 2016, Vol. 34
2016, Vol. 34扩展功能
文章信息
- 韦雪梅, 韦恒叶, 邱振
- WEI XueMei, WEI HengYe, QIU Zhen
- 瓜德鲁普末期生物灭绝是由峨眉山大火成岩省(LIP)引起的吗?
- Was the End-Guadalupian Mass Extinction Caused by the Emeishan LIP Eruption?
- 沉积学报, 2016, 34(3): 436-451
- ACTA SEDIMENTOLOGICA SINCA, 2016, 34(3): 436-451
- 10.14027/j.cnki.cjxb.2016.03.002
-
文章历史
- 收稿日期: 2015-05-04
- 收修改稿日期: 2015-10-15



2. 中国石油勘探开发研究院 北京 100083
2. Petrochina Research Institute of Petroleum Exploration and Development, Beijing 100083, China
瓜德鲁普统末期是地球演化历史中重大转折时期,浅海生物面貌、全球海平面、板块运动以及地球内部动力(如地磁极性及火山作用)均发生重大的转变。瓜德鲁普末期(部分学者称为卡匹敦中期生物灭绝事件[1,2])生物灭绝事件是晚古生代末期生物大灭绝事件的第一幕[3, 4]。但关于该次生物灭绝的原因不清楚,前人提出了各种成因机制,包括峨眉山大火成岩省(LIP)的喷发[5, 6, 7]、全球海平面下降[8, 9, 10, 11]、卡穆拉(Kamura)变冷事件[11, 12]以及深海缺氧[12, 13]。由于放射性同位素及生物地层数据均显示峨眉山LIP喷发与瓜德鲁普末期生物灭绝的时间一致[7, 14, 15, 16, 17],部分学者认为两者存在某种成因联系[14, 18, 19]。前人研究表明峨眉山LIP的喷发引起大陆特别是海洋环境的恶化,加大生物生存压力,从而引起生物灭绝事件。火山喷发引起的环境扰动在碳同位素地球化学记录中保存较好,Wignall et al.[9]通过研究华南贵州省熊家场剖面发现峨眉山玄武岩的喷发与碳酸盐岩碳同位素负偏及生物灭绝在时间上存在吻合,从而明确提出峨眉山LIP是导致瓜达鲁普末期生物灭绝的主要因素。近年来,一些年轻学者给出新解,如Yan et al.[20]认为碳同位素负偏是由于当时海洋初级生产力下降引起的;Saitoh et al.[13]认为朝天剖面的G-L界线处碳同位素负偏是由于当时上升洋流带来富12C的底部冷水混入海洋表层海水所致;而Jost et al.[21]甚至认为部分著名剖面(如铁桥剖面、朝天剖面)在G-L界线处的同位素负偏是由于成岩作用引起的,不代表原始的海水溶解无机碳同位素信号。瓜德鲁普统—乐平统(G-L)界线生物灭绝事件的成因争论再起。为更好地弄清楚G-L界线生物灭绝的原因,本文试图在全面分析生物灭绝步调及峨眉山玄武岩发喷过程的基础上,从能反映气候与构造活动规模的碳同位素与锶同位素有机结合分析的角度出发,深入剖析生物灭绝与构造活动、气候剧变的协同关系,探讨两者之间的因果关系。
1 瓜德鲁普末期生物灭绝事件地质历史时期生物灭绝事件表现为短期内超过50%的生物种类快速灭绝。在整个显生宙的化石记录中,存在5次大的生物灭绝事件[22,23]。其中晚古生代末期生物大灭绝是最为严重的一次生物危机事件,其导致全球的生态系统遭受了到前所未有的浩劫。该灭绝事件在时间上分为两幕[3,4],第一幕发生于259.1 Ma的瓜德鲁普统—乐平统界线(中—上二叠统);第二幕发生于252.18 Ma[24]的二叠—三叠系(P-T)界线。G-L界线生物危机是一个独立的生物灭绝事件,且从G-L界线开始,地球内部动力从长期的稳定开始变得频繁的不稳定,石炭至二叠纪长期稳定的环境与气候开始出现扰动,该生物灭绝事件比P-T界线生物灭绝事件更为重要[25]。
G-L界线生物灭绝事件最先被发现于浅海有孔虫的更替,特别是 (竹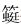 )类有孔虫的灭绝[3,4]。在随后的研究中发现,其他的浅海无脊椎动物如腕足、珊瑚、双壳、腹足、菊石和苔藓虫[19, 25, 26, 27, 28, 29]以及浅海植物钙质藻类[7, 30, 31]在该次生物灭绝事件中同样遭受重创(图 1)。类有孔虫67%的属在该次生物灭绝事件中消失[32],构造复杂的大型类,尤其是具有蜂窝层者包括Neoschwagerinidae科、Schwagerinidae科及Verbeekinidae科在晚瓜德鲁普世的卡匹敦期(Capitanian)大部分消失殆尽[33,34,35]。其中,Neoschwagerinidae和Verbeekinidae科的所有生物种全部灭绝[36]。取而代之的是常见于G-L过渡期的构造较为简单、较小的Ozawainellidae科和Schubertellidae科,如Reichelina属和Codonofusiella属[1, 35]。相比 类有孔虫,非类小型有孔虫(尤其是瓶虫超科)受到的影响较少[30, 31, 37],但仍然有37%的属级灭绝[32]。在该生物危机事件中,类和非类有孔虫的灭绝开始于卡匹敦中期的Jinogondolella altudaensis牙形刺带[7];Shen和Shi[38]通过广西来宾GSSP剖面的 类有孔虫研究则认为 类有孔虫的灭绝出现在Jinogondolella granti牙形刺带(图 1),灭绝时间更晚,但均属于卡匹顿的中晚期。
)类有孔虫的灭绝[3,4]。在随后的研究中发现,其他的浅海无脊椎动物如腕足、珊瑚、双壳、腹足、菊石和苔藓虫[19, 25, 26, 27, 28, 29]以及浅海植物钙质藻类[7, 30, 31]在该次生物灭绝事件中同样遭受重创(图 1)。类有孔虫67%的属在该次生物灭绝事件中消失[32],构造复杂的大型类,尤其是具有蜂窝层者包括Neoschwagerinidae科、Schwagerinidae科及Verbeekinidae科在晚瓜德鲁普世的卡匹敦期(Capitanian)大部分消失殆尽[33,34,35]。其中,Neoschwagerinidae和Verbeekinidae科的所有生物种全部灭绝[36]。取而代之的是常见于G-L过渡期的构造较为简单、较小的Ozawainellidae科和Schubertellidae科,如Reichelina属和Codonofusiella属[1, 35]。相比 类有孔虫,非类小型有孔虫(尤其是瓶虫超科)受到的影响较少[30, 31, 37],但仍然有37%的属级灭绝[32]。在该生物危机事件中,类和非类有孔虫的灭绝开始于卡匹敦中期的Jinogondolella altudaensis牙形刺带[7];Shen和Shi[38]通过广西来宾GSSP剖面的 类有孔虫研究则认为 类有孔虫的灭绝出现在Jinogondolella granti牙形刺带(图 1),灭绝时间更晚,但均属于卡匹顿的中晚期。

|
| 图 1 G-L界线牙形刺生物带及其相应的生物灭绝事件、构造及海平面升降事件图 牙形刺带据Jin et al.[48],生物灭绝事件据Shen and Shi[38],年代界线据Wignall et al.[9]和Zhong et al.[17],海平面变化据Chen et al.[49] Fig. 1 The diagram of conodont biozones,extinction horizons of fossil groups,Emeishan basalt eruption and sea level change across the G-L boundary conodont biozones data is from Jin et al.[48]; mass extinction events data is from Shen and Shi[38]; age-constraint according to Wignall et al.[9] and Zhong et al.[17]; sea level change is from Chen et al.[49] |
相比有孔虫,腕足类在该事件中虽然也存在灭绝[26, 38, 39],但灭绝率较低[40],属级的灭绝率只有30%[41],尽管种级别的灭绝率高达87%[42]。一些具茅口期特征的属如Cryptospirifer,Kiangsiella,Urushtenoidea和Vediproductus消失,导致吴家坪早期的腕足类分异度明显降低[41],然而乐平世繁盛的属种如Edriosteges,Gubleria,Peltichia,Strophalosiina及Tyloplecta yangtzeensis的出现,使得吴家坪晚期腕足类的分异度很快得到恢复[39, 42]。在该次生物危机事件中,腕足类属级的更替开始于卡匹敦期的Jinogondolella xuanhanensis牙形刺带[38](图 1)。
珊瑚78%的属和75%的科在该事件中消失[19]。其中,皱壁珊瑚和横板珊瑚在种级别的灭绝率分别高达80%和99%[19]。Wang和Sugiyama[19]认为,皱壁珊瑚灭绝于卡匹敦期的Jinogondolella postserrata牙形刺带,而Shen和Shi[38]认为皱壁珊瑚发生转变发生于Jinogondolella postserrata至Jinogondolella xuanhanensis牙形刺带,灭绝于J. xuanhanensis带。表明珊瑚的灭绝比其他无脊椎动物的灭绝较早,消失于卡匹敦早中期。可长达1 m的大型双壳Alatoconchidae,其灭绝于卡匹敦晚期的Jinogondolella altudaensis至Jinogondolella prexuanhanensis牙形刺带[43,44]。腹足类的灭绝率尚存在争议,杨遵义等[45]认为62个属中有60个属消失,Erwin和Pan[46]及潘华璋[47]认为腹足在该时期灭绝的属高达60%,而Knoll et al.[27]认为腹足类属的灭绝率仅为42%。由于中二叠世腹足类的研究工作不够,其精确的灭绝时间尚未清楚,但其灭绝时间可确定为卡匹敦期[47]。
菊石类在该次事件中也遭受了影响[45, 50],如Shouchangoceratidae、Paragastrioceratidae及Cyclolobidae急剧减少[45]。菊石的灭绝时间与其他底栖无脊椎动物不同,灭绝的高峰在吴家坪早期[51]。在广西来宾GSSP剖面上发现瓜德鲁普期典型的菊石Kufengoceras subglobosum出现在吴家坪阶底部地层中[38, 52]。表明菊石的灭绝发生在吴家坪早期,灭绝的过程较为漫长,可能与长期的环境变化有关[51]。该次菊石的危机造成“古生代”菊石向“中生代”菊石过渡,也即由游泳型向漂浮型过渡[53]。钙质藻类物种更替的时间几乎与有孔虫灭绝的时间一致。在华南,二叠钙藻Permocalculus、巨孔藻(棒状粗枝藻)Macroporella以及钙质钙扇藻Udoteacean发生灭绝,被裸松藻Gymnocodium、米齐藻Mizzia和假蠕孔藻Pseudovermiporella更替[7, 30, 31]。
总体上看,瓜德鲁普世晚期各种海洋无脊椎动物和钙质藻类均发生了不同程度的灭绝,其灭绝时间又各不相同。最先灭绝的是生活于浅海的珊瑚类,之后腕足类灭绝于卡匹顿晚期,紧接着发生大型有孔虫和钙质藻类的灭绝,最后是常见于深水环境中的菊石类在吴家坪早期发生灭绝。灭绝的时间与其生活的环境有关,窄盐度的浅海无脊椎动物最先发生灭绝,生活环境范围宽、生活水体深的菊石最晚灭绝,说明生物的灭绝受其生存环境压力的影响。
2 峨眉山及其他同时期大火成岩省(LIP)瓜德鲁普末期华南扬子台地西缘发生大规模的玄武岩喷发,被称为峨眉山大火成岩省喷发(图 1)。玄武岩的覆盖面积约0.25×106 km2,体积约0.3×106km3[54]。绝大部分学者认为峨眉山大陆喷发玄武岩是一个地幔柱成因的大火成岩省 [55, 56, 57, 58, 59, 60],其规模相对其他大火成岩省(平均规模为2×106 km3)来说偏小[58]。峨眉山大火成岩省岩石组合包括辉岩、橄榄岩、玄武岩、玄武质安山岩、玄武质火山碎屑岩、流纹岩和粗面岩,其中大部分为玄武岩[54, 61, 62, 63, 64]。流纹岩和粗面岩一般位于这些火山岩的最上部[54, 64, 65, 66]。此外,侵入岩中还发现大量的花岗岩、黑花岗岩、辉长岩和基性岩墙。这些长英质喷出岩是峨眉山大火成岩省的重要组成部分,其他的大火成岩省如Etendeka、Karoo和Yemen也同样具有类似的长英质喷出岩组合[67]。正是这些长英质火成岩的喷发,造成了大量的酸性火山灰沉积。然而,对于遍布俄罗斯远东、华南、以及特提斯洋西部的瓜德鲁普—乐平统界线酸性火山凝灰岩来说[12],这些火山灰是否来自峨眉山玄武岩的喷发?Isozaki[25]认为峨眉山LIP规模太小,酸性喷出岩厚度仅为50 m,不太可能是世界各地G-L界线酸性凝灰岩的来源。He et al.[68]认为由于喷发中心区流纹岩和粗面岩遭受强烈的剥蚀,导致残留的酸性喷出岩较少,其原始规模要比所看到的规模大。
除了峨眉山LIP,同时期还存在位于印度西北部的Panjal大火成岩省,其覆盖面积大约12 000 km2,最大的厚度分布在喀什米尔峡谷,为2.5 km[69],体积不超过0.03×106 km3,仅为峨眉山LIP的十分之一左右。Panjal大火成岩省在地层上夹在下中二叠统地层以及上二叠统深海地层之中[70],喷发的时间与峨眉山LIP大致相同。位于Pangea古陆东部奥斯陆(挪威)[71,72]、阿曼[73]以及澳大利亚西部[74,75]的与地幔有关的碱性火成岩可能与中二叠世来自核幔边界的超级地幔柱有关[25]。然除峨眉山和Panjal之外的火山活动时间还存在疑问。
峨眉山玄武岩40Ar/39Ar年龄为245~256 Ma的年龄[76, 77, 78],这种同位素测年方法比U-Pb法偏年轻且易受到该区后期的区域构造运动的影响[76,77]。而利用锆石SHRIMP、LA-ICP-MS和CA-TIMS U-Pb测年法得到的同位素年龄则较为可靠。部分研究人员直接利用玄武岩来进行锆石U-Pb定年得到253.7±6.1 Ma[78]、251.0±1.0 Ma(玄武质凝灰岩)[79]、259.6±5.9 Ma和259.1±4.0 Ma(斑状玄武岩)[80],其年龄与生物地层年龄产生严重的偏差或者年龄本身存在较大的误差,进一步验证了玄武岩锆石U-Pb测年的不准确性。Zhou et al.[14]对辉长岩锆石U-Pb测年得到较为可靠的年龄258.7±1.5 Ma和256.0±1.0 Ma。对黑花岗和侵入泥盆地层的岩墙的锆石U-Pb测年分别得到261±4 Ma[81]和262±3 Ma[82]的同位素年龄。最近,对峨眉山LIP内带的镁铁质和长英质侵入岩锆石LA-ICP-MS U-Pb定年得到255.4±3.1 Ma至259.5±2.7 Ma的同位素年龄[83]。同年,Shellnutt et al.[84]对攀西地区镁铁质和长英质侵入岩锆石CA-TIMS U-Pb 定年,得到257.6±0.5 Ma至259.6±0.5 Ma的同位素年龄。Zhong et al.[17]对云南平川剖面的玄武岩及火山碎屑岩的CA-TIMS的U-Pb年龄分析,认为峨眉山玄武岩的喷发终止于259.1 Ma。广元朝天剖面G-L界线的王坡页岩火山灰锆石SHRIMP U-Pb同位素年龄为260±4 Ma[57]。这些放射性同位素年龄表明,这些侵入岩的年龄分布较窄,从257 Ma至259 Ma之间(图 2),峨眉山玄武岩的喷发时间间隔约为2 Myr[84],这与古地磁的数据结果大致相当[85, 86, 87]。

