 2016, Vol. 43
2016, Vol. 43文章信息
- 人胃癌组织中CLIC4基因表达与微卫星不稳定性的关系
- Expression of CLIC4 in Gastric Carcinoma Tissues and Its Relevance to Microsatellite Instability
- 肿瘤防治研究, 2016, 43(7): 572-576
- Cancer Research on Prevention and Treatment, 2016, 43(7): 572-576
- http://www.zlfzyj.com/CN/10.3971/j.issn.1000-8578.2016.07.006
- 收稿日期: 2015-07-27
- 修回日期: 2015-11-20
引用本文 |

2. 730000 兰州,三爱堂医院病理科
2. Department of Pathology, San Ai Tang Hospital, Lanzhou 730000, China
胃癌的发生发展过程是由多因素引起、多基因参与、多阶段演变的过程。其发病机制涉及一系列遗传学改变,包括癌基因的激活及抑癌基因的失活。微卫星DNA是广泛分布于真核生物基因组中的具有高度多态性的、简单的、短的串联重复序列。数目巨大,在全基因组中其拷贝数可达(3.5~10)×104个。微卫星DNA的基本构成单位是2~6个核苷酸,以二核苷酸(CA)n重复序列最为常见。除二核苷酸串联重复外,还有三、四核苷酸串联重复DNA,但仍以二核苷酸重复序列提供的信息最丰富。微卫星DNA可能对启动子、重组位点或DNA拓扑酶结合位点的调控起作用。近年研究发现,微卫星DNA不稳定性(microsatellite instability, MSI)是导致肿瘤发生、发展的分子机制之一,是肿瘤细胞的又一重要标志[1-2]。大量研究表明,错配修复基因异常可引起肿瘤基因组的不稳定性,从而导致肿瘤易感[3-5]。CLIC4基因定位于人染色体1p36.1, 有六个外显子,编码CLIC4蛋白,是细胞内的一种氯通道[6],作为通道调节剂或信号蛋白参与许多细胞功能,其中包括细胞增殖、细胞凋亡及肿瘤的浸润转移[7-10]。位于1p36.1的D1S234恰好在CLIC4基因上可能会影响CLIC4基因的表达。本研究旨在探索微卫星DNA不稳定在胃癌发生中的作用及其与CLIC4表达的关系。
1 资料与方法 1.1 资料40例胃癌及相应的手术切缘正常组织来自北京大学肿瘤医院/北京肿瘤研究所肿瘤标本库,为2010年5月—9月手术切除标本,手术切除后立即收集癌和手术切缘正常组织,剔除出血坏死组织后,10%福尔马林固定,石蜡包埋,其中高分化腺癌8例、中分化腺癌12例、低分化腺癌15例、黏液腺癌5例,用于微卫星DNA不稳定性检测及CLIC4免疫组织化学检测。另外55例用于CLIC4免疫组织化学检测的胃癌组织为三爱堂医院病理科2011年3月—2013年2月存档石蜡包埋标本,其中高分化腺癌7例、中分化腺癌18例、低分化腺癌20例、黏液腺癌10例。95例胃癌患者中,男55例、女40例,年龄25~78岁,中位年龄55.7岁。所有标本术前均未行放、化疗,同时取距离肿瘤组织 > 5 cm的正常胃黏膜15例,作为免疫组织化学检测的正常对照。DNA提取试剂盒(DP304)为天根生化科技(北京)有限公司产品; CLIC4兔抗人单克隆抗体[EPR14253]ab183043(稀释度为1:150)购于英国Abcam公司,免疫组织化学染色SP试剂盒(SP-9000)购于北京中杉金桥生物技术公司。
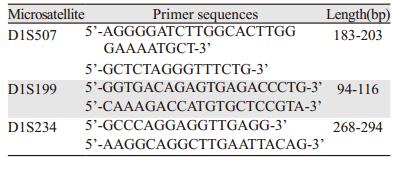
1.2 方法 1.2.1 MSI检测选取位于1p36.1上的D1S234、D1S199、D1S507三个微卫星位点。3对引物由上海生工公司提供,见表 1。采用美国Santa Cruz公司生产的PCR热循环仪扩增微卫星DNA,循环参数如下:95℃3 min,95℃30 s,54℃30 s,72℃30 s,30个循环后72℃延伸10 min。反应总体积为50μl,其中含10×Buffer 5μl,dNTP 4μl,上下游引物各1μl,基因组DNA 5μl,Tap DNA聚合酶1μl,去离子水补至50μl。扩增产物在2%琼脂糖凝胶上电泳证实无杂带后,取5μl扩增产物与3倍体积上样缓冲液混匀,8%的聚丙烯酰胺凝胶电泳,100 V,50℃,约4.5 h,常规银染。
