生物席中的碳酸盐来源以沉淀为主而不是捕获周围沉积物(图 5e,f)。SQ3第二个亚层序的顶部的均一石局部还见到冲刷面构造,说明这些块状泥晶在形成过程中曾受到过水流作用的改造,可能更多是受到较强的风暴流的破坏和改造[32],在冲刷面上多为微小颗粒质的沉积物所填充。
3.2 微观特征相比较为简单的宏观组构,均一石的微观组构跟其他类型的微生物碳酸盐岩一样,复杂且难以解释,并在暗色致密泥晶组构中,发育着较为典型的附枝菌等钙化蓝细菌化石(图 6)。尽管有一些证据表明均一石生物丘应该是蓝细菌微生物席复杂的钙化作用的产物[33],但泥晶占优的组分和微晶主导的结构掩盖了其一系列可能的起源[8, 26]。这些贫乏构造的块状泥晶灰岩跟其他岩性一起构成了河北唐县马家沟剖面寒武系苗岭统三级层序SQ2和SQ3在海退晚期的沉积,蕴藏着许多有待进一步发现的信息。

|
| 图 6 马沟剖面寒武系苗岭统徐庄组均一石中一些典型的钙化蓝细菌 (a)灌木丛状附枝菌菌落,可见棒状或球状附枝菌枝晶状生长;(b)房室状和扇状附枝菌菌落,枝晶较短,从内向外呈放射状;(c)房室状和簇球状附枝菌菌落,部分房室壁内的枝晶状结构不再明显,附枝菌与肾形菌的菌体就变得相似起来(左侧);(d)泥晶粗枝杈状附枝菌,横切面为树枝状,直径约为75 μm;(e)泥晶细枝杈状附枝菌,横切面为较细的树枝状,直径约为40 μm,包裹在亮晶或者微亮晶胶结物之中。所有照片都为单偏光镜照片 Fig.6 Photographs taken under plane polarized light showing typical calcified cyanobacteria in the Miaolingian Xuzhuang Formation at Magou section (a) bush-like Epiphyton composed of rod-shaped or micritic thalli with branching and dendritic growths; (b) chambered and fanshaped Epiphyton, with short dendrites radiating from inside to outside; (c) chambered and cystic Epiphyton, and the dendritic structure of some atrioventricular walls missing due to diagenesis, making it similar to Renalcis (left); (d) bifurcating rodlike branches (~75 µm) with occasional segmented branches; (e) Epiphyton with bifurcating fine branches (~40 µm) cemented by sparite |
附枝菌及其组合(Epithyton Group)是马沟剖面苗岭统均一石中比较常见的钙化蓝细菌化石,以明显的密而坚固的分枝为特征(图 6)。这些分枝由棒状或球粒状的致密泥晶所构成,菌落整体则呈房室状、扇形状和灌木丛状产出和发育。马沟剖面苗岭统均一石中附枝菌的分枝尺寸和菌落形态拥有广泛的变化,Woo et al.[34]所描述的山东张夏组中的4种附枝菌形态都有出现,大致可分为泥晶粗枝杈状、泥晶细枝杈状、分段枝杈状和管状枝杈等,微生物群落呈树枝状、房室状、簇球状、囊状等形态。当钙化作用和成岩作用抹掉房室壁的内部结构时,房室壁内的枝晶状结构不再明显,附枝菌与肾形菌的菌体形态就变得相似而难以区别(图 6c),以致有人提出过附枝菌和肾形菌(Renalcis)只不过是同一类钙化微生物生命循环中的不同阶段[35]。
附枝菌的生物亲和性至今还存在争议,由于其形态特征接近现代水体中的高级藻类而被解释为红藻[36-38],甚至归为异养生物等[39]。但目前大多数学者认为附枝菌更接近原核生物而被定义为一种蓝细菌群落[4, 34, 40-44]。一些特殊形态的附枝菌,菌落纵向呈房室状和扇形状,菌体呈明显放射状分叉的管丝状或囊状的蓝细菌又被命名为基座菌(Hedstroemia),其相邻的管有时候会共享隔膜或泥晶壁,可以类比于现代固氮的胶须菌(Rivulariaceae)[10, 45]或第谷菌(Tychonema)[46]。这些蓝细菌化石的保存很大程度上依靠早期钙化作用及成岩作用,成岩作用类型和过程的不同导致了钙化微生物与周围的胶结物之间的明显区别,也使附枝菌叶状体放射丛往往被包裹在亮晶或者微亮晶胶结物之中[34]。
4 叠层石 4.1 宏观特征马沟剖面苗岭统叠层石主要产出自SQ2强迫型海退体系域和SQ3的高位体系域,叠层石内部纹层都较粗糙,层理不十分清晰。按照其沉积组构特征大致可以分为3种类型(图 7)。类型Ⅰ为短柱状叠层石,为多次分叉(多为3次分叉)的短柱体,直径通常在1 cm之内,但分叉前柱体剧烈增宽,可为原来柱体的4倍,然后分为数个微散开的子柱体(图 7a,b)。柱体分叉频繁,横断面呈椭圆形,侧部可具有瘤状突起,从而使柱体边缘崎岖不平。当水流冲刷作用减弱时,柱体分叉可以跟旁边的柱体相融合,分布混乱,彼此之间难以区分(图 7a,b)。短柱状叠层石在地层中向上变为类型Ⅱ近水平缓波状叠层石,该叠层石同一个柱体的直径经常变化,时细时粗。柱体和柱体之间沉积物的界限不明显,内部纹层粗糙,层理不十分清晰(图 7c)。两类叠层石所在的地层顶部为一淹没不整合型层序界面,代表了较为典型的相对海平面下降期的沉积记录[24]。

|