晚石炭至早二叠世大冰川之后,中—晚二叠世全球气候逐渐变暖[89,90]。中二叠世沃德末期,在地球内部核幔边界开始形成一个超级地幔柱,外核热不稳定性可诱发地磁极性的反转和不稳定,从而降低地磁强度,允许更多的宇宙射线进入地球大气层,使云层增厚,增强云层对太阳辐射的反射能力,引起地球表面变冷,引发卡穆拉变冷事件[25]。该事件反映在异常高的碳同位素记录之中,可达5‰至6‰[12]。卡穆拉变冷事件之后(或晚期),华南的峨眉山和印度北部的Panjal大火成岩省喷发,同时环特提斯洋地区发生大规模与地幔相关的碱性火成岩侵位。火山喷发释放出大量的CO2、SO2、氧化氮(NOX)、卤素气体、HF、水蒸气和火山灰。这些气体和火山灰如果量足够大,可诱发以下气候效应。SO2与大气中的水蒸气反应生成硫酸盐微粒气溶胶,弥漫在大气对流层或平流层中,反射和吸收太阳辐射[91],与火山灰尘一起可诱发地球表面短期的变冷。如果火山喷出的气体能到达平流层,则该变冷事件能持续达2年,而如果只能到达对流层,微粒将由于被雨水冲洗和参与云朵形成而不能长期停留在大气中,变冷事件仅能持续几天至几十天[92]。火山喷发导致的变冷事件之后,由于火山喷发释放大量的温室气体CO2以及CH4,特别是由于地幔柱上升过程中岩浆的侵位烘烤碳酸盐岩地层以及有机质形成大量的热成因CO2以及甲烷,在短期内(<2 Myr)[84]迅速进入大气圈,造成较长时间的全球变暖。随后,由于玄武岩具有较高的化学活性,这些火成岩发生强烈的化学风化作用,消耗掉大量的CO[93,94]2,造成持续几百千年的全球变冷[95]。全球气候快速变暖常被用来解释生物灭绝的原因[5, 96, 97],然而气候的快速变冷同样(或更加严重)威胁到生物的生存[44, 98]。能否产生这些气候效应,取决于该时期火山喷发规模大小,峨眉山大火成岩省规模小,其同时期的火山喷发规模更小,能否引发如此气候跌宕起伏尚存疑问。虽然Chen et al.[99]通过牙形刺的氧同位素计算出温度高—低—高变化过程,但其也指出这些变化也有可能是牙形刺生活水深变化引起。
4 瓜德鲁普统—乐平统界线Sr、C同位素变化海水Sr同位素组成的变化能表征大火成岩省的喷发及其随后的风化过程[95]。地质历史时期,大火成岩省的喷发能引起海水Sr同位素记录的波动,如Siberia大火成岩省[100]、Karoo-Ferrar大火成岩省[101,102]、以及Deccan高原大火成岩省[103,104]。海水Sr同位素的组成是由洋中脊玄武岩热液蚀变来源的非放射性成因的Sr与古陆风化来源的放射性成因的Sr混合的结果。洋壳热液蚀变来源的非放射性成因的Sr具有较低的87Sr/86Sr比值,约为0.703 0,而古陆化学风化来源的放射性成因的Sr具有较高的87Sr/86Sr比值,平均为0.711 9[105]。因此,主喷发期为卡匹敦中晚期的峨眉山大火成岩省大规模喷发(特别是水下喷发)能增加非放射性成因的Sr同位素的贡献,引起海水Sr同位素比值在该时期降低(图 3)。而后喷发于陆地硅酸盐强烈的化学风化(该化学风化或许同时受到LIP喷出的温室气体引发的气候变暖的影响而加剧)以及海平面的下降则能增加放射性成因的Sr同位素的贡献,从而引起海水Sr同位素比值的升高。海水Sr同位素组成综合反映LIP的喷发和(或)古陆风化的程度。Sr在海水中存留时间约为3 百万年,其同位素比值反映较长时期的海水同位素比值的变化。

|
| 图 3 我国华南G-L界线碳同位素和日本G-L界线87Sr/86Sr比值变化图 碳同位素变化图修改自Jost et al.[21];87Sr/86Sr数据来自Kani et al.[107]。与数字对应的牙形刺带:2.Jinogondolella shannioni; 3.J.altudaensis;4.J.prexuanhaenensis; 5.J. xuanheanensis; 6.J. granti; 7.Clarkina postbitteri hongshuiensis; 8.C. postbitteri postbitteri; 9.C. dukouensis; 10.C. asymmetrica. 牙形刺数据来自Chen et al. [99](蓬莱滩剖面),Saitoh et al. [108](朝天剖面),以及 Bond et al. [2] (贵州综合剖面)。 Fig. 3 Carbon isotopic change in South China and the change of 87Sr/86Sr ratio across G-L Boundary in Japan Carbon isotope data is modified from Jost et al.[21] and 87Sr/86Sr ratios data is from Kani et al.[107]. The number corresponds to conodont zone: 2. Jinogondolella shannioni; 3.J.altudaensis;4.J.prexuanhaenensis; 5.J. xuanheanensis; 6.J. granti; 7.Clarkina postbitteri hongshuiensis; 8.C. postbitteri postbitteri; 9.C. dukouensis; 10.C. asymmetrica. Conodont zone data is from Chen et al.[99](for Penglaitan section),Saitoh et al.[108] (for Chaotian section),and Bond et al.[2](for Guizhou combined section). |
Sr同位素比值在显生宙的最低值出现在瓜德鲁普世[106],在卡匹敦期Sr同位素比值极低,穿过G-L界线,Sr同位素比值逐渐升高[100, 107](图 3)。卡匹敦期Sr同位素的极低值可能与该时期地球内部动力的不稳定性(如伊拉瓦拉磁极反转)有关[25]。核幔边界热不稳定性极可能引发新一轮地幔柱活动的开始[25],将有更多的火山/热液物质进入海洋中,引起Sr同位素比值的降低。Sr同位素的极低值结束的时间与卡匹敦生物灭绝的时间吻合,可能暗示着地球内部动力的变化与生物灭绝事件存在一定的关系。但峨眉山是否与生物灭绝存在关系,在此并不能证明。
G-L界线发生δ13C的负偏[2, 7, 12, 20, 30, 31, 109, 110, 111](图 3)。引发δ13C负偏有多种原因,比如海平面变化[111],海洋分层和缺氧[12],甲烷的释放[1, 112, 113]以及峨眉山火山作用等[31]。然而,该负偏存在一个缺陷:碳同位素负偏在不同剖面出现在不同的牙形刺带(或其他生物带),比如在四川广元朝天剖面负偏位于J. shannoni带[108]、广西来宾蓬莱滩剖面则位于 C. postbitteri postbitteri带[111],两者相差7个牙形刺带,大约3 Ma[15]。而由于G-L界线在华南普遍存在不整合面缺失,导致碳同位素负偏虽然在不同剖面位于不同牙形刺带,但都是穿过G-L界线,也即个别剖面存在长约3 Ma的沉积间断。也正因不整合面的存在,Jost et al.[21]认为 G-L界线幅度达2‰至6‰的碳同位素负偏可能是由于成岩作用或局部埋藏环境所致,而不是代表原始海水的碳同位素信号。Yan et al.[20]通过无机碳—有机碳同位素的差值在蓬莱滩GSSP剖面G-L界线的变化,认为,该剖面的同位素变化代表当时大气碳循环的扰动,碳同位素的负偏反 映的是当时海洋初级生产力的减少。Saitoh et al.[108]则认为G-L界线碳同位素负偏是由于上升洋流带来的富12C的底部贫氧水体所致。而Yan et al.[20]和Saitoh et al.[108]做研究的两个剖面均被Jost et al.[21]认为是成岩作用引起的碳同位素负偏。
5 峨眉山LIP与生物灭绝的成因关系分析 5.1 碳同位素变化与峨眉山LIP喷发的关系Jost et al.[21]认为G-L界线处碳同位素负偏(包括来宾地区剖面)是由于成岩作用造成。然Kaiho et al.[109]通过研究蓬莱滩GSSP剖面的全岩有机碳同位素以及生物标志物长直链烷烃C27的分子同位素比值在G-L界线变化,发现这些同位素比值在G-L界线处存在幅度约为2‰的负偏。利用生物标志物的分子碳同位素分析技术可以剔除有机质母质来源的影响,集中反映生物合成、同化等生物化学如光合作用过程及其成岩作用过程中碳同位素分馏。长直链烷烃C27主要来自高等植物蜡,同位素比值不受成熟度的控制[114]。所以,Kaiho et al.[109]认为来宾市蓬莱滩剖面的G-L界线处碳同位素负偏(幅度为2‰,据其文中图 3)代表当时大气二氧化碳碳同位素扰动。不过,G-L界线的负偏幅度为K-T界线的2/3。与Kaiho et al.[109]的观点类似,Yan et al.[20]认为铁桥剖面碳同位素主要代表海水的原始信号。
蓬莱滩和铁桥剖面的G-L界线处是两种不同岩性的分界线,界线之下是碳酸盐岩,之上是层状硅质岩(图 4);同时该界线也是沉积相突变的界面,界线之下为富含海百合和腕足动物的较为浅水的上斜坡相,之上为深水放射虫丰富深水盆地相[10, 115]。在这种相变的界面上发生碳同位素负偏,有可能是因为深水相有机质矿化过程中富12C孔隙水早期沉淀碳酸盐矿物比例较多所致。在蓬莱滩剖面,Zhong et al.[16]通过研究其界线附近的火山凝灰岩层的地球化学特征,认为,该处火山凝灰岩是陆源碎屑成因,其来源可能是峨眉山LIP,也即峨眉山LIP来源的沉积物与G-L界线处碳同位素负偏时间上吻合,两者可能存在因果关系。而Qiu et al.[116]研究该火山灰层发现火山凝灰岩中存在一部分(约三分之一)的棱角状锆石,这种锆石并没有指示河流搬运的证据,可能部分是空降产物。如此,来宾地区G-L界线火山凝灰岩与峨眉山LIP喷发有关[16]。峨眉山LIP喷发引起华南地壳的抬升或许对来宾G-L界线沉积物地球化学信号有所影响。

|
| 图 4 广西来宾市蓬莱洲中心岛的北岸G-L界线为茅口组碳酸盐岩变至合山组硅质灰岩和硅质岩相变(佳能相机包(20 cm长)作为比例尺) Fig. 4 Field photo showing lithofacies change from the carbonate in the Maokou Formation to the siliceous limestone and chert in the Henshan Formation across the G-L boundary at the north bank of Penglaizhou center island in Laibin city,Guangxi province (Canon camera bag (20 cm wide) for scale) |
与上述间接的证据不同,Wignall et al.[7]报道了一个将峨眉山LIP与G-L界线生物灭绝联系起来的直接证据。他们通过研究贵州织金熊家场剖面的有孔虫、 类与藻类的生物分布、碳酸盐岩碳同位素与该剖面两期玄武岩喷发的关系时发现,位于J. prexuanhanensis-J. xuanhanensis的第一期玄武岩的开始喷发与上述生物灭绝同时,之后发生了幅度达约5‰的碳同位素负偏,他们认为是峨眉山LIP的喷发导致了生物的灭绝,并诱发了同位素比值负偏。之后,Bond et al.[2]归纳贵州省G-L界线碳同位素变化(图 3),认为自卡匹敦阶至吴家坪阶下部碳同位素发生两次负偏,第一次出现在J. prexuanhanensis-J. xuanhanensis牙形刺带,负偏幅度达5‰,第二次出现在G-L界线,负偏幅度仅为2‰。来宾地区两个剖面在在J. prexuanhanensis-J. xuanhanensis牙形刺带碳同位素比值存在微弱负偏(约为1‰),第二次负偏发生在G-L界线(约为2‰)(见参考文献[111, 117])。熊家场剖面在J. prexuanhanensis-J. xuanhanensis牙形刺带岩性为硅质灰岩,含放射虫和硅质海绵骨针[7](图 5)。此硅质灰岩夹在上下两段玄武岩之中,硅的来源很可能就来自玄武岩的喷发。而蓬莱滩剖面在J. prexuanhanensis-J. xuanhanensis牙形刺带的石灰岩中富含交代成因的硅质条带(图 6),硅的来源与峨眉山LIP的喷发有关[118]。蓬莱滩剖面的J. prexuanhanensis-J. xuanhanensis牙形刺带碳同位素负偏可以与熊家场剖面的负偏可能有关联。在日本卡穆拉剖面,同样存在两期类似的碳同位素负偏。尽管没有牙形刺地层的约束,卡穆拉剖面的两次负偏可与蓬莱滩剖面的两次负偏相对比(详见参考文献[12]):第一次碳同位素负偏幅度约为1.5‰,第二次负偏幅度约为2.5‰[12]。对于其他剖面,熊家场剖面卡匹敦中期碳同位素负偏较大,在该剖面负偏段采集样品发现,其硅质灰岩存在重结晶现象(图 7)。且该剖面是一个峨眉山玄武岩喷发地点,火山喷出的气体以及诱发的其他例如甲烷的气体也会造成该地区海水较外围地区更为富12C的溶解无机碳(DIC),这也可能会造成碳同位素负偏幅度相对其他地区更大。对于G-L界线的第二次碳同位素负偏,之前认为的负偏幅度达2.5‰至6.0‰很可能是由于成岩作用的影响(详见[21])。而且,在伊朗剖面,虽然缺乏精细牙形刺地层的标定,然而其G-L界线处也存在一个幅度约1‰的碳同位素负偏[110]。最近Nishikane et al.[119]报道了日本Gujo-hachiman剖面硅质岩干酪根碳同位素比值G-L界线处的负偏幅度均仅为1.4‰。这些数据表明卡匹敦期碳同位素负偏可能存在两期,一期为中卡匹敦期负偏(幅度约为1‰至1.5‰),第一次负偏为G-L界线负偏(幅度约为1.4‰至2‰),但这结论尚待证明。

|
| 图 5 熊家场剖面J. prexuanhanensis-J. xuanhanensis牙形带段硅质灰岩显微镜下特征(含硅质海绵骨针(白色长箭头)和放射虫(黄色短箭头所指的圆粒),单偏光) Fig. 5 Microphotograph of siliceous limestones from J. prexuan-hanensis to J. xuanhanensis zones at Xiongjiachang section(containing siliceous sponge spicules (white arrows) and radiolarian (yellow arrows),Plane polarized light) |

