



2. 中国医学科学院北京协和医学院血液学研究所实验血液学国家重点实验室, 天津 300020
2. State Key Laboratory of Experimental Hematology, Institute of Hematology, Beijing Union Medical College & China Academy of Medical Sciences, Tianjin 300020, China
肿瘤坏死因子相关凋亡诱导配体(TRAIL)是TNF家族唯一一个被发现对肿瘤细胞具有诱导凋亡作用,而对正常细胞没有影响的蛋白,其可溶形式(sTRAIL,114-281)现在已进入Ⅱ期临床试验[1]。目前所用的传统化疗药物存在毒性大、临床耐药率高等问题,TRAIL对肿瘤杀伤的选择性、特异性使其有可能成为今后新兴的肿瘤生物治疗“明星”药物,然而进一步的研究显示,一些血液肿瘤及实体瘤,尤其是恶性程度高的肿瘤往往对TRAIL引起的凋亡呈天然耐受[2]。由于TRAIL具有良好的肿瘤靶向性及低毒性,因此,寻找克服TRAIL耐受的机制或增强TRAIL对肿瘤细胞杀伤的策略成为近年来抗肿瘤研究的重点。
乳腺癌是女性最常见的恶性肿瘤之一,居女性恶性肿瘤之首,且患者死亡率居高不下。尽管近年来乳腺癌的预后得到了很大的改善,但乳腺癌对传统化疗药物的多药耐药现象的产生往往代表了不良预后——化疗失败、肿瘤复发和转移。肿瘤的多药耐药指的是肿瘤细胞对结构不同、作用机制相异的传统化疗药物均具有耐药性的现象。由此,开发出新的、可克服耐药的新化疗方案将有助于解决上述问题。PHⅡ-7是靛玉红的衍生物(结构如Fig1),在我们前期的研究中发现,PHⅡ-7对许多不同来源的肿瘤细胞,尤其是耐药的肿瘤细胞有效[3]。在本研究中,我们选择高表达MDR-1的乳腺癌耐药细胞MCF-7/ADR及其敏感细胞MCF-7为研究对象,并以乳腺癌高转移细胞MDA-MB-231及MDA-MB-361为对照,重点探讨PHⅡ-7对sTRAIL诱导的肿瘤细胞凋亡作用的影响。

|
| Fig 1 Structure of indirubin and derivative of indirubin, PHⅡ-7 |
人乳腺癌细胞系MCF-7及其耐药株MCF-7/ADR、转移性乳腺癌细胞系MDA-MB-231、MDA-MB-361均由中国医学科学院血液病研究所惠赠。细胞均培养于含10%灭活胎牛血清的RPMI 1640或DMEM高糖型培养基,37℃、5% CO2恒温培养箱常规培养,隔天换液,取处于对数生长期的细胞用于实验。此外,耐药细胞株MCF-7/ADR常规培养中加入阿霉素(adriamycin,ADR),终浓度为1 mg·L-1以维持耐药性,实验前2周撤药。
1.2 试剂与仪器sTRAIL(PROSPEC,Israel);CCK-8 reagent(日本同仁化学研究所);TRIzol及逆转录试剂(Invitrogen,USA);Real time试剂(大连宝生物公司);阿霉素购自美国辉瑞制药有限公司;BCA蛋白定量试剂盒购自Thermo Scientific 公司;兔抗人caspase-8抗体、兔抗人PARP-1抗体、兔抗人GAPDH抗体、兔抗人H2A.X抗体及兔抗人磷酸化-H2A.X抗体均购自Cell Signaling Technology公司。PCR 引物由Invitrogen公司合成。凝胶电泳仪购自上海天能(Tanon) 公司。
1.3 CCK-8试剂检测细胞活力采用CCK-8试剂测定细胞对药物的敏感性。取对数生长期的肿瘤细胞,用含有10%小牛血清的RPMI 1640培养液配成1×108·L-1,接种于96孔培养板(细胞数为每孔6×103),在37℃、5% CO2条件下培养24 h,分组加药,每个浓度设3个平行孔,实验组加入对应浓度的药物,阴性对照加入等体积的生理盐水,使每孔的终体积为200 μL。培养20 h后,每孔加入CCK-8 20 μL (5 g·L-1),37℃继续培养4 h,在酶标仪上检测450 nm光密度(OD)值。肿瘤细胞生长抑制率按以下公式计算:抑制率/%=(对照组OD值-加药组OD值)/对照组OD值×100%。以同一药物的不同浓度对肿瘤细胞生长抑制率作图可得到剂量反应曲线,根据线性回归方程求出该药物的半数杀伤浓度IC50,即细胞存活率减少50%时的药物剂量[4]。
1.4 人外周血单个核细胞的提取①采正常人外周静脉血约10 mL,经抗凝处理后,按 1∶1 的体积比加入淋巴细胞分离液,静置20 min,1 500 r·min-1离心15 min,用吸管小心吸取白膜层相,尽量不要将其它层面的细胞或液体吸入。②将上面收集到的淋巴细胞用 PBS 洗涤2遍后,以含10%胎牛血清的 RPMI 1640培养基培养2 d,同时加入IL-2(1 000 kU·L-1)刺激细胞。
1.5 活性氧(ROS)检测取处于对数生长期的细胞,稀释接种于6孔板中培养(每孔3×105细胞),对照组(细胞未经药物处理)和实验组(细胞经过药物处理)处理后,胰酶消化细胞,于细胞悬液中加入终浓度10 μmol·L-1 DCFH2-DA染料,避光孵育30 min;孵育结束后重悬、收集细胞,最后用冰预冷的PBS洗涤2次,用流式细胞仪检测细胞内的ROS水平[5]。
1.6 细胞凋亡实验胰酶消化细胞MCF-7/ADR、MCF-7,10%胎牛血清终止消化后PBS洗2遍,制成单细胞悬液,调整细胞浓度为1×108·L-1,6孔板每孔加2 mL细胞悬液,培养过夜后加入药物处理。参照Annexin Ⅴ/PI双染凋亡试剂盒操作说明书操作。收集细胞制成单细胞悬液,冰预冷的PBS洗2遍后,用100 μL Binding buffer将待测细胞的密度调整为5×105~1×106细胞悬液,5 μL FITC-Annexin Ⅴ标记液和5μL PI,轻轻混匀。避光室温染色15 min,再补充400 μL Binding buffer重悬,流式细胞仪(BD,LSRⅡ)测定各组细胞发生凋亡的百分数。实验重复3次,取均值与标准差进行统计分析。
1.7 RNA提取,cDNA合成和实时定量-PCR收集上述乳腺癌细胞,胰酶消化贴壁细胞,10%胎牛血清终止消化后PBS洗2遍,调整细胞浓度,收集2×105个细胞沉淀,以TRIzol裂解。cDNA合成根据Invitrogen cDNA合成试剂盒说明操作。根据GeneBank数据库提供的基因序列信息,采用Premier 5.0软件设计特异性引物,并经BLASÉ分析,由Invitrogen公司合成,PAGE纯化,本研究所用的引物序列如Tab1所示。