| 图 7 马沟剖面寒武系苗岭统叠层石的宏观特征 (a)短柱状叠层石,多为多为3次分叉)的短柱体,分叉前柱体剧烈增宽,可为原来柱体的4倍,然后分为数个微散开的子柱体,徐庄组上部;(b)是(a)的素描,叠层石所处环境水动力逐渐减弱,下部由于水流强烈的冲刷作用,叠层石大多呈碎片状分布。向上冲刷作用减弱,柱体相互融合,柱体间沉积物粗粒物质也逐渐减少;(c)近水平缓波状叠层石,柱体的直径变化大,内部纹层粗糙,层理不清晰,徐庄组;(d)产出在鲕粒滩中的小型叠层石生物丘,张夏组;(e)是(d)中小型叠层石生物丘的放大;(f)是(e)中小型叠层石生物丘的特写,注意其粗糙的纹层和硬底构造 Fig.7 Fundamental features of stromatolites in the Cambrian Miaolingian Series at the Magou section (a) short-columnar stromatolites, mostly with three-branched short column which are distinctly widened (4× original width), then subdivided into several scattered sub-columns, Xuzhuang Formation; (b) sketch of (a), showing that the hydrodynamic force of the environment gradually decreased; (c) stromatolites with gentle wavy and blurred laminae with widely varying column diameters, Xuzhuang Formation; (d) small stromatolitic mounds produced in oolitic grain banks, Zhangxia Formation; (e) enlargement of (d) showing one of the small stromatolitic mounds; and (f) close-up of (e), note the coarse laminae and solid ground structure |
类型Ⅲ是位于SQ3的高位体系域的小型叠层石生物丘,生长在鲕粒滩之中(图 7d~f)。在硬地构造后,叠层石柱体之间的沉积物发生了质的变化,或许代表了一个水动力暂歇的一个阶段。在硬底构造之下,叠层石柱状形态反映了叠层石“高处更高”(the tall get taller)的生长机制[47-48],柱体之间的基质多为粗粒沉积,部分沉积颗粒被封闭在叠层石之中。局部可见冲刷面,表明较为强烈的风暴作用曾经破坏过叠层石的生长(图 7f)。而硬底之上的叠层石尽管也生长在鲕粒滩中,但叠层石柱体之间多为更加富集泥质物的碳酸盐泥晶所组成(图 7d~f),体现了构建叠层石的蓝细菌微生物席的选择性捕获作用,而鲕粒等主要再沉积的内碎屑等形式被外力(如风暴)卷入叠层石(图 7e,f)。叠层石在不到1 m的小型生物丘中能根据环境变化迅速做出调整,表明了构建叠层石的蓝细菌微生物席较高的适应能力和强烈的生命活性。
4.2 微观特征在显微镜下,短柱状叠层石可以观察到较好的微叠层构造,由较厚的富含有机质的泥晶层和较薄的亮晶或微亮晶层组成(图 8a,b),按照Schmid[49]的分类可定名为泥晶叠层石。叠层石纹层和柱体间填充物未出现分异,泥晶纹层厚度较为稳定,可解释为生物膜内微生物诱发的碳酸盐沉淀作用产物。缓波状叠层石拥有不规则或扭曲的纹层,其微叠层构造在早期钙化和后期成岩作用过程中遭受了破坏,不管是叠层石纹层还是纹层见的填充物中均有大量不均匀分布的石英粉砂(图 8c,d)。这些石英粉砂大小在5~15 μm之间,次棱角状至次圆状,分选良好,成分成熟度较高,表现出风成石英粉砂的特点[50],也说明了形成这类叠层石的微生物席较强的黏聚作用特征和分布于潮间带沉积环境[51]。尽管叠层石结构的形成是微生物席在表面增生并捕获沉积物所致,但马沟剖面苗岭统徐庄组短柱状叠层石和缓波状叠层石中并没有识别出较为特征的钙化微生物和蓝细菌化石(图 8a~d)。

|
| 图 8 马沟剖面寒武系苗岭统叠层石的微观特征 (a)(b)由较厚的富含有机质的泥晶层和较薄的亮晶或微亮晶层组成的微叠层构造,叠层石纹层和柱体间填充物未出现分异,徐庄组短柱状叠层石;(c)(d)不规则或扭曲的纹层,以及大量不均匀分布的石英粉砂,徐庄组缓波状叠层石;(e)~(h)叠层石中含少量鲕粒和生物碎屑,以及分布有大量不规则状的由蓝细菌丝状体构成的暗色泥晶凝块,放大以后可以见到丝状体; (h)是(g)中葛万菌的特写,张夏组小型叠层石生物丘。所有照片都为单偏光镜照片 (a)(b) micro-laminated structure consisting of thick organic-rich micritic layers and thin sparite layers without any differentiation between column and fillings, in short-columnar stromatolites of the Xuzhuang Formation; (c)(d) gentle wavy stromatolites with irregular or twisted laminates and a large amount of unevenly distributed quartz silt, Xuzhuang Formation; (e)~(h) stromatolites containing a small amount of oolitic grains and bioclasts and dark micritic clots of cyanobacterial filaments, the enlargement shows obvious filaments; and (h) is a close-up photography of Girvanella within(g), in small-scale stromatolite biomounds of Zhangxia Formation |