|
| 图 6 西来宾市蓬莱滩剖面瓜德鲁普统至乐平统岩性 柱状图(层一栏中右边对应Jin et al.[48]的编号;T.火山灰;C.硅质岩;M.灰泥石灰岩;W.颗粒质灰泥石灰岩;P.灰泥质颗粒石灰岩) Fig. 6 The columnar section of the Guadalupian to Lopingian succession at the Penglaitan section,Laibin,Guangxi (Note that in the Bed column,the right side is according to Jin et al.[48]; T. tuff; C. chert; M. lime mudstone; W. wackestone; P. packstone.) |

|
| 图 7 贵州熊家场剖面J. prexuanhanensis-J. xuanhanensis(牙形带段重结晶化的硅质灰岩,单偏光) Fig. 7 Partially recrystallized siliceous limestone from the J. prexuanhanensis to J. xuanhanensis zones at the (Xiongjiachang section,Guizhou,Plane polarized light) |
大火成岩省火山喷发释放各种气体和火山灰进入大气中,包括SO2、CO2、H2O、Cl2、CH4以及卤素F。其中SO2、CO2和CH4是较为重要的气体组分[120,121]。SO2是温室气体,其最初的气候效应是区域性甚至全球性长达数月的气候变暖[5]。当SO2与水反应形成硫酸盐气溶胶时,这些固体颗粒能背散射和吸收太阳辐射[91, 120, 121],从而导致可长达数十年的全球变冷(比如[5, 122]),随后由于雨水的不断冲洗作用而消失[123,124]。也即,SO2的气候效应是先变短期变暖再长期变冷。火山灰在大气中停留时间很短,不可能造成明显的气候变冷[91]。Cl2形成数年至数十年的酸雨并能引起臭氧层的破坏[125]。CO2可以长期的停留在大气之中,多次火山喷发的累积效应可以造成全球变暖[5]。
峨眉山LIP的玄武岩体积为0.3×106 km3[59],现代玄武岩每喷发1 km3体积释放3.5×10 12 g[126]或16×1012 g的二氧化碳[127],按照每1 km3释放5×1012 g的二氧化碳计算[5],峨眉山LIP释放出来的二氧化碳为1 500 Gt。根据Berner[128]的模拟计算,假设大气二氧化碳的δ13C为-6‰,峨眉山LIP所含的1 500 Gt的二氧化碳在快速喷出的情况下能引起碳同位素负偏幅度为0.23‰左右,远低于卡匹敦阶至吴家坪阶底部的两次碳同位素负偏的幅度(1‰至2‰)。因此,Wignall et al.[7]认为卡匹敦阶碳同位素负偏是由峨眉山喷发的二氧化碳以及岩浆侵入造成的热成因甲烷及全球变暖海底甲烷冰的释放造成的。Ganino和Arndt[129]认为四川盆地前乐平世地层中含有较多的白云石、页岩和蒸发岩,峨眉山LIP喷发引起的热解变质作用释放出来大量的二氧化碳和甲烷能够引起全球性较大的碳同位素负偏。甲烷一般会引起碳同位素快速负偏,而卡匹敦期两次碳同位素负偏均是长期逐渐负偏,与甲烷释放关系不大[31]。
SO2、卤素F和氯气形成酸雨并毒害植物和动物[5, 130]。海洋动物对痕量金属元素含量敏感,过多的量会使动物中毒,导致生物多样性减少[131]。全球变暖能引发蛋白质的损坏[132]以及新陈代谢氧需求量的增加[133],过高的温度会导致生物的直接死亡[97]。全球气候变冷(cooling)会导致一些适应热带温暖气候的动物死亡[12, 134]。轻碳的大量集中释放进入大气和海洋,改变海洋的物理化学状态如酸碱度,会造成海洋初级生产力崩溃;同时初级生产力的降低将会诱发碳同位素的负偏[20, 135, 136, 137]。海洋初级生产者的大量死亡造成原正常的海洋生物泵受到破坏,导致大量的轻碳进一步累积在海洋之中,很可能会导致海洋向大气反向释放大量的二氧化碳,造成全球变暖[137,138]。同样,规模较小的峨眉山大火成岩省喷发能否引起如此全球性生物和环境事件尚存疑问。
5.3 瓜德鲁普末期生物灭绝的主因分析瓜德鲁普末期生物灭绝早在20世纪末就已经被确定为一个独立的生物灭绝事件[3,4],且其灭绝严重性可与白垩末期生物灭绝以及三叠末期生物灭绝相媲美[4, 27]。经重新计算后,McGhee et al.[36]认为,瓜德鲁普统末期生物危机的严重性在11个显生宙生物多样性危机事件中的排行由之前认为的第三降到第九。遭受灭绝的是一些造礁的底栖生物如海绵[28]以及一些与光合作用生物共生的形态复杂 类(如新希瓦 以及费伯克 ),存活下来的是一些个体小、形态简单的有孔虫[36]。生物礁生态系统由后生生物主导的系统变为由微生物主导的生态系统[139]。总而言之,该生物危机对底栖造礁生态系统、与光合作用共生的有孔虫生态系统以及浮游菊石的生态系统均造成重大影响[36]。引起生物灭绝的原因主要有全球气候变暖[97]、甲烷大量集中释放[7]、火山作用引发的短期变冷[140,141]以及伊拉瓦拉磁极反转引发的长期变冷[12]、海退引起的栖息地的丧失[10, 39, 142]以及海洋的缺氧[13, 143, 144]。
全球变暖能引发生物危机[97]。但对于瓜德鲁普末期生物灭绝事件来说是否如此?Limarino et al.[145]通过沉积学研究认为瓜德鲁普晚期至乐平世为半干旱至干旱气候,在中—晚二叠世过渡时期为从冰室气候变向温室气候过渡。而Fielding et al.[146]认为瓜德鲁普晚期至乐平早期澳大利亚仍然存在冰川沉积。Chen et al.[99]通过对来宾地区蓬莱滩和铁桥两个剖面的牙形刺氧同位素分析,计算出来的温度在J. xuanhanensis至C. postbitteri postbitteri有升高趋势,但他们认为这个结果是由于气候和海平面变化双重因素导致的,也即不一定代表当时全球气温的升高。全球变暖能引起海洋酸化、碳酸盐浓度过高以及海洋缺氧。海洋温度升高会降低其从大气中去除二氧化碳的能力[147]。大气二氧化碳浓度的升高导致海洋表层水体CO2浓度升高从而引发海洋酸化。Clapham和Payne[148]认为由于海洋酸化和海洋缺氧等影响,卡匹敦期海洋退化而引发了一些高钙质生物的生理危机,但是该次生理危机比二叠末期的危机要弱。Jost et al.[21]通过全球多个剖面的Ca同位素分析发现,卡匹敦期至吴家坪早期并没有存在明显的海洋酸化特征。但由于瓜德鲁普末期生物危机事件的主要灭绝生物是一些高度钙化的皱壁珊瑚、 类和部分腕足类等,这些生物对海洋的物理化学条件的改变较为敏感,较弱的海洋酸化以及高碳酸血症也有可能弱化其生态系统从而引发生物灭绝。
气候的突然变冷能够引起一些适应温暖气候的动物产生不适而死亡,如奥陶晚期生物灭绝[149, 150, 151, 152, 153]。对于瓜德鲁普末期生物灭绝事件来说,峨眉山LIP喷发释放的SO2形成硫酸盐气溶胶以及火山灰等引发的短期变冷虽然能引起喜温动物的不适,但由于其时间过短,不太可能成为瓜德鲁普末期生物灭绝的主要原因。日本学者Isozaki et al.[12, 154]认为长期的卡穆拉变冷事件是瓜德鲁普末期低纬度适宜暖水动物死亡的直接原因。卡穆拉变冷事件作为瓜德鲁普末期生物灭绝的原因尚存在两个问题:一是时间问题,卡穆拉事件发生的时间是早—中卡匹敦期Yabeina 带[154],而生物灭绝主要发生在卡匹敦中期和晚期[155](图 1),且变冷事件引起的生物灭绝一般发生在变冷的开始阶段,如晚奥陶世生物灭绝事件;二是全球性问题,虽然Isozaki et al.[12, 154, 156]在当时位于泛大洋中部的日本卡穆拉地区环礁石灰岩和当时位于古特提斯洋最西部的克罗地亚Brusane剖面碳酸盐中均发现了卡穆拉变冷事件,但在特提斯东部的我国华南地区如来宾蓬莱滩和铁桥剖面[117]和广元市上寺剖面[157]中均没有发现卡穆拉事件。最近,Kirschvink et al.[158]利用超导磁力计降噪技术的提高,在卡穆拉地区G-L界线碳酸盐岩中测出较好的古磁极数据,并通过与我国华南磁极对比,认为卡穆拉事件可能对应着J. postserrata至J. granti牙形刺带,也即中—晚卡匹敦期,正好对应着生物灭绝的主要时期以及峨眉山LIP主喷发期。Kirschvink et al.[158]的数据是基于卡穆拉地区10个零散剖面拼接起来的,而不是一个连续的剖面,其磁极对比的可靠性尚存疑问。Vogt[131]认为,石炭纪末期及第四纪冰期并没有引起大的生物灭绝,且在极度寒冷的气候下,一些热带生物能够存活下来,变冷作为生物灭绝的原因难于让人信服。短期变冷则由于硫酸盐气溶胶在大气中存在时间较短难以引起较明显的气候变化,导致其地质证据较少(如参考文献[5]),这也可能是制约其作为生物灭绝原因的因素之一。
海平面下降导致适合生物生存的浅海栖息地的缺失,增大生物的生存环境压力从而引发生物灭绝[159]。瓜德鲁普末期几乎是全球海平面在显生宙最低的时期[8, 25]。由于海退的时间与生物灭绝的时间较为吻合,该时期大规模的海退被认为是瓜德鲁普末期生物灭绝的重要的原因[10, 11, 31, 39, 42, 142, 159]。然而,地球历史过程中,一些大规模的海退并没有引起明显的生物灭绝,比如第四纪冰川引发的大规模海退缺乏生物危机事件相伴。海退之后,虽然原来的栖息地丧失,但生活在浅海的生物会随之迁徙到浅海地带,从这一点来说,这些浅海生物仍然有生存的空间,只不过生存空间可能受到压缩减少。大规模海退可以是生物灭绝的原因之一,但不是唯一的原因。
海洋缺氧同样能引起可生活的栖息地减少,与海平面下降相比,海洋缺氧能直接杀死海洋生物。海洋缺氧是指海水中不含氧或含有硫化氢气体,这样的条件不适合需氧生物生存。因此,海洋缺氧普遍被认为是生物灭绝的主要原因[160]。G-L过渡期海洋的氧化还原条件的研究较少。Isozaki[143]在分析日本西南部深海相硅质岩剖面时观察到瓜德鲁普统的岩性为红色氧化的硅质岩,而之上的吴家坪阶地层为灰色缺氧的硅质岩,认为G-L时期深海海水由氧化状态变为还原状态。Saitoh et al.[13, 144]认为朝天剖面G-L界线存在贫氧至缺氧段,并推测盆地环境中存在硫化水体,这些硫化水体的上涌造成浅海底栖生物的灭亡。Yan et al.[20]认为来宾铁桥剖面G-L界线硫同位素比值的负偏(-37‰)很可能是由于当时海水的氧化还原分层,富含硫化氢的水体随着海侵毒害浅海底栖生物而导致生物灭绝。这些研究均表明,G-L界线附近存在海洋的缺氧,且与生物灭绝的时间基本一致,说明两者之间存在联系。然而,这些研究还存在一些问题。Isozaki[143]的研究没有生物灭绝数据,无法直接分析海洋的缺氧与瓜德鲁普末期生物灭绝的关系。Yan et al.[20]和Saitoh et al.[13, 144]的研究缺氧事件不是出现在同一个牙形刺带,前者来宾铁桥剖面缺氧层位出现在J. granti至C. postbitteri hongshuiensis带,而后者广元市朝天剖面缺氧层位出现在J. shannoni带,比前者早了三个牙形刺带。这些矛盾都需要对氧化还原条件做进一步的研究。目前作者正在开展相关研究。
上述分析表明,气候变暖、大规模海平面下降以及海洋缺氧的时间大致峨眉山大火成岩省的喷发同步。但峨眉山大火成岩省过小的规模与这些气候和环境的剧变相矛盾。峨眉山大火成岩省的喷发会造成一定的气候和环境变化,这些变化很可能不会直接引起生物大灭绝。 可能还会存在其他的环境因素。中二叠世末期是地球内部动力变化的开始,这变化很可能会引起地球表生系统产生相应的环境气候剧变,从而引起生物灭绝。这些剧变的厘定与全球海平面剧变、全球海水氧化还原以及古海洋初级生产力大小有关。这几个因素的变化与峨眉山大火成岩省喷发演变过程的关系如何,目前尚不清楚。所以否定峨眉山大火成岩省喷发对该时期生物灭绝的影响条件不充分。
6 未来研究展望大规模海退与海洋缺氧最有可能成为瓜德鲁普末期生物灭绝的两个主要原因。海平面变化的研究较多,该时期大规模海退基本已经达成共识。大规模海退必然会驱使浅海生物向深海方向迁移,而该时期深水的氧化还原条件控制底栖生物灭绝的方式。而该时期海洋氧化还原尚未充分研究,海洋的氧化还原状态尚未完全清楚,是未来的一个主要研究方向。此外,瓜德鲁普世海洋具有高的海洋初级生产力,而乐平统初期海洋初级生产力下降。该海洋初级生产力下降受什么条件控制,它与该时期生物灭绝有什么样的关系?这方面还缺乏详细的研究,是研究该时期生物灭绝的一个突破口。
致谢 非常感谢两位审稿专家提出的富有建设性的建议。
| [1] | Bond D P G, Hilton J, Wignall P B, et al. The Middle Permian (Capitanian) mass extinction on land and in the oceans[J]. Earth-Science Reviews, 2010, 102(1/2): 100-116. |
| [2] | Bond D P G, Wignall P B, Wang Wei, et al. The mid-Capitanian (Middle Permian) mass extinction and carbon isotope record of South China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2010, 292(1/2): 282-294. |
| [3] | Jin Yugan, Zhang Jing, Shang Qinghua. Two phases of the end-Permian mass extinction[M]//Embry A F, Beauchamp B, Glass D J. Pangea: Global Environments and Resources. Calgary: Memoir Canadian Society of Petroleum Geologists, 1994, 17: 813-822. |
| [4] | Stanley S M, Yang X. A double mass extinction at the end of the Paleozoic era[J]. Science, 1994, 266(5189): 1340-1344. |
| [5] | Wignall P B. Large igneous provinces and mass extinctions[J]. Earth-Science Reviews, 2001, 53(1/2): 1-33. |
| [6] | Courtillot V E, Renne P R. On the ages of flood basalt events[J]. Comptes Rendus Geoscience, 2003, 335(1): 113-140. |