收集上述乳腺癌细胞,胰酶消化贴壁细胞,10%胎牛血清终止消化后PBS洗2遍。调整细胞浓度为1×107个,胰酶消化后,按2×106/100μL RIPA浓度加入含有蛋白酶抑制剂的RIPA中裂解,4℃裂解30 min,10 000×g离心20 min,取上清,BCA法进行总蛋白定量。每组取50 μg蛋白,用10%的SDS-PAGE分离蛋白,然后转印至PVDF膜上。5%脱脂牛奶封闭2 h后,按预染标志物标记的分子量裁剪转印膜,分别加入目标蛋白一抗,4℃孵育过夜。再以PBS清洗后加入二抗,室温下孵育30 min,ECL法显色。Tanon凝胶图像分析系统照相并分析结果。
1.9 统计学处理数据结果以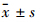 表示,两组均数间比较采用双尾Student’s t检验,多组均数间比较采用方差分析(ANOVA)。所有数据采用SP软件包(SP,Chicago,IL,USA)进行统计学处理。
表示,两组均数间比较采用双尾Student’s t检验,多组均数间比较采用方差分析(ANOVA)。所有数据采用SP软件包(SP,Chicago,IL,USA)进行统计学处理。
不同浓度的PHⅡ-7及阿霉素处理乳腺癌细胞MCF-7及其耐药株MCF-7/ADR 24 h后,其对PHⅡ-7的IC50分别为(4.49±1.55)、(3.44±0.90) μmol·L-1;同时MCF-7及MCF-7/ADR对阿霉素的IC50值分别为(3.45±0.05)和(137±12.77) μmol·L-1,差异具有统计学意义(P<0.05),说明MCF-7/ADR确是耐药的乳腺癌细胞株。PHⅡ-7对乳腺癌细胞MCF-7及MCF-7/ADR的增殖抑制作用均存在剂量依赖性,且当PHⅡ-7浓度为1 μmol·L-1时,细胞抑制率<10%,因此,在以下的实验中选择1μmol·L-1PHⅡ-7为联合用药浓度。
2.2 sTRAIL对乳腺癌细胞的增殖抑制作用乳腺癌高转移细胞MDA-MB-231是文献报道的对TRAIL较敏感的乳腺癌高转移细胞系[6],MDA-MB-361是对TRAIL耐受的乳腺癌高转移细胞,在本实验中我们将它们作为对照。
不同浓度(0~400 μg·L-1)的sTRAIL处理乳腺癌细胞MDA-MB-231、MDA-MB-361、MCF-7及MCF-7/ADR 24h后,与TRAIL敏感细胞MDA-MB-231细胞相比,其他乳腺癌细胞生长抑制率均低于20%,表现出对TRAIL杀伤不敏感。Western blot进一步证实了sTRAIL可有效地诱导TRAIL敏感细胞MDA-MB-231细胞凋亡(Fig2)。