所有标本均经10%甲醛固定,常规石蜡包埋后行4μm连续切片,每例均行HE染色和SP免疫组织化学染色,光学显微镜观察。免疫组织化学操作步骤严格按SP试剂盒说明书进行。一抗浓度为1:150,以PBS代替一抗作阴性对照。
1.3 结果判定如果胃癌组织与其相应的正常组织相比出现条带增加或条带位置发生改变,判断为MSI阳性,如果出现完全一致的电泳条带,判断为MSI阴性,见图 1。用低倍和高倍镜观察全片,CLIC4的阳性染色为在细胞膜或细胞质内出现棕黄色颗粒。

|
| T: gastric cancer tissues; N: normal tissues; 1: MSI negative, identical electrophoresis banding; 2: MSI positive, electrophoresis banding position change; 3: MSI positive, electrophoresis banding increase 图 1 胃癌组织中的MSI判断标准 Figure 1 Judgement standard of microsatellite instability (MSI) in gastric tissues |
实验结果重复三次,应用SPSS11.5统计软件进行数据分析,检验水准为α=0.05,P < 0.05为差异有统计学意义。
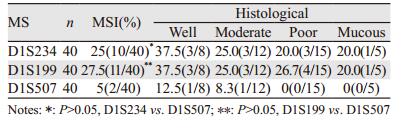
2 结果 2.1 胃癌组织中MSI及其与组织病理类型的关系40例胃癌组织中,D1S234阳性率为25%(10/40),D1S199阳性率为27.5%(11/40),D1S507阳性率为5%(2/40),D1S234和D1S199阳性率明显高于D1S507。各组织病理类型之间差异无统计学意义(P > 0.05),见表 2。
 |
CLIC4在正常胃黏膜中的表达为腺体阳性,间质阴性,见图 2A。40例胃癌组织中,CLIC4在肿瘤实质中阳性10例,阳性率为25%,肿瘤间质中阳性27例,阳性率为67.5%,同时阳性6例,占15%,同时阴性5例,占12.5%。Spearman等级相关分析显示D1S199与CLIC4肿瘤实质阳性率呈正相关(r=0.137, P=0.042),D1S507与CLIC4肿瘤间质阳性率呈负相关(r=-4.22, P=0.009)。
2.3 胃癌组织中CLIC4的表达与临床病理的关系95例胃癌组织中,CLIC4肿瘤间质阳性69例,阳性率为72.3%,肿瘤实质阳性表达49例,阳性率为51.6%,同时阳性表达23例,阳性率为24.2%。肿瘤间质阳性率明显高于肿瘤实质阳性率(χ2=8.945, P=0.003),肿瘤实质阳性病例主要为高-中分化腺癌,见图 2B、2C,肿瘤间质阳性病例主要为低分化及黏液腺癌,见图 2D、2E,随着肿瘤分化程度的降低,CLIC4间质阳性表达率增加,血管阳性表达明显增加,见图 2F。

|
| A: normal gastric mucosa; B: well-differentiated adenocarcinoma; C: moderately-differentiated adenocarcinoma; D: poorly-differentiated adenocarcinoma; E: mucous adenocarcinoma; F: blood vessels 图 2 CLIC4在不同胃组织中的表达(SP×100) Figure 2 Expression of CLIC4 in different gastric tissues (SP×100) |