在马沟剖面苗岭统张夏组小型叠层石生物丘中,鲕粒和生物碎屑(如三叶虫碎片或骨刺)较为常见,更为特别的是分布有大量不规则状的暗色泥晶凝块(图 8e,f),类似杨仁超等[52]描述的“微生物团块”。在高倍镜下可观察到较为典型的葛万菌鞘化石:稠密的泥晶方解石围绕菌丝体鞘选择性沉淀,形成明显的厚度均匀(1~5 μm)的暗色泥晶壁,其内部被亮晶或微亮晶方解石充填(图 8g,h)。规则且均匀的泥晶壁说明了葛万菌丝管状体较早的钙化作用,至少要早于形成叠层石的微生物席的整体钙化时间,并最终以凝块的形式在叠层石中保存下来。
5 讨论顾名思义,微生物岩是在与微生物相关的作用下形成的岩石[10-14],具体而言是微生物席和微生物膜经过生物/有机矿化作用和石化/成岩作用的产物[10, 14, 26]。但截至目前,对形成微生物岩的生物,尤其是产生迷宫状构造的生物还知之甚少,使其成了名副其实的“生物学中沉睡的巨人”[53]。而且,微生物碳酸盐岩简单的矿物相(通常是单矿物相)往往掩盖了它们岩相学、化学性质和沉积过程的复杂性[28]。宏观上相对简单、呈隐晶状态,微观组构却复杂多样的均一石就是一个典型的实例。
此外,同一分类的微生物碳酸盐岩,可以形成于完全不同的环境[8-10, 54-55],例如叠层石几乎能够形成于任何水体中。南极冰川湖泊中、新西兰热水体系中、巴哈马台地和澳大利亚鲨鱼湾等热带、亚热带海域,都有现代叠层石的发现[48, 56-57],这使得用环境来解释微生物碳酸盐岩的成因或将微生物碳酸盐岩作为某种沉积环境的指示器都变得困难。尽管如此,同一时期同一盆地内沉积的微生物碳酸盐岩的类型和特征仍然具有一定规律性,代表着微生物与沉积物、水流之间的相互作用,其构造形态、宏微观组构和钙化作用特征反过来也可以反映和解释古地理环境[12-13],甚至具有展示生命与环境长周期历史的重要潜力[48](图 7, 9)。

|
| 图 9 马沟剖面寒武系苗岭统的沉积特征 (a)(b)钙化(蓝细菌)微生物席残余物中的黄铁矿(褐铁矿化),(b)是(a)在反射光下的现象,单偏光,徐庄组;(c)类型Ⅲ迷宫状微生物岩中的生物碎屑和由蓝细菌丝状体构成的凝块(白色箭头),徐庄组;(d)张夏组小型叠层石生物丘中大量生物碎屑和由蓝细菌丝状体构成的凝块(白色箭头);(e)均一石中“修饰”纹层,徐庄组;(f)干裂纹,徐庄组 Fig.9 Sedimentary phenomena of Miaolingian Series at the Magou section (a)(b)photograph taken under polarized light of pyrites (limonitization) within residual calcified microbial mat, Xuzhuang Formation; (b) as for (a), taken under reflected light; (c) bioclasts and calcified cyanobacteria in Type Ⅲ maceriate microbialites, Xuzhuang Formation; (d) large number of bioclasts and microclots consisting of calcified cyanobacteria within small-scale stromatolites, Zhangxia Formation; (e)'modified'laminae within leiolities, Xuzhuang Formation; and (f) desiccation cracks, Xuzhuang Formation |
在整个寒武纪和奥陶纪,蓝细菌在海相沉积物中都非常繁盛,而且也是构成微生物岩的最重要的微生物[7-13]。作为色素体的初创者,蓝细菌在藻类和植物进化中扮演着重要的角色[56]。尽管在现代正常环境中一般很少作为主要的沉积物,但在地质历史时期是一种可以大规模形成微生物碳酸盐岩的极为重要的微生物[10-13, 57-58]。蓝细菌通过光合作用吸收无机碳促进碳酸钙的沉淀,这种钙化作用会在微生物席中产生丝状微化石,而由蓝细菌所主导的微生物席最终会作为叠层石、凝块石等保存下来[8-13, 26]。马沟剖面寒武系苗岭统微生物岩中发育较为典型的钙化蓝细菌,如葛万菌(Girvanella)、附枝菌(Epithyton)和基座菌(Hedstroemia)等,也偶见介于上述5种蓝细菌之间的形态类型。以及如构建张夏组小型叠层石生物丘的微生物席表现出的强烈的生命活性和适应能力,都佐证了这些微生物岩是微生物席,尤其是蓝细菌所主导的微生物席的复杂的钙化作用的产物[51, 56-58]。