|
| Fig 2 Cytotoxic effect of TRAIL to breast cancer cell line A: Breast cancer cells MDA-MB-231,MDA-MB-361,MCF-7 and its multidrug resistant counterpart MCF-7/ADR were treated by various concentrations of TRAIL(0~400 μg·L-1) for 24 h, then after incubated by CCK-8 reagent for 2 h, the absorbance was read in 450nm and the inhibition rate was calculated. B: Cell lysates containing equal amounts of protein(50μg)were separated by SDS-PAGE and immunoblotted with anti-PARP-1/caspase-8/cleaved caspase-8 antibody. GAPDH was shown as an internal control.*P<0.05,**P<0.01 vs survival rate of 0 concentration of each group |
不同浓度的PHⅡ-7(0~20 μmol·L-1)处理人外周血单个核细胞(peripheral blood mononuclear cells,PBMC),低浓度(<1 μmol·L-1)的PHⅡ-7对PBMC的增殖抑制作用较弱(<80%),即使与50 μg·L-1的sTRAIL联用,对PBMC的抑制作用仍不明显(Fig3A 、B)。

|
| Fig 3 PHⅡ-7 with low concentration augment TRAIL-induced apoptosis in breast cancer cell line A:Cytotoxic effect of PHⅡ-7(0~20μmol·L-1)on human peripheral blood mononuclear cell(PBMC). PBMC was treated with each concentration of PHⅡ-7 for 24h.*P<0.05,**P<0.01 vs control. B: Cell survival rate of PBMC treated by low concentration of PHⅡ-7(0~1μmol·L-1)alone or in combination with sTRAIL(50 μg·L-1) for 24h. C: Cell survival rate of breat cancer cell line MCF-7 and its multidrug resistant counterpat MCF-7/ADR treated by low concentration of PHⅡ-7(0~1μmol·L-1)alone or in combination with sTRAIL(50 μg·L-1) for 24h. *P<0.05 vs PH Ⅱ-7 alone. D: Apoptosis detected in breast cancer cells by Annexin Ⅴ-FITC tests when PHⅡ-7(1μmol·L-1) in combination with sTRAIL(50 μg·L-1). E: Apoptosis rates counted with three independent repeated experiments. *P<0.05 vs control.F: Western blot analysis of apoptosis pathway protein. Cell lysates containing equal amounts of protein(50μg) were separated by SDS-PAGE and immunoblotted with anti-PARP-1 and anti-caspase-8 antibody. GAPDH was shown as an internal control. |
低浓度的PHⅡ-7(0~1 μmol·L-1)与sTRAIL联合处理乳腺癌细胞MCF-7及其耐药株MCF-7/ADR,如Fig3C所示,尽管上述浓度范围内的PHⅡ-7对乳腺癌细胞的增殖抑制作用有限,但一旦与sTRAIL联用,显示出剂量依赖性的增殖抑制作用,其中1 μmol·L-1PHⅡ-7与50 μg·L-1 sTRAIL联用后乳腺癌细胞MCF-7及其耐药株MCF-7/ADR的抑制率分别为(68.03±1.23)%、(60.03±2.44)%。
2.3.3 低浓度的PHⅡ-7与sTRAIL联用可有效诱导乳腺癌细胞凋亡如Fig3D、E所示,1 μmol·L-1PHⅡ-7及sTRAIL(50 μg·L-1)单用均未明显诱导乳腺癌细胞凋亡,而一旦1μmol·L-1PHⅡ-7与sTRAIL联用,细胞凋亡率明显升高,MCF-7及MCF-7/ADR的凋亡率分别为(25.00±3.23)%、(28.18±0.33)%。
2.3.4 低浓度的PHⅡ-7与sTRAIL联用可激活乳腺癌细胞凋亡信号通路Western blot法证实1 μmol·L-1PHⅡ-7与sTRAIL(50 μg·L-1)联用可有效激活caspase-8,诱导乳腺癌细胞凋亡(Fig3F)。
2.4 低浓度的PHⅡ-7通过升高乳腺癌细胞内的活性氧水平上调DR4、DR5的表达 2.4.1 低浓度的PHⅡ-7可激活升高细胞内的活性氧水平利用活性氧的特异探针DCFH2-DA,我们检测了1μmol·L-1 PHⅡ-7作用于MCF-7及MCF-7/ADR后细胞内活性氧的动态变化情况。如Fig4A所示,细胞内的活性氧水平在药物作用2 h后达到最高,之后下降,符合细胞氧化应激的特征。细胞内活性氧水平升高会对细胞内的生物大分子如蛋白质、DNA造成损伤。H2A.X是ATM的效应蛋白,当细胞的DNA受损后,激活ATM/ATR激酶系统,从而将H2A.X磷酸化,因此H2A.X的磷酸化是检测DNA损伤的一个重要指标。如Fig4B所示,乳腺癌细胞中H2A.X的磷酸化状态随着PHⅡ-7作用时间的增加逐渐增强,说明PHⅡ-7引致的ROS升高可能造成了DNA损伤。