MSI是指由于复制错误引起的简单重复序列的改变,肿瘤遗传学研究表明,细胞恶性转化与遗传物质的不稳定性有关,而遗传物质的不稳定性系错配修复基因的改变所致,由于错配修复基因的突变造成DNA频发的复制错误,导致细胞微卫星DNA发生改变[11]。CLIC4基因,定位于人类基因组中1p36.11,编码的CLIC4是由253个氨基酸构成的相对分子质量为29 kD的蛋白,CLIC4是CLIC氯通道家族成员中唯一在线粒体上表达的氯通道蛋白[12]。位于1p36.1的D1S234恰好在CLIC4基因上,D1S199、D1S507位于其周围(GenBank),D1S234,D1S199、D1S507三个微卫星位点的不稳定可能会影响CLIC4基因的表达。本研究发现40例胃癌中, D1S234、D1S199、D1S507阳性率分别为25%(10/40)、27.5%及5%(2/40),D1S234和D1S199阳性率明显高于D1S507,各组织病理类型之间差异无统计学意义(P > 0.05)。D1S234、D1S199及D1S507的MSI与胃癌的发生密切相关。
Ponsioen等[13]的研究表明CLIC4的功能活性依赖于其氧化还原状态,活性氧产物能够改变细胞分化相关的基因表达,包括纤维化组织的纤维母细胞向肌纤维母细胞转化、上皮间质转化等。CLIC4可促进VEGF介导的血管内皮细胞的增生和血管腔形成[14-15]。有研究显示,CLIC4参与了卵巢上皮癌肿瘤间质相关成纤维细胞的转化[16]。TGF-β1可以使乳腺组织纤维母细胞的CLIC4转录水平升高,CLIC4在乳腺癌组织的肿瘤间质相关成纤维细胞高表达,而正常组织间质中几乎无表达。来自上皮性卵巢癌肿瘤细胞的旁分泌信号TGF-β1刺激,使纤维母细胞内ROS增加,进而促进CLIC4的表达上调,导致纤维母细胞向肌纤维母细胞转化,即肿瘤相关成纤维细胞的活化[17]。Shukla等[18]研究发现,CLIC4通过TGF-β途径产生肿瘤间质,改变肿瘤微环境,导致肿瘤异质性,促进肿瘤的侵袭与转移。肿瘤相关成纤维细胞活化后形成的肿瘤间质肌纤维母细胞,具有促进上皮恶性转化并永生化的功能,肌纤维母细胞还可以通过分泌纤维母细胞生长因子家族、表皮生长因子家族、转化生长因子家族、胰岛素样生长因子家族等多种生长因子作用于肿瘤上皮细胞, 促进肿瘤细胞增殖[19]。肌纤维母细胞还具有促进肿瘤血管形成的能力,Kojima等[20]的实验表明, 肿瘤间质肌纤维母细胞促进微血管增生的能力比正常纤维母细胞高417倍, 其聚集血管内皮细胞的能力是正常纤维母细胞的412倍,而且认为该作用与肌纤维母细胞分泌的间质细胞源性因子-1(SDF-1/CXCL12)有关, 该因子作用于血管内皮细胞的同源受体CXCR4, 从而聚集血管内皮细胞祖细胞, 促进肿瘤血管形成。阻断ROS或CLIC4的制剂能明显下调与细胞转化相关的促血管因子的产生,即抑制肿瘤的血管生成。还有研究表明,间质CLIC4高表达的肿瘤,预后较差[21]。Deng等[22]报道,CLIC4联合ERp29和Smac检测,可作为预测结直肠癌预后的风险指标。Suh等[23]研究报道,CLIC4可以抑制皮肤鳞状上皮细胞癌的生长。He等[24]发现CLIC4在巨噬细胞中大量表达,稳定的巨噬细胞过度表达CLIC4诱发更多的TNF、IL-6、IL-12等产生,因此CLIC4可以作为一种积极的LPS信号调节器。本研究中,95例胃癌组织中,CLIC4在肿瘤间质阳性69例,阳性率为72.3%,肿瘤实质阳性49例,阳性率为51.6%,同时阳性23例,阳性率为24.2%。肿瘤间质阳性率明显高于肿瘤实质阳性率,肿瘤间质阳性病例主要为低分化及黏液腺癌,肿瘤实质阳性病例主要为高-中分化腺癌,随着肿瘤分化程度的降低,CLIC4间质阳性率增加,提示CLIC4可能参与了胃癌肿瘤间质的成纤维化,促进胃癌的演进。
40例胃癌组织中,CLIC4在肿瘤实质阳性10例,阳性率为25%,肿瘤间质阳性27例,阳性率为67.5%,同时阳性6例,占15%,同时阴性5例,占12.5%。Spearman等级相关分析显示D1S199的MSI与CLIC4肿瘤实质阳性率呈正相关,D1S507的MSI与CLIC4肿瘤间质阳性率呈负相关,而D1S234的MSI与肿瘤实质、间质的阳性率无相关性。由此推理出,D1S199和D1S507的不稳定与胃癌的分化程度呈正相关,与胃癌的发生发展密切相关,其机制有待进一步深入研究。
致谢
本研究部分工作在北京大学临床肿瘤学院/北京大学肿瘤医院完成,得到了季加孚教授及其团队的指导与支持,在此表示深深的感谢!