| [7] | Wignall P B, Sun Yadong, Bond D P G, et al. Volcanism, mass extinction, and carbon isotope fluctuations in the Middle Permian of China[J]. Science, 2009, 324(5931): 1179-1182. |
| [8] | Haq B U, Schutter S R. A chronology of Paleozoic sea-level changes[J]. Science, 2008, 322(5898): 64-68. |
| [9] | Wignall P B, Védrine S, Bond D P G, et al. Facies analysis and sea-level change at the Guadalupian-Lopingian global stratotype (Laibin, South China), and its bearing on the end-Guadalupian mass extinction[J]. Journal of the Geological Society, 2009, 166(4): 655-666. |
| [10] | Qiu Zhen, Wang Qingchen, Zou Caineng, et al. Transgressive-regressive sequences on the slope of an isolated carbonate platform (Middle-Late Permian, Laibin, South China)[J]. Facies, 2014, 60(1): 327-345. |
| [11] | Kofukuda D, Isozaki Y, Igo H. A remarkable sea-level drop and relevant biotic responses across the Guadalupian-Lopingian (Permian) boundary in low-latitude mid-Panthalassa: Irreversible changes recorded in accreted paleo-atoll limestones in Akasaka and Ishiyama, Japan[J]. Journal of Asian Earth Sciences, 2014, 82: 47-65. |
| [12] | Isozaki Y, Kawahata H, Minoshima K. The Capitanian (Permian) Kamura cooling event: the beginning of the Paleozoic-Mesozoic Transition[J]. Palaeoworld, 2007, 16(1/2/3): 16-30. |
| [13] | Saitoh M, Isozaki Y, Yao Jianxin, et al. The appearance of an oxygen-depleted condition on the Capitanian disphotic slope/basin in South China: middle-upper Permian stratigraphy at Chaotian in northern Sichuan[J]. Global and Planetary Change, 2013, 105: 180-192. |
| [14] | Zhou Meifu, Malpas J, Song Xieyan, et al. A temporal link between the Emeishan large igneous province (SW China) and the end-Guadalupian mass extinction[J]. Earth and Planetary Science Letters, 2002, 196(3/4): 113-122. |
| [15] | Sun Yadong, Lai Xulong, Wignall P B, et al. Dating the onset and nature of the middle Permian Emeishan large igneous province eruptions in SW China using conodont biostratigraphy and its bearing on mantle plume uplift models[J]. Lithos, 2010, 119(1/2): 20-33. |
| [16] | Zhong Yuting, He Bin, Xu Yigang. Mineralogy and geochemistry of claystones from the Guadalupian-Lopingian boundary at Penglaitan, South China: insights into the pre-Lopingian geological events[J]. Journal of Asian Earth Sciences, 2013, 62: 438-462. |
| [17] | Zhong Yuting, He Bin, Mundil R, et al. CA-TIMS zircon U-Pb dating of felsic ignimbrite from the Binchuan section: implications for the termination age of Emeishan large igneous province[J]. Lithos, 2014, 204: 14-19. |
| [18] | Courtillot V, Jaupart C, Manighetti I, et al. On causal links between flood basalts and continental breakup[J]. Earth and Planetary Science Letters, 1999, 166(3/4): 177-195. |
| [19] | Wang Xiangdong, Sugiyama T. Diversity and extinction patterns of Permian coral faunas of China[J]. Lethaia, 2000, 33(4): 285-294. |
| [20] | Yan Detian, Zhang Liqin, Qiu Zhen. Carbon and sulfur isotopic fluctuations associated with the end-Guadalupian mass extinction in South China[J]. Gondwana Research, 2013, 24(3/4): 1276-1282. |
| [21] | Jost A B, Mundil R, He Bin, et al. Constraining the cause of the end-Guadalupian extinction with coupled records of carbon and calcium isotopes[J]. Earth and Planetary Science Letters, 2014, 396: 201-212. |
| [22] | Raup D M, Sepkoski Jr J J. Mass extinctions in the marine fossil record[J]. Science, 1982, 215(4539): 1501-1503. |
| [23] | Sepkoski Jr J J. Patterns of Phanerozoic extinction: a perspective from global data bases[M]//Walliser O H. Global Events and Event Stratigraphy in the Phanerozoic. Berlin: Springer, 1996: 35-51. |
| [24] | Shen Shuzhong, Crowley J L, Wang Yue, et al. Calibrating the End-Permian mass extinction[J]. Science, 2011, 334(6061): 1367-1372. |
| [25] | Isozaki Y. Illawarra reversal: the fingerprint of a super plume that triggered Pangean breakup and the end-Guadalupian (Permian) mass extinction[J]. Gondwana Research, 2009, 15(3/4): 421-432. |
| [26] | 金玉玕,张进,尚庆华. 前乐平统海洋动物灾变事件[J]. 古生物学报,1995,34(4):410-427. [Jin Yugan, Zhang Jin, Shang Qinghua. Pre-Lopingian catastrophic event of marine faunas[J]. Acta Palaeontologica Sinica, 1995, 34(4): 410-427.] |
| [27] | Knoll A H, Bambach R K, Canfield D E, et al. Comparative earth history and late Permian mass extinction[J]. Science, 1996, 273(5274): 452-457. |
| [28] | Weidlich O. Permian reefs re-examined: extrinsic control mechanisms of gradual and abrupt changes during 40 my of reef evolution[J]. Geobios, 2002, 35(S1): 287-294. |
| [29] | Clapham M E, Shen Shuzhong, Bottjer D J. The double mass extinction revisited: reassessing the severity, selectivity, and causes of the end-Guadalupian biotic crisis (Late Permian)[J]. Paleobiology, 2009, 35(1): 32-50. |
| [30] | Lai Xulong, Wang Wei, Wignall P B, et al. Palaeoenvironmental change during the end-Guadalupian (Permian) mass extinction in Sichuan, China[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2008, 269(1/2): 78-93. |
| [31] | Wei Hengye, Chen Daizhao, Yu Hao, et al. End-Guadalupian mass extinction and negative carbon isotope excursion in Xiaojiaba section, Guangyuan, Sichuan[J]. Science China Earth Sciences, 2012, 55(9): 1480-1488. |
| [32] | Bond D P G, Wignall P B. Latitudinal selectivity of foraminifer extinctions during the Late Guadalupian crisis[J]. Paleobiology, 2009, 35(4): 465-483. |
| [33] | Yang Xianning, Zhou Jianping, Liu Jiarun, et al. Evolutionary pattern of fusulinacean foraminifer in Maokouan, Middle Permian[J]. Science in China Series D: Earth Sciences, 1999, 42(5): 456-464. |
| [34] | Yang Xianning, Liu Jiarun, Shi Guijun. Extinction process and patterns of Middle Permian fusulinaceans in southwest China[J]. Lethaia, 2004, 37(2): 139-147. |
| [35] | Ota A, Isozaki Y. Fusuline biotic turnover across the Guadalupian-Lopingian (Middle-Upper Permian) boundary in mid-oceanic carbonate buildups: biostratigraphy of accreted limestone in Japan[J]. Journal of Asian Earth Sciences, 2006, 26(3/4): 353-368. |
| [36] | McGhee Jr G R, Clapham M E, Sheehan P M, et al. A new ecological-severity ranking of major Phanerozoic biodiversity crises[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2013, 370: 260-270. |
| [37] | Zhang Zhou, Wang Yue, Zheng Quanfeng. Middle Permian smaller foraminifers from the Maokou formation at the Tieqiao section, Guangxi, South China[J]. Palaeoworld, 2015, 24(3): 263-276. |
| [38] | Shen Shuzhong, Shi G R. Latest Guadalupian brachiopods from the Guadalupian/Lopingian boundary GSSP section at Penglaitan in Laibin, Guangxi, South China and implications for the timing of the pre-Lopingian crisis[J]. Palaeoworld, 2009, 18(2/3): 152-161. |