尽管如此,寒武纪第二世末期到奥陶纪早期发生“显生宙早期第一幕蓝细菌钙化作用事件”的具体机制仍是一个谜[8-13]。可能影响寒武纪及早奥陶统期间微生物岩(礁)发育的因素被归结为6种[59]:1)生物演化;2)板块运动;3)相对海平面变化;4)营养物的可利用性;5)古气候;6)海水化学性质的强制改变,这些因素在一定程度上都可能影响了生物岩(礁)的建造。Riding[8-13]认为蓝细菌钙化作用事件(CCEs,Cyanobacterial Calcification Events)这可能与大气圈二氧化碳的浓度、全球气候(高温可以提高碳酸盐沉淀率)以及来自分层盆地碱泵的产生等有着密切的联系。
马沟剖面寒武系苗岭统微生物岩表明微生物进行的捕获和黏结作用过程可能并不占主导,促使酸钙沉淀的更重要的原因是有机矿化作用,包含生物诱导的矿化和生物影响的矿化两个过程[60]。生物诱导的有机矿化作用中,最重要的机制是蓝细菌等的光合作用和硫酸盐还原细菌(Sulfate-Reducing Bacteria,SRB)的硫酸盐还原反应导致的“碱度发动机”:在微生物席顶部含氧区域,蓝细菌等通过光合作用吸收CO2和/或HCO3-增加了周围环境的碱度,进而促进了碳酸盐的沉淀[61];在微生物席下部的贫氧层和/或缺氧层,硫酸盐还原菌通过硫酸盐还原反应消耗了SO42-和有机碳化合物,增加了微环境的碱度,促进了碳酸盐的沉淀[28, 62]。生物影响的有机矿化作用主要发生在微生物席的底部,其中EPS的降解和钙化作用最为重要。微生物席的这部分环境更为封闭,EPS被硫酸盐还原细菌、产甲烷细菌等“终极”细菌降解后为碳酸盐矿物的沉淀提供成核中心,同时将吸附的大量Mg2+等释放,影响微环境的地化性质,进而形成矿化作用的场所,促进了碳酸盐的沉淀[28, 60]。而较为普遍的黄铁矿晶体的残余物说明了形成这些均一石的微生物沉淀作用和早期石化作用与硫酸盐还原反应存在着成因联系(图 9a,b)。
从沉积环境和层序位置来看,马家沟剖面寒武系苗岭统微生物岩既可以呈现为简单的丘状、复合丘状,也可以持续延展形成生物层。但不论那种形态,都出现在高位体系域或强迫型海退阶段。随着海退的进行和碳酸盐的加积及进积作用过程,苗岭统微生物岩大致演变为:类型Ⅰ迷宫状微生物岩—类型Ⅱ迷宫状微生物岩—类型Ⅲ迷宫状微生物岩—均一石生物丘(层)—短柱状(指状)叠层石—缓波状叠层石(图 10)。而产出在苗岭统张夏组高能鲕粒滩中的分米至米级别的小型叠层石生物丘,则是一种特别的类型。

|
| 图 10 马沟剖面寒武系苗岭统微生物碳酸盐岩的沉积模式示意图 Fig.10 Depositional model of microbial carbonates for Miaolingian Series at Magou section |
微生物岩与各种生物化石的同时出现表明微生物岩形成于生物繁盛的正常盐度的环境[34]。迷宫状微生物岩主要发育在SQ2的高位体系域中,其中类型Ⅰ迷宫状微生物岩呈生物丘或者生物层的样式产出,迷宫体内部结构杂乱,呈凝块状或树形;迷宫之间的沉积物灰泥为主,包含有少量生屑;潮上带特征完全没有出现(如干燥开裂,窗格孔洞和帐篷结构等沉积构造)等表明类型Ⅰ迷宫状微生物岩应该形成于正常浪基面以下的潮下带低能环境。类型Ⅱ和类型Ⅲ迷宫状微生物岩之间平滑过渡,没有截然的界限,形成于潮下带上部至潮间带的中—高能环境。但类型Ⅱ迷宫状微生物岩的柱体大多为贫乏层理的柱状,柱体之间以灰泥为主;而Ⅲ迷宫状微生物岩柱体之间更多的粗粒沉积物,杂乱的迷宫边缘和柱体中更多的颗粒、生屑含量综合表明类型Ⅲ迷宫状微生物岩形成环境的能量更高(图 3)。
虽然均一石以相对贫乏构造为特征[25-27],但是也反映出与水流之间的相互作用而表现为至少是中等能量的,风暴浪基面之上的浅水环境中的产物[32]。而马沟剖面更浅,大致在潮间带环境,与潮间短柱状叠层石和潮上带的近水平缓波状叠层石一起构成了SQ2的强迫型海退体系域。潮间带由于经常暴露地表和发生脱水作用,导致均一石的“均一”结构受扰,表面会出现一些“修饰”纹层(图 9e)。在均一石和叠层石所在地层,也可见到典型的干裂纹(图 9f)。短柱状叠层石和近水平缓波状叠层石粗糙、杂乱的纹层,干裂纹和角砾化的倾向也反应其形成的水体很浅(图 7a~c)。
发育在SQ3的高位体系域鲕粒滩中的小型叠层石生物丘是微生物席广泛的适应能力和强烈的生命活性的体现[35](图 7d~f)。用微生物碳酸盐岩的沉积构造和组构进行环境解释时,不得不面对它的形态和组构是受环境控制还是受微生物群落控制的问题。长期以来,都把叠层石的形态特征与水动力条件相联系,一般认为近水平缓波状叠层石往往在水动力弱的潮上带形成,缓波状叠层石主要分布于水动力弱的潮间带,而锥状和半球状叠层石则形成于水动力很强的潮间带[47-49]。但实际情况要复杂得多,最近在南极常年被冰覆盖的Untersee湖中就发现了小尖塔状叠层石和大型锥状叠层石,并认识到生物学行为是造成这种泥质叠层石形态学特征的主要起因[63]。张夏组鲕粒滩中即便不到1 m的小型叠层石生物丘,在硬底构造之上的部分和之下部分叠层石的形态的迅速变化(图 7d~f),体现了微生物群落对叠层石建造的制约性、主导性和适应性。目前来看,叠层石内部柱体的分叉、纹层的特征和微构造是各种因素共同作用的结果,究竟那种作用占主导地位仍需进一步研究。
6 结论(1)华北地台寒武系苗岭统包括毛庄组、徐庄组、张夏组和崮山组,分别构成4个三级层序即SQ1至SQ4。这些三级层序由米级旋回有序叠加形成的各具特色的沉积相序列所构成,反映了海平面的升降变化及沉积学响应。其中,以发育在SQ2的高位体系域和强迫型海退体系域,SQ3和SQ4的高位体系域中构成的生物层、简单的生物丘或生物丘复合体的迷宫微生物岩、均一石和叠层石最为特别。
(2)华北地台寒武系苗岭统微生物岩的种类和沉积环境包括形成于正常浪基面以下、潮下带低能环境的类型Ⅰ迷宫状微生物,潮下带上部至潮间带的中—高能环境的类型Ⅱ和类型Ⅲ迷宫状微生物岩,风暴浪基面之上的浅水环境中的均一石,潮间带短柱状叠层石和潮上带的近水平缓波状叠层石,以及高能鲕粒滩中的小型叠层石生物丘,从迷宫状微生物岩到均一石,再到叠层石可能代表了从较厚的微生物席到较薄的微生物膜的变化序列。