| [1] | Turaga K, Shibata D. K-Ras and MSI: potential markers of both patient prognosis and treatment efficacy[J]. Ann Surg Oncol, 2010, 17 (2) : 354–5. |
| [2] | Guastadisegni C, Colafranceschi M, Ottini L, et al. Microsatellite instability as a marker of prognosis and response to therapy: a meta-analysis of colorectal cancer survival data[J]. Eur J Cancer, 2010, 46 (15) : 2788–98. |
| [3] | Richman S. Deficient mismatch repair: Read all about it (Review)[J]. Int J Oncol, 2015, 47 (4) : 1189–202. |
| [4] | Thompson BA, Spurdle AB. Microsatellite instability use in mismatch repair gene sequence variant classification[J]. Genes (Basel), 2015, 6 (2) : 150–62. |
| [5] | Li GM. Decoding the histone code: Role of H3K36me3 in mismatch repair and implications for cancer susceptibility and therapy[J]. Cancer Res, 2013, 73 (21) : 6379–83. |
| [6] | Peretti M, Angelini M, Savalli N, et al. Chloride channels in cancer: Focus on chloride intracellular channel 1 and 4 (CLIC1 AND CLIC4) proteins in tumor development and as novel therapeutic targets[J]. Biochim Biophys Acta, 2015, 1848 (10 Pt B) : 2523–31. |
| [7] | He G, Ma Y, Chou SY, et al. Role of CLIC4 in the host innate responses to bacterial lipopolysaccharide[J]. Eur J Immunol, 2011, 41 (5) : 1221–30. |
| [8] | Patel D, Ythier D, Brozzi F, et al. Clic4, a novel protein that sensitizesβ-cells to apoptosis[J]. Mol Metab, 2015, 4 (4) : 253–64. |
| [9] | Argenzio E, Margadant C, Leyton-Puig D, et al. CLIC4 regulates cell adhesion andβ1 integrin trafficking[J]. J Cell Sci, 2014, 127 (24) : 5189–203. |
| [10] | Leanza L, Biasutto L, Managò A, et al. Intracellular ion channels and cancer[J]. Front Physiol, 2013, 4 : 227. |
| [11] | Poulogiannis G, Frayling IM, Arends MJ. DNA mismatch repair deficiency in sporadic colorectal cancer and Lynch syndrome[J]. Histopathology, 2010, 56 (2) : 167–79. |
| [12] | Suh KS, Mutoh M, Nagashima K, et al. The organellular chloride channel protein CLIC4/mtCLIC translocates to the nucleus in response to cellular stress and accelerates apoptosis[J]. J Biol Chem, 2004, 279 (6) : 4632–41. |
| [13] | Ponsioen B, van Zeij L, Langeslag M, et al. Spatiotemporal regulation of chloride intracellular channel protein CLIC4 by RhoA[J]. Mol Biol Cell, 2009, 20 (22) : 4664–72. |
| [14] | Ulmasov B, Bruno J, Gordon N, et al. Chloride intracellular channel Protein-4 functions in angiogenesis by supporting acidification of vacuoles along the intracellular tubulogenic Pathway[J]. Am J Pathol, 2009, 174 (3) : 1084–96. |
| [15] | Tung JJ, Hobert O, Berryman M, et al. Chloride intracellular channel 4 is involved in endothelial Proliferation and morphogenesis in vitro[J]. Angiogenesis, 2009, 12 (3) : 209–20. |
| [16] | Yao Q, Qu X, Yang Q, et al. Blockage of transdifferentiation from fibroblast to myofibroblast in experimental ovarian cancer models[J]. Mol Cancer, 2009, 8 : 78. |
| [17] | Yao Q. CLIC4 participate in epithelial ovarian tumor stromal fibroblasts transformation and its related regulation[D]. Ji' nan: Shandong Da Xue, 2010. [姚勤. CLIC4参与上皮性卵巢癌肿瘤间质相关成纤维细胞的转化及其调控[D].济南:山东大学, 2010.] |
| [18] | Shukla A, Edwards R, Yang Y, et al. CLIC4 regulates TGF-β-dependent myofibroblast differentiation to produce a cancer stroma[J]. Oncogene, 2014, 33 (7) : 842–50. |
| [19] | Yamaguchi H, Sakai R. Direct Interaction between Carcinoma Cells and Cancer Associated Fibroblasts for the Regulation of Cancer Invasion[J]. Cancers (Basel), 2015, 7 (4) : 2054–62. |
| [20] | Kojima Y, Acar A, Eaton EN, et al. Autocrine TGF-beta and stromal cell-derived factor-1 (SDF-1) signaling drives the evolution of tumor-promoting mammary stromal myofibroblasts[J]. Proc Natl Acad Sci U S A, 2010, 107 (46) : 20009–14. |
| [21] | Suh KS, Crutchley JM, Koocherk A, et al. Reciprocal Modifications of CLIC4 n Tumor Epithelium and Stroma Mark Malignant Progression of Multiple human Cancer[J]. Clin Cancer Res, 2007, 13 (1) : 121–31. |
| [22] | Deng YJ, Tang N, Liu C, et al. CLIC4, ERp29, and Smac/DIABLO derived from metastatic cancer stem-like cells stratify prognostic risks of colorectal cancer[J]. Clin Cancer Res, 2014, 20 (14) : 3809–17. |
| [23] | Suh KS, Malik M, Shukla A, et al. CLIC4 is a tumor suppressor for cutaneous squamous cell cancer[J]. Carcinogenesis, 2012, 33 (5) : 986–95. |
| [24] | He G, Ma Y, Chou SY, et al. Role of CLIC4 in the host innate responses to bacterial lipopolysaccharide[J]. Eur J Immunol, 2011, 41 (5) : 1221–30. |