| [39] | Shen Shuzhong, Shi G R. Paleobiogeographical extinction patterns of Permian brachiopods in the Asian-western Pacific region[J]. Paleobiology, 2002, 28(4): 449-463. |
| [40] | Erwin D H, Pan H Z. Recoveries and radiations: gastropods after the Permian - Triassic mass extinction[M]//Hart M B. Biotic Recovery from Mass Extinction Events. Plymouth: Geological Society London Special Publication, 1996, 102(1): 223-229. |
| [41] | 孙东立,沈树忠. 华南二叠纪—三叠纪腕足动物多样性模式[M]//戎嘉余,方宗杰. 生物大灭绝与复苏:来自华南古生代和三叠纪的证据:下卷. 合肥:中国科学技术大学出版社,2004:543-569,1066. [Sun Dongli, Shen Shuzhong. The diversity patterns of brachiopod of Permian - Triassic in South China[M]//Rong Jiayu, Fang Zongjie. Mass Extinction and Recovery—Evidence from Paleozoic and Triassic of South China. Hefei: Press of University of Science and Technology of China, 2004: 543-569, 1066.] |
| [42] | Shen Shuzhong, Shi G R. Diversity and extinction patterns of Permian brachiopoda of South China[J]. Historical Biology: An International Journal of Paleobiology, 1996, 12(2): 93-110. |
| [43] | Aljinovic' D, Isozaki Y, Sremac J. The occurrence of giant bivalve Alatoconchidae from the Yabeina zone (Upper Guadalupian, Permian) in European Tethys[J]. Gondwana Research, 2008, 13(3): 275-287. |
| [44] | Isozaki Y, Aljinovic' D. End-Guadalupian extinction of the Permian gigantic bivalve Alatoconchidae: end of gigantism in tropical seas by cooling[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2009, 284(1/2): 11-21. |
| [45] | 杨遵义,吴顺宝,殷鸿福,等. 华南二叠—三叠纪过渡期地质事件[M]. 北京:地质出版社,1991. [Yang Zunyi, Wu Shunbao, Yin Hongfu, et al. The Geological Events from Permian to Triassic Transition in South China[M]. Beijing: Geological Publishing House, 1991.] |
| [46] | Erwin D H. Understanding biotic recoveries: extinction, survival, and preservation during the end-Permian mass extinction[M]//Jablonski D, Erwin D H, Lipps J H. Evolutionary Paleobiology. Chicago: University of Chicago Press, 1996: 398-418. |
| [47] | 潘华璋. 二叠纪至中三叠世腹足类灭绝与复苏评述[M]//戎嘉余,方宗杰. 生物大灭绝与复苏—来自华南古生代和三叠纪的证据. 合肥:中国科学技术大学出版社,2004:719-729,1071. [Pan Huazhang. The recovery snails extinction in Permian-Triassic[M]//Rong Jiayu, Fang Zongjie. Mass Extinction and Recovery—Evidence from Paleozoic and Triassic of South China. Hefei: Press of University of Science and Technology of China, 2004: 719-729, 1071.] |
| [48] | Jin Yugan, Shen Shuzhong, Henderson C M, et al. The Global Stratotype Section and Point (GSSP) for the boundary between the Capitanian and Wuchiapingian Stage (Permian)[J]. Episodes, 2006, 29(4): 253-262. |
| [49] | Chen Zhongqiang, George A D, Yang Wanrong. Effects of Middle-Late Permian sea-level changes and mass extinction on the formation of the Tieqiao skeletal mound in the Laibin area, South China[J]. Australian Journal of Earth Sciences, 2009, 56(6): 745-763. |
| [50] | Brayard A, Escarguel G, Bucher H, et al. Good genes and good luck: ammonoid diversity and the end-Permian mass extinction[J]. Science, 2009, 325(5944): 1118-1121. |
| [51] | Villier L, Korn D. Morphological disparity of Ammonoids and the mark of Permian mass extinctions[J]. Science, 2004, 306(5694): 264-266. |
| [52] | Ehiro M, Shen Shuzhong. Permian ammonoid Kufengoceras from the uppermost Maokou Formation (earliest Wuchiapingian) at Penglaitan, Laibin Area, Guangxi Autonomous Region, South China[J]. Paleontological Research, 2008, 12(3): 255-259. |
| [53] | Leonova T B. Ammonoid evolution in marine ecosystems prior to the Permian-Triassic crisis[J]. Paleontological Journal, 2009, 43(8): 858-865. |
| [54] | Xu Yigang, Chung Sunlin, Jahn B M, et al. Petrologic and geochemical constraints on the petrogenesis of Permian-Triassic Emeishan flood basalts in southwestern China[J]. Lithos, 2001, 58(3/4): 145-168. |
| [55] | Chung S L, Jahn B M. Plume-lithosphere interaction in generation of the Emeishan flood basalts at the Permian-Triassic boundary[J]. Geology, 1995, 23(10): 889-892. |
| [56] | Chung S L, Jahn B M, Wu G Y, et al. The Emeishan flood basalt in SW China: a mantle plume initiation model and its connection with continental breakup and mass extinction at the Permian - Triassic boundary[M]//Flower M F J, Chung S L, Lo C H, et al. Mantle Dynamics and Plate Interactions in East Asia. Washington: American Geophysical Union Geodynamic Series, 1998, 27: 47-58. |
| [57] | He Bin, Xu Yigang, Huang Xiaolong, et al. Age and duration of the Emeishan flood volcanism, SW China: geochemistry and SHRIMP zircon U-Pb dating of silicic ignimbrites, post-volcanic Xuanwei Formation and clay tuff at the Chaotian section[J]. Earth and Planetary Science Letters, 2007, 255(3/4): 306-323. |
| [58] | Ali J R, Thompson G M, Zhou Meifu, et al. Emeishan large igneous province, SW China[J]. Lithos, 2005, 79(3/4): 475-489. |
| [59] | Xu Yigang, He Bin, Chung Sunlin, et al. Geologic, geochemical, and geophysical consequences of plume involvement in the Emeishan flood-basalt province[J]. Geology, 2004, 32(10): 917-920. |
| [60] | Ali J R, Fitton J G, Herzberg C. Emeishan large igneous province (SW China) and the mantle-plume up-doming hypothesis[J]. Journal of the Geological Society, 2010, 167(5): 953-959. |
| [61] | Song Xieyan, Zhou Meifu, Hou Zhengqian, et al. Geochemical constraints on the mantle source of the upper Permian Emeishan continental flood basalts, southwestern China[J]. International Geology Review, 2001, 43(3): 213-225. |
| [62] | Xiao Long, Xu Y G, Mei H J, et al. Distinct mantle sources of low-Ti and high-Ti basalts from the western Emeishan large igneous province, SW China: implications for plume-lithosphere interaction[J]. Earth and Planetary Science Letters, 2004, 228(3/4): 525-546. |
| [63] | Hanski E, Kamenetsky V S, Luo Zhenyu, et al. Primitive magmas in the Emeishan large igneous province, southwestern China and northern Vietnam[J]. Lithos, 2010, 119(1/2): 75-90. |
| [64] | Shellnutt J G, Jahn B M. Formation of the Late Permian Panzhihua plutonic-hypabyssal-volcanic igneous complex: implications for the genesis of Fe-Ti oxide deposits and A-type granites of SW China[J]. Earth and Planetary Science Letters, 2010, 289(3/4): 509-519. |
| [65] | 张云湘,骆耀南,杨崇喜. 攀西裂谷[M]. 北京:地质出版社,1988: 466. [Zhang Yunxiang, Luo Yaonan, Yang Chongxi. The Panxi Rift[M]. Beijing: Geological Publising House, 1988: 466.] |