(3)无论是在迷宫状微生物岩、均一石中,还是在叠层石中,都见到了一种或多种钙化蓝细菌化石,如葛万菌(Girvanella)、附枝菌(Epithyton)和基座菌(Hedstroemia)等,也偶见介于上述蓝细菌之间的形态类型,表明这些微生物岩是由蓝细菌所主导的微生物席的复杂的钙化作用的产物,而大量呈弥散状分布的黄铁矿晶体或颗粒则表明硫酸盐还原菌等非光合作用细菌和异养细菌可能在促进碳酸盐沉淀过程中扮演了重要角色,这些现象为理解微生物岩复杂多样的沉积作用和形成机制提供了重要线索。
| [1] |
Wood R. The ecological evolution of reefs[J]. Annual Review of Ecology and Systematics, 1998, 29: 179-206. |
| [2] |
Lee J H, Chen J T, Choh S J, et al. Furongian(Late Cambrian) sponge-microbial maze-like reefs in the North China Platform[J]. Palaios, 2014, 29(1): 27-37. |
| [3] |
Hong J, Cho S H, Choh S J, et al. Middle Cambrian siliceous sponge-calcimicrobe buildups(Daegi Formation, Korea):metazoan buildup constituents in the aftermath of the Early Cambrian extinction event[J]. Sedimentary Geology, 2012, 253-254: 47-57. |
| [4] |
Riding R. Microbial carbonate abundance compared with fluctuations in metazoan diversity over geological time[J]. Sedimentary Geology, 2006, 185(3/4): 229-238. |
| [5] |
Lee J H, Chen J T, Chough S K. Paleoenvironmental implications of an extensive maceriate microbialite bed in the Furongian Chaomidian Formation, Shandong Province, China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2010, 297(3/4): 621-632. |
| [6] |
Chen J T, Chough S K, Lee J H, et al. Sequence-stratigraphic comparison of the upper Cambrian Series 3 to Furongian succession between the Shandong region, China and the Taebaek area, Korea:high variability of bounding surfaces in an epeiric platform[J]. Geosciences Journal, 2012, 16(4): 357-379. |
| [7] |
Chen J T, Lee J H, Woo J. Formative mechanisms, depositional processes, and geological implications of Furongian(late Cambrian)reefs in the North China Platform[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 414: 246-259. |
| [8] |
Riding R. Microbial carbonates:the geological record of calcified bacterial-algal mats and biofilms[J]. Sedimentology, 2000, 47(S1): 179-214. |
| [9] |
Riding R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition[J]. Geobiology, 2006, 4(4): 299-316. |
| [10] |
Riding R. Structure and composition of organic reefs and carbonate mud mounds:concepts and categories[J]. Earth Science Reviews, 2002, 58(1): 163-231. |
| [11] |
Riding R. Microbialites, stromatolites, and thrombolites[M]//Reitner J, Thiel V. Encyclopedia of geobiology. Dordrecht: Springer, 2011: 635-654.
|
| [12] |
Riding R. Calcified cyanobacteria[M]//Reitner J, Thiel V. Encyclopedia of geobiology. Heidelberg: Springer, 2011: 211-223.
|
| [13] |
Riding R. A hard life for cyanobacteria[J]. Science, 2012, 336(6080): 427-428. |
| [14] |
王龙, Latif K, Riaz M, 等. 微生物碳酸盐岩的成因、分类以及问题与展望:来自华北地台寒武系微生物碳酸盐岩研究的启示[J]. 地球科学进展, 2018, 33(10): 1005-1023. [Wang Long, Latif K, Riaz M, et al. The genesis, classification, problems and prospects of microbial carbonates:Implications from the Cambrian carbonate of North China Platform[J]. Advances in Earth Science, 2018, 33(10): 1005-1023.] |
| [15] |
Peters S E, Gaines R R. Formation of the 'Great Unconformity' as a trigger for the Cambrian explosion[J]. Nature, 2012, 484(7394): 363-366. |
| [16] |
冯增昭, 彭勇民, 金振奎, 等. 中国寒武纪和奥陶纪岩相古地理[M]. 北京: 石油工业出版社, 2004: 1-233. [Feng Zengzhao, Peng Yongmin, Jin Zhenkui, et al. Lithofacies paleogeography of the Cambrian and Ordovician in China[M]. Beijing: Petroleum Industry Press, 2004: 1-233.]
|
| [17] |
王龙, 吴海, 张瑞, 等. 碳酸盐台地的类型、特征和沉积模式:兼论华北地台寒武纪陆表海-淹没台地的沉积样式[J]. 地质论评, 2018, 64(1): 62-76. [Wang Long, Wu Hai, Zhang Rui, et al. The types, characteristics and depositional models of carbonate platform:Implications for Cambrian sedimentary patterns of epeiric-drowned carbonate platform in North China[J]. Geological Review, 2018, 64(1): 62-76.] |
| [18] |
韩作振, 陈吉涛, 张晓蕾, 等. 鲁西寒武系第三统张夏组附枝菌与附枝菌微生物灰岩特征研究[J]. 地质学报, 2009, 83(8): 1097-1103. [Han Zuozhen, Chen Jitao, Zhang Xiaolei, et al. Characteristics of epiphyton and epiphyton microbialites in the Zhangxia Formation (Third Series of Cambrian), Shandong province[J]. Acta Geologica Sinica, 2009, 83(8): 1097-1103.] |
| [19] |
彭善池, 赵元龙. 全球寒武系第三统和第五阶"金钉子"正式落户我国[J]. 地层学杂志, 2018, 42(3): 325-327. [Peng Shanchi, Zhao Yuanlong. The proposed global standard stratotypesection and point(GSSP)for the conterminous base of Miaolingian Series and Wuliuan Stage at Balang, Jianhe, Guizhou, China was ratified by IUGS[J]. Journal of Stratigraphy, 2018, 42(3): 325-327.] |
| [20] |
马永生, 梅冥相, 周润轩, 等. 层序地层框架下的鲕粒滩形成样式:以北京西郊下苇甸剖面寒武系第三统为例[J]. 岩石学报, 2017, 33(4): 1021-1036. [Ma Yongsheng, Mei Mingxiang, Zhou Runxuan, et al. Forming patterns for the oolitic bank within the sequence-stratigraphic framework:An example from the Cambrian Series 3 at the Xiaweidian section in the western suburb of Beijing[J]. Acta Petrologica Sinica, 2017, 33(4): 1021-1036.] |
| [21] |
Aitken J D. Classification and environmental significance of cryptalgal limestones and dolomites, with illustrations from the Cambrian and Ordovician of southwestern Alberta[J]. Journal of Sedimentary Research, 1967, 37(4): 1163-1178. |
| [22] |
Shapiro R S, Awramik S M. Favosamaceria cooperi new group and form:a widely dispersed, time-restricted thrombolite[J]. Journal of Paleontology, 2006, 80(3): 411-422. |
| [23] |
Shapiro R S. A comment on the systematic confusion of thrombolites[J]. Palaios, 2000, 15(2): 166-169. |
| [24] |
梅冥相, 张瑞, 李屹尧, 等. 华北地台东北缘寒武系芙蓉统叠层石生物丘中的钙化蓝细菌[J]. 岩石学报, 2017, 33(4): 1073-1093. [Mei Mingxiang, Zhang Rui, Li Qiyao, et al. Calcified cyanobacterias within the stromatolotic bioherm for the Cambrian Furongian Series in the northeastern margin of the North-China Platform[J]. Acta Petrologica Sinica, 2017, 33(4): 1073-1093.] |
| [25] |
Braga J C, Martin J M, Riding R. Controls on microbial dome fabric development along a carbonate-siliciclastic shelf-basin transect, Miocene, SE Spain[J]. Palaios, 1995, 10(4): 347-361. |
| [26] |
Dupraz C, Reid R P, Visscher P T. Microbialites, modern[M]//Reitner J, Thiel V. Encyclopedia of geobiology. Dordrecht: Springer, 2011: 617-635.