| [66] | Ali J R, Wignall P B. Comment on “Fusiline biotic turnover across the Guadalupian-Lopingian (middle-upper permian) boundary in mid-oceanic carbonate build-ups: biostratigraphy of accreted limestone in Japan” by Ayano Ota and Yukio Isozaki[J]. Journal of Asian Earth Sciences, 2007, 30(1): 199-200. |
| [67] | Bryan S E, Rilly T R, Jerram D A, et al. Silicic volcanism: an under-valued component of large igneous provinces and volcanic rifted margins[M]//Menzies M A, Klemperer S L, Ebinger C J, et al. Volcanic Rifted Margins. Boulder, Colorado: Geological Society of American Special Paper, 2002, 362: 99-120. |
| [68] | He Bin, Xu Yigang, Zhong Yuting, et al. The Guadalupian-Lopingian boundary mudstones at Chaotian (SW China) are clastic rocks rather than acidic tuffs: implication for a temporal coincidence between the end-Guadalupian mass extinction and the Emeishan volcanism[J]. Lithos, 2010, 119(1/2): 10-19. |
| [69] | Honegger K, Dietrich V, Frank W, et al. Magmatism and metamorphism in the Ladakh Himalayas (the Indus-Tsangpo suture zone)[J]. Earth and Planetary Science Letters, 1982, 60(2): 253-292. |
| [70] | Nakazawa K, Kapoor H M, Ishii K, et al. The upper Permian and the lower Triassic in Kashmir, India[J]. Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy, 1975, 42: 1-106. |
| [71] | Neumann E R, Olsen K H, Baldridge W S, et al. The Oslo rift: a review[J]. Tectonophysics, 1992, 208(1/2/3): 1-18. |
| [72] | Larson B T, Olaussen S, Sundvoll B, et al. The Permo-Carboniferous Oslo rift through six stages and 65 million years[J]. Episodes, 2008, 31(1): 52-58. |
| [73] | Rabu D, Le Metour J, Bechennec F, et al. Sedimentary aspects of the Eo-Alpine cycle on the northeast edge of the Arabian Platform (Oman Mountains)[M]//Robertson A H F, Searle M P, Ries A C. The Geology and Tectonics of the Oman Region. London: Geological Society of London Special Publication, 1990, 49: 49-68. |
| [74] | Le Maitre R W. Volcanic rocks from edel no.1 petroleum exploration well, offshore Carnarvon basin, Western Australia[J]. Journal of the Geological Society Australia, 1975, 22(2): 167-174. |
| [75] | Veevers J J, Tewari R C. Permian-Carboniferous and Permian-Triassic magmatism in the rift zone bordering the Tethyan margin of southern Pangea[J]. Geology, 1995, 23(5): 467-470. |
| [76] | Boven A, Pasteels P, Punzalan L E, et al. 40Ar/39Ar geochronological constraints on the age and evolution of the Permo-Triassic Emeishan volcanic province, southwest China[J]. Journal of Asian Earth Sciences, 2002, 20(2): 157-175. |
| [77] | Ali J R, Lo C H, Thompson G M, et al. Emeishan basalt Ar-Ar overprint ages define several tectonic events that affected the western Yangtze platform in the Mesozoic and Cenozoic[J]. Journal of Asian Earth Sciences, 2004, 23(2): 163-178. |
| [78] | 范蔚茗,王岳军,彭头平,等. 桂西晚古生代玄武岩Ar-Ar和U-Pb年代学及其对峨眉山玄武岩省喷发时代的约束[J]. 科学通报,2004,49(18):1892-1900. [Fan Weiming, Wang Yuejun, Peng Touping, et al. Ar-Ar and U-Pb geochronology of Late Paleozoic basalts in western Guangxi and its constraints on the eruption age of Emeishan basalt magmatism [J]. Chinese Science Bulletin, 2004, 49(18): 1892-1900.] |
| [79] | 朱江,张招崇,侯通,等. 贵州盘县峨眉山玄武岩系顶部凝灰岩LA-ICP-MS锆石U-Pb年龄:对峨眉山大火成岩省与生物大规模灭绝关系的约束[J]. 岩石学报,2011,27(9):2743-2751. [Zhu Jiang, Zhang Zhaochong, Hou Tong, et al. LA-ICP-MS zircon U-Pb geochronology of the tuffs on the uppermost of the Emeishan basalt succession in Panxian county, Guizhou province: constraints on genetic link between Emeishan large igneous province and the mass extinction[J]. Acta Petrologica Sinica, 2011, 27(9): 2743-2751.] |
| [80] | Fan Weiming, Zhang Chunhong, Wang Yuejun, et al. Geochronology and geochemistry of Permian basalts in western Guangxi Province, Southwest China: evidence for plume-lithosphere interaction[J]. Lithos, 2008, 102(1/2): 218-236. |
| [81] | Luo Zhenyu, Xu Yigang, He Bin, et al. Geochronologica and petrochemical evidence for the genetic link between the Maomaogou nepheline syenites and the Emeishan large igneous provinces[J]. Chinese Science Bulletin, 2007, 52(7): 949-958. |
| [82] | Guo Feng, Fan Weiming, Wang Yuejun, et al. When did the Emeishan mantle plume activity start? geochronological and geochemical evidence from ultramafic-mafic dikes in southwestern China[J]. International Geology Review, 2004, 46(3): 226-234. |
| [83] | Zhong Hong, Campbell I H, Zhu Weiguang, et al. Timing and source constraints on the relationship between mafic and felsic intrusions in the Emeishan large igneous province[J]. Geochimica et Cosmochimica Acta, 2011, 75(5): 1374-1395. |
| [84] | Shellnutt J G, Denyszyn S W, Mundil R. Precise age determination of mafic and felsic intrusive rocks from the Permian Emeishan large igneous province (SW China)[J]. Gondwana Research, 2012, 22(1): 118-126. |
| [85] | Huang Kainian, Opdyke N D. Magnetostratigraphic investigations on an Emeishan basalt section in western Guizhou Province, China[J]. Earth and Planetary Science Letters, 1998, 163(1/2/3/4): 1-14. |
| [86] | Ali J R, Thompson G M, Song Xieyan, et al. Emeishan Basalts (SW China) and the 'end-Guadalupian’ crisis: magnetobiostratigraphic constraints[J]. Journal of the Geological Society, 2002, 159(1): 21-29. |
| [87] | Zheng Liandi, Yang Zhenyu, Tong Yabo, et al. Magnetostratigraphic constraints on two-stage eruptions of the Emeishan continental flood basalts[J]. Geochemistry, Geophysics, Geosystems, 2010, 11(12): Q12014, doi: 10.1029/2010GC003267. |
| [88] | Shellnutt J G. The Emeishan large igneous province: a synthesis[J]. Geoscience Frontiers, 2014, 5(3): 369-394. |
| [89] | Crowell J C. Pre-Mesozoic ice ages: their bearing on understanding the climate system[J]. Geological Society of America Memoirs, 1999, 192: 1-112. |
| [90] | Jones A T, Fielding C R. Sedimentological record of the late Paleozoic glaciation in Queensland, Australia[J]. Geology, 2004, 32(2): 153-156. |
| [91] | Devine J D, Sigurdsson H, Davis A N, et al. Estimates of sulfur and chlorine yield to the atmosphere from volcanic eruptions and potential climatic effects[J]. Journal of Geophysical Research, 1984, 89(B7): 6309-6325. |
| [92] | Self S, Thordarson T, Widdowson M. Gas fluxes from flood basalt eruptions[J]. Elements, 2005, 1(5): 283-287. |
| [93] | Dessert C, Dupré B, François L M, et al. Erosion of Deccan Traps determined by river geochemistry: impact on the global climate and the 87Sr/86Sr ratio of seawater[J]. Earth and Planetary Science Letters, 2001, 188(3/4): 459-474. |
| [94] | Dessert C, Dupré B, Gaillardet J, et al. Basalt weathering laws and the impact of basalt weathering on the global carbon cycle[J]. Chemical Geology, 2003, 202(3/4): 257-273. |