|
| [27] |
Schieber J. Sedimentary pyrite:a window into the microbial past[J]. Geology, 2002, 30(6): 531-534. |
| [28] |
Dupraz C, Reid R P, Braissant O, et al. Processes of carbonate precipitation in modern microbial mats[J]. Earth-Science Reviews, 2009, 96(3): 141-162. |
| [29] |
Bontognali T R R, McKenzie J A, Warthmann R J, et al. Microbially influenced formation of Mg-calcite and Ca-dolomite in the presence of exopolymeric substances produced by sulphatereducing bacteria[J]. Terra Nova, 2014, 26(1): 72-77. |
| [30] |
Rickard D, Mussmann M, Steadman J A. Sedimentary sulfides[J]. Elements, 2017, 13(2): 117-122. |
| [31] |
钟建华, 梁刚. 沉积构造的研究现状及发展趋势[J]. 地质论评, 2009, 55(6): 831-839. [Zhong Jianhua, Liang Gang. Situation of study and development tendency of sedimentary structure[J]. Geological Review, 2009, 55(6): 831-839.] |
| [32] |
Myrow P M, Southard J B. Tempestite deposition[J]. Journal of Sedimentary Research, 1996, 66(5): 875-887. |
| [33] |
Xiao E Z, Latif K, Riaz M, et al. Calcified microorganisms bloom in Furongian of the North China Platform:evidence from Microbialitic-Bioherm in Qijiayu Section, Hebei[J]. Open Geosciences, 2018, 10(1): 250-260. |
| [34] |
Woo J, Chough S K, Han Z. Chambers of epiphyton thalli in microbial buildups, Zhangxia Formation(Middle Cambrian), Shandong Province, China[J]. Palaios, 2008, 23(1): 55-64. |
| [35] |
Luchinina V A. Remalcis and Epiphyton as different stages in the life cycle of calcareous algae[J]. Paleontological Journal, 2009, 43(4): 463-468. |
| [36] |
Woo J, Chough S K. Growth patterns of the Cambrian microbialite:Phototropism and speciation of Epiphyton[J]. Sedimentary Geology, 2010, 229(1): 1-8. |
| [37] |
Luchinina V A, Terleev A A. The morphology of the genus Epiphyton Bornemann[J]. Geologia Croatica, 2008, 61(2/3): 105-111. |
| [38] |
Adachi N, Ezaki Y, Liu J B. The late early Cambrian microbial reefs immediately after the demise of archaeocyathan reefs, Hunan Province, South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 407: 45-55. |
| [39] |
Săsăran E, Bucur I I, Pleş G, et al. Late Jurassic Epiphytonlike cyanobacteria:indicators of long-term episodic variation in marine bioinduced microbial calcification?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 401: 122-131. |
| [40] |
刘丽静, 杨海军, 潘文庆, 等. 中国新疆塔里木盆地上奥陶统良里塔格组的钙藻化石[J]. 微体古生物学报, 2012, 29(1): 18-38. [Liu Lijing, Yang Haijun, Pan Wenqing, et al. Calcareous algae from the Upper Ordovician Lianglitage Formation in the Tarim Basin, Xinjiang, China[J]. Acta Micropalaeontologica Sinica, 2012, 29(1): 18-38.] |
| [41] |
Laval B, Cady S L, Pollack J C, et al. Modern freshwater microbialite analogues for ancient dendritic reef structures[J]. Nature, 2000, 407(6804): 626-629. |
| [42] |
代明月, 齐永安, 常玉光, 等. 河南登封地区寒武系第三统馒头组二段中的核形石及其意义[J]. 沉积学报, 2014, 32(3): 410-417. [Dai Mingyue, Qi Yong'an, Chang Yuguang, et al. Oncoids and their significance from the second member of the Mantou Formation(Cambrian Series 3), Dengfeng area, Henan[J]. Acta Sedimentologica Sinica, 2014, 32(3): 410-417.] |
| [43] |
Adachi N, Kotani A, Ezaki Y, et al. Cambrian series 3 lithistid sponge-microbial reefs in Shandong Province, North China:reef development after the disappearance of archaeocyaths[J]. Lethaia, 2015, 48(3): 405-416. |
| [44] |
罗平, 王石, 李朋威, 等. 微生物碳酸盐岩油气储层研究现状与展望[J]. 沉积学报, 2013, 31(5): 807-823. [Luo Ping, Wang Shi, Li Pengwei, et al. Review and prospectives of microbial carbonate reservoirs[J]. Acta Sedimentologica Sinica, 2013, 31(5): 807-823.] |
| [45] |
Liu L J, Wu Y S, Jiang H X, et al. Paleoenvironmental distribution of ordovician calcimicrobial associations in the Tarim Basin, Northwest China[J]. Palaios, 2017, 32(7): 462-489. |
| [46] |
de los Ríos A, Ascaso C, Wierzchos J, et al. Microstructure and cyanobacterial composition of microbial mats from the High Arctic[J]. Biodiversity and Conservation, 2015, 24(4): 841-863. |
| [47] |
曹瑞骥, 袁训来. 中国叠层石研究的历史和现状[J]. 微体古生物学报, 2003, 20(1): 5-14. [Cao Ruiji, Yuan Xunlai. Brief history and current status of stromatolite study in China[J]. Acta Micropalaeontologica Sinica, 2003, 20(1): 5-14.] |
| [48] |
Bosak T, Knoll A H, Petroff A P. The meaning of stromatolites[J]. Annual Review of Earth and Planetary Sciences, 2013, 41: 21-44. |
| [49] |
Schmid D U. Marine mikrobolithe und mikroinkrustierer aus dem oberjura[J]. Profil, 1996, 9: 101-251. |
| [50] |
Marshall J R, Bull P A, Morgan R M. Energy regimes for aeolian sand grain surface textures[J]. Sedimentary Geology, 2012, 253-254: 17-24. |
| [51] |
常玉光, 齐永安, 郑伟, 等. 叠层石微生物席生态系研究进展[J]. 河南理工大学学报(自然科学版), 2013, 32(3): 356-364. [Chang Yuguang, Qi Yong'an, Zheng Wei, et al. Research progress on microbial mat ecosystem of stromatolites[J]. Journal of Henan Polytechnic University(Natural Science), 2013, 32(3): 356-364.] |
| [52] |
杨仁超, 樊爱萍, 韩作振, 等. 山东寒武系毛庄阶微生物团块的形态特征与成因[J]. 中国科学(D辑):地球科学, 2013, 43(3): 423-432. [Yang Renchao, Fan Aiping, Han Zuozhen, et al. Characteristics and genesis of microbial lumps in the Maozhuang Stage(Cambrian Series 2), Shandong province, China[J]. Science China(Seri. D):Earth Sciences, 2013, 43(3): 423-432.] |
| [53] |
Stolz J F, Reid R P, Visscher P T, et al. The microbial communities of the modern marine stromatolites at Highborne Cay, Bahamas[J]. Atoll Research Bulletin, 2009, 567: 1-29. |
| [54] |
齐永安, 孙晓芳, 代明月, 等. 豫西鲁山寒武系馒头组微生物岩旋回及其演化[J]. 微体古生物学报, 2017, 34(2): 170-178. [Qi Yong'an, Sun Xiaofang, Dai Mingyue, et al. The microbialite cycles and their evolution from the Cambrian Mantou Formation, Lushan, western Henan province, central China[J]. Acta Micropalaeontologica Sinica, 2017, 34(2): 170-178.] |
| [55] |
倪新锋, 沈安江, 韦东晓, 等. 碳酸盐岩沉积学研究热点与进展:AAPG百年纪念暨2017年会及展览综述[J]. 天然气地球科学, 2018, 29(5): 729-742. [Ni Xinfeng, Shen Anjiang, Wei Dongxiao, et al. Current hot topics and advances of carbonate sedimentology:AAPG 100 anniversary and 2017 annual meeting and exhibition[J]. Natural Gas Geoscience, 2018, 29(5): 729-742.] |
| [56] |
Raven J A. The Evolution of cyanobacterial symbioses[J]. Biology and Environment:Proceedings of the Royal Irish Academy, 2002, 102B(1): 3-6. |
| [57] |
Jones B, Renaut R W, Konhauser K O. Genesis of large siliceous stromatolites at Frying Pan Lake, Waimangu geothermal field, North Island, New Zealand[J]. Sedimentology, 2005, 52(6): 1229-1252. |
| [58] |
Stinchcomb B L, Angeli N A. New Cambrian and lower Ordovician monoplacophorans from the Ozark uplift, Missouri[J]. Journal of Paleontology, 2002, 76(6): 965-974. |
| [59] |
Rowland S M, Shapiro R S. Reef patterns and environmental influences in the Cambrian and earliest Ordovician[M]//Kiessling W, Flügel E, Golonka J. Phanerozoic reef patterns. Tulsa: SEPM Special Publication, 2002: 95-128.
|
| [60] |
Gerdes G. What are microbial mats?[M]//Seckbach J, Oren A. Microbial mats. Dordrecht: Springer, 2010: 3-25.
|
| [61] |
Arp G, Reimer A, Reitner J. Photosynthesis-induced biofilm calcification and calcium concentrations in Phanerozoic oceans[J]. Science, 2001, 292(5522): 1701-1704. |
| [62] |
Braissant O, Decho A W, Dupraz C, et al. Exopolymeric substances of sulfate-reducing bacteria:interactions with calcium at alkaline PH and implication for formation of carbonate minerals[J]. Geobiology, 2010, 5(4): 401-411. |
| [63] |
Andersen D T, Sumner D Y, Hawes I, et al. Discovery of large conical stromatolites in Lake Untersee, Antarctica[J]. Geobiology, 2011, 9(3): 280-293. |