| [95] | Cohen A S, Coe A L. The impact of the central Atlantic Magmatic Province on climate and on the Sr- and Os-isotope evolution of seawater[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2007, 244(1/2/3/4): 374-390. |
| [96] | Racki G, Wignall P. Late Permian double-phased mass extinction and volcanism: an oceanographic perspective[M]//Over D J, Morrow J R, Wignall P B. Understanding Late Devonian and Permian-Triassic Biotic and Climatic events: Towards an Integrated Approach. Cambridge: Cambridge University Press, 2005: 263-297. |
| [97] | Sun Yadong, Joachimski M M, Wignall P B, et al. Lethally hot temperatures during the Early Triassic greenhouse[J]. Science, 2012, 338(6105): 366-370. |
| [98] | Roscher M, Stordal F, Svensen H. The effect of global warming and global cooling on the distribution of the latest Permian climate zones[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 309(3/4): 186-200. |
| [99] | Chen Bo, Joachimski M M, Sun Yadong, et al. Carbon and conodont apatite oxygen isotope records of Guadalupian-Lopingian boundary sections: climatic or sea-level signal?[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 311(3/4): 145-153. |
| [100] | Martin E E, Macdougall J D. Sr and Nd isotopes at the Prmian/Triassic Boundary: a record of climate change[J]. Chemical Geology, 1995, 125(1/2): 73-99. |
| [101] | Jones C E, Jenkyns H C. Seawater strontium isotopes, oceanic anoxic events, and seafloor hydrothermal activity in the Jurassic and Cretaceous[J]. American Journal of Science, 2001, 301(2): 112-149. |
| [102] | McArthur J M, Howarth R J, Bailey T R. Strontium isotope stratigraphy: LOWESS version 3: best fit to the marine Sr-isotope curve for 0-509 Ma and accompanying look-up table for deriving numerical age[J]. The Journal of Geology, 2001, 109(2): 155-170. |
| [103] | Martin E E, Macdougall J D. Seawater Sr isotopes at the Cretaceous/Tertiary boundary[J]. Earth and Planetary Science Letters, 1991, 104(2/3/4): 166-180. |
| [104] | Vonhof H B, Smit J. High-resolution late Maastrichtian-early Danian oceanic 87Sr/86Sr record: implications for Cretaceous-Tertiary boundary events[J]. Geology, 1997, 25(4): 347-350. |
| [105] | Palmer M R, Edmond J M. The strontium isotope budget of the modern ocean[J]. Earth and Planetary Science Letters, 1989, 92(1): 11-26. |
| [106] | Veizer J, Ala D, Azmy K, et al. 87Sr/86Sr, δ13C and δ18O evolution of Phanerozoic seawater[J]. Chemical Geology, 1999, 161(1/2/3): 59-88. |
| [107] | Kani T, Hisanabe C, Isozaki Y. The Capitanian (Permian) minimum of 87Sr/86Sr ratio in the mid-Panthalassan paleo-atoll carbonates and its demise by the deglaciation and continental doming[J]. Gondwana Research, 2013, 24(1): 212-221. |
| [108] | Saitoh M, Isozaki Y, Ueno Y, et al. Middle-Upper Permian carbon isotope stratigraphy at Chaotian, South China: pre-extinction multiple upwelling of oxygen-depleted water onto continental shelf[J]. Journal of Asian Earth Sciences, 2013, 67-68: 51-62. |
| [109] | Kaiho K, Chen Zhongqiang, Ohashi T, et al. A negative carbon isotope anomaly associated with the earliest Lopingian (Late Permian) mass extinction[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2005, 223(1/2): 172-180. |
| [110] | Shen Shuzhong, Cao Changqun, Zhang Hua, et al. High-resolution δ13Ccarb chemostratigraphy from latest Guadalupian through earliest Triassic in South China and Iran[J]. Earth and Planetary Science Letters, 2013, 375: 156-165. |
| [111] | Wang Wei, Cao Changqun, Wang Yue. The carbon isotope excursion on GSSP candidate section of Lopingian-Guadalupian boundary[J]. Earth and Planetary Science Letters, 2004, 220(1/2): 57-67. |
| [112] | Retallack G J, Jahren A H. Methane release from igneous intrusion of coal during late Permian extinction events[J]. The Journal of Geology, 2008, 116(1): 1-20. |
| [113] | Retallack G J, Metzger C A, Greaver T, et al. Middle-Late Permian mass extinction on land[J]. Geological Society of America Bulletin, 2006, 118(11/12): 1398-1411. |
| [114] | Ishiwatari R, Kon M, Yamada K, et al. Application of gas chromatography/isotope ratio mass spectrometry to petroleum geochemistry: carbon isotopic composition of individual long chain normal alkanes in oils from Northeast Japan[J]. Journal of the Japanese Association for Petroleum Technology, 1999, 64(6): 551-559. |
| [115] | 邱振,王清晨. 广西来宾铁桥剖面中上二叠统沉积微相[J]. 沉积学报,2010,28(5):1020-1036. [Qiu Zhen, Wang Qingchen. middle and upper permian sedimentary microfacies in the Tieqiao Section in Laibin, Guangxi, China[J]. Acta Sedimentologica Sinica, 2010, 28(5): 1020-1036.] |
| [116] | Qiu Zhen, Sun Shu, Wei Hengye, et al. SIMS zircon U-Pb dating from bentonites in the Penglaitan Global Stratotype Section for the Guadalupian-Lopingian boundary (GLB), South China[J]. Geological Journal, 2015, doi: 10.1002/gj.2697. |
| [117] | 邱振. 黔桂盆地中晚二叠世沉积环境变化及其对烃源岩发育的制约[D]. 北京:中国科学院研究生院,2011. [Qiu Zhen. Sedimentary environmental change from the Middle to Late Permian in the Qiangui Basin, and its impact on source rock forming[D]. Beijing: Institute of Geology and Geophysics, CAS, 2011.] |
| [118] | Qiu Zhen, Wang Qingchen. Geochemical evidence for submarine hydrothermal origin of the Middle-Upper Permian chert in Laibin of Guangxi, China[J]. Science China Earth Sciences, 2011, 54(7): 1011-1023. |
| [119] | Nishikane Y, Kaiho K, Henderson C M, et al. Guadalupian-Lopingian conodont and carbon isotope stratigraphies of a deep chert sequence in Japan[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2014, 403: 16-29. |
| [120] | Sigurdsson H. Evidence of volcanic loading of the atmosphere and climate response[J]. Global and Planetary Change, 1990(3), 89: 277-289. |
| [121] | Sigurdsson H. Assessment of the atmospheric impact of volcanic eruptions[M]//Sharpton V L, Ward P D. Global Catastrophes in Earth History. London: Geological Society of America Special Paper, 1990, 247: 99-110. |
| [122] | Genin A, Lazar B, Brenner B. Vertical mixing and coral death in the Red Sea following the eruption of Mount Pinatubo[J]. Nature, 1995, 377(6549): 505-510. |
| [123] | Pinto J P, Turco R P, Toon O B. Self-limiting physical and chemical effects in volcanic eruption clouds[J]. Journal of Geophysical Research, 1989, 94(D8): 11165-11174. |
| [124] | Officer C B, Hallam A, Drake C L, et al. Late Cretaceous and paroxysmal Cretaceous/Tertiary extinctions[J]. Nature, 1987, 326(6109): 143-149. |
| [125] | Cockell C S. Crises and extinction in the fossil record-a role for ultraviolet radiation[J]. Paleobiology, 1999, 25(2): 212-225. |
| [126] | Leavitt S W. Annual volcanic carbon dioxide emission: an estimate from eruption chronologies[J]. Environmental Geology, 1982, 4(1): 15-21. |
| [127] | Gerlach T M, Graeber E J. Volatile budget of Kilauea volcano[J]. Nature, 1985, 313(6000): 273-277. |
| [128] | Berner R A. Examination of hypotheses for the Permo-Triassic boundary extinction by carbon cycle modeling[J]. Proceedings of National Academy of Sciences of United States America, 1985, 99(7): 4172-4177. |
| [129] | Ganino C, Arndt N T. Climate changes caused by degassing of sediments during the emplacement of large igneous provinces[J]. Geology, 2009, 37(4): 323-326. |
| [130] | Thordarson T, Self S, Óskarsson N, et al. Sulfur, chlorine, and fluorine degassing and atmospheric loading by the 1783-1784 AD Laki (Skaftár Fires) eruption in Iceland[J]. Bulletin of Volcanology, 1996, 58(2/3): 205-225. |
| [131] | Vogt P R. Evidence for global synchronism in mantle plume convection, and possible significance for geology[J]. Nature, 1972, 240(5380): 338-342. |
| [132] | Ellis R J. Biochemistry: tackling unintelligent design[J]. Nature, 2010, 463(7278): 164-165. |
| [133] | Pörtner H O. Climate change and temperature dependent biogeography: systemic to molecular hierarchies of thermal tolerance in animals[J]. Comparative Biochemistry and Physiology A, 2002, 132: 739-761. |
| [134] | Axelrod D I. Role of volcanism in climate and evolution[J]. Geological Society of America Special Papers, 1981, 185: 1-60. |
| [135] | Holser W T, Magaritz M. Cretaceous/Tertiary and Permian/Triassic boundary events compared[J]. Geochimica et Cosmochimica Acta, 1992, 56(8): 3297-3309. |
| [136] | Magaritz M, Krishnamurthy R V, Holser W T. Parallel trends in organic and inorganic carbon isotopes across the Permian/Triassic boundary[J]. American Journal of Science, 1992, 292(10): 727-739. |
| [137] | Wang K, Geldsetzer H H J, Krouse H R. Permian-Triassic extinction: organic δ13C evidence from British Columbia, Canada[J]. Geology, 1994, 22(7): 580-584. |
| [138] | Hsü K J, McKenzie J A. A “Strangelove” ocean in the earliest Tertiary[M]//Sundquist E T, Broecker W S. The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present. Washington: American Geophysical Union, 1985, 32: 487-492. |
| [139] | Flügel E, Kiessling W. Patterns of Phanerozoic reef crises[M]//Kiessling W, Flügel E, Golonka J. Phanerozoic Reef Patterns. SEPM. Special Publication, 2002, 72: 691-733. |
| [140] | Rampino M R, Stothers R B. Flood basalt volcanism during the past 250 million years[J]. Science, 1988, 241(4866): 663-668. |
| [141] | Courtillot V. Evolutionary Catastrophics: The Science of Mass Extinction[M]. Cambridge: Cambridge University Press, 1999: 237. |
| [142] | Hallam A, Wignall P B. Mass extinctions and Their Aftermath[M]. Oxford: Oxford University Press, 1997. |
| [143] | Isozaki Y. Permo-Triassic boundary superanoxia and stratified superocean: records from lost deep sea[J]. Science, 1997, 276(5310): 235-238. |
| [144] | Saitoh M, Ueno Y, Isozaki Y, et al. Isotopic evidence for water-column denitrification and sulfate reduction at the end-Guadalupian (Middle Permian)[J]. Global and Planetary Change, 2014, 123(Part A): 110-120. |
| [145] | Limarino C O, Césari S N, Spalletti L A, et al. A paleoclimatic review of southern South America during the late Paleozoic: a record from icehouse to extreme greenhouse conditions[J]. Gondwana Research, 2014, 25(4): 1396-1421. |
| [146] | Fielding C R, Frank T D, Isbell J L, et al. Stratigraphic signature of the late Palaeozoic Ice age in the Parmeener Supergroup of Tasmania, SE Australia, and inter-regional comparisons[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2010, 298(1/2): 70-90. |
| [147] | McLean D M. Mantle degassing unification of the Trans-K-T geobiological record[J]. Evolutionary Biology, 1985, 19: 287-313. |
| [148] | Clapham M E, Payne J L. Acidification, anoxia, and extinction: a multiple logistic regression analysis of extinction selectivity during the Middle and Late Permian[J]. Geology, 2011, 39(11): 1059-1062. |
| [149] | Brenchley P J, Carden G A F, Marshall J D, et al. Environmental changes associated with the “first strike” of the Late Ordovician mass extinction[J]. Modern Geology, 1995, 20(1): 69-82. |
| [150] | Gibbs M T, Barron E J, Kump L R. An atmospheric pCO2 threshold for glaciation in the Late Ordovician[J]. Geology, 1997, 25(5): 447-450. |
| [151] | Sheehan P M. The Late Ordovician mass extinction[J]. Annual Review of Earth and Planetary Sciences, 2001, 29: 331-364. |
| [152] | Sutcliffe O E, Dowdeswell J A, Whitington R J, et al. Calibrating the Late Ordovician glaciation and mass extinction by the eccentricity cycles of Earth’s orbit[J]. Geology, 2006, 28(11): 967-970. |
| [153] | Wei Hengye, Shen Jun, Schoepfer S D, et al. Environmental controls on marine ecosystem recovery following mass extinctions, with an example from the Early Triassic[J]. Earth-Science Reviews, 2015, 149: 108-135. |
| [154] | Isozaki Y, Kawahata H, Ota A. A unique carbon isotope record across the Guadalupian-Lopingian (Middle-Upper Permian) boundary in mid-oceanic paleo-atoll carbonates: the high-productivity “Kamura event” and its collapse in Panthalassa[J]. Global and Planetary Change, 2007, 55(1/2/3): 21-38. |
| [155] | Shen Bing, Xiao Shuhai, Kaufman A J, et al. Stratification and mixing of a post-glacial Neoproterozoic ocean: evidence from carbon and sulfur isotopes in a cap dolostone from northwest China[J]. Earth and Planetary Science Letters, 2008, 265(1/2): 209-228. |
| [156] | Isozaki Y, Aljinovic' D, Kawahata H. The guadalupian (Permian) Kamura event in European Tethys[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2011, 308(1/2): 12-21. |
| [157] | Luo Genming, Huang Junhua, Bai Xiao, et al. Absence of Middle Permian Kamura event in the Paleo-Tethys Ocean[J]. Journal of Earth Science, 2010, 21(1): 86-89. |
| [158] | Kirschvink J L, Isozaki Y, Shibuya H, et al. Challenging the sensitivity limits of Paleomagnetism: Magnetostratigraphy of weakly magnetized Guadalupian-Lopingian (Permian) limestone from Kyushu, Japan[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2015, 418: 75-89. |
| [159] | Hallam A, Wignall P B. Mass extinctions and sea-level changes[J]. Earth-Science Reviews, 1999, 48(4): 217-250. |
| [160] | Wignall P B, Twitchett R J. Oceanic anoxia and the end Permian mass extinction[J]. Science, 1996, 272(5265): 1155-1158. |