|
| Fig 4 1 μmol·L-1 PHⅡ-7 generated ROS production in breast cancer cells A: DCFH2-DA probe detected the ROS production by PHⅡ-7(1μmol·L-1) in breast cancer cells; B: Western blot analysis confirmed DNA damage. Cells were treated with PHⅡ-7(1μmol·L-1) for various time(0-24h). Equal amouts of protein(50μg)from cell lysates were separated by SDS-PAGE and immunoblotted with anti-phospho-H2A.X/ anti-H2A.X antibody. GAPDH was shown as an internal control.*P<0.05 vs control |
实时定量PCR及Western blot分析结果显示,PHⅡ-7可升高乳腺癌细胞内DR4、DR5的表达,且呈剂量依赖性;用ROS的抑制剂NAC处理之后,细胞内DR4、DR5的表达水平恢复至与对照组相当(Fig5)。

|
| Fig 5 1μmol·L-1 PHⅡ-7 up-regulated expression of TRAIL receptor, DR4 and DR5 and ROS scavenger could block the effect A.Realtime PCR analysis confirmed TRAIL receptors, DR4 and DR5 up-regulation by PHⅡ-7(1 μmol·L-1) with various time(0-24h), and ROS scavenger, NAC, blocked the effect. B. Western blot analysis confimed DR4 and DR5 up-regulation by PHⅡ-7(1 μmol·L-1) and once NAC in combination with PHⅡ-7, the expression of DR4 and DR5 remain to control.*P<0.05 vs control |
TRAIL 因其对肿瘤细胞良好的特异性,且对正常细胞毒性很小而显示出很好的应用前景。TRAIL为Ⅱ型跨膜蛋白,其可溶形式(114-281肽段)(soluble TRAIL,sTRAIL)作为跨膜蛋白 TRAIL 胞外区的一部分,可以与完整的TRAIL 分子一样,募集细胞膜表面的功能性受体 DR4、DR5,启动外源性凋亡[7]。然而,尽管TRAIL显示出令人欣喜的抗肿瘤前景,许多肿瘤组织对TRAIL杀伤的耐受也是不容忽视的问题。
有报道表明,c-Fos在前列腺癌组织中,因参与抑制抗凋亡蛋白c-FLIP的功能故可以促进TRAIL介导的细胞凋亡[8, 9, 10]。在我们实验室保存的耐药细胞株K562/A02及MCF-7/ADR也发现c-Fos高表达,并被认为与多药耐药相关蛋白P-gp的过表达相关[4]。受到Zhang等[8]的研究启发,我们比较了sTRAIL分子对耐药及敏感细胞的杀伤,结果发现sTRAIL对它们的杀伤并无差异,说明c-Fos在上述两株耐药细胞株中并未对TRAIL启动的外源性凋亡通路产生明显影响。
尽管部分化疗药物可不同程度地有助于肿瘤细胞克服TRAIL的耐受[11, 12],但这些化疗药物大多对已经获得多药耐药性的肿瘤细胞无效。临床上,常常由于肿瘤细胞高表达P-gp,获得对许多传统化疗药物的多药耐药性而导致化疗失败。这样一来,因为耐药性的产生,这些化疗药物在细胞内不能达到有效的药物浓度,造成这些细胞依旧对与TRAIL联合的用药方案不敏感。同时,由于传统化疗药物在治疗过程中的毒性较大,不仅降低了病人的生活质量,也将对治疗的依从性造成很大的影响。
PHⅡ-7是我们实验室以传统中药当归龙荟丸的有效成分靛玉红为模板,经过构效关系分析合成得到的靛玉红衍生物,我们之前的研究已发现PHⅡ-7可有效地降低多药耐药肿瘤细胞K562/A02、MCF-7/ADR中多药耐药蛋白MDR-1的表达,其可能机制是通过升高细胞内的活性氧水平[5, 13];而多项研究发现,细胞内活性氧水平的高低可影响肿瘤细胞对TRAIL的敏感性[14]。本研究的结果显示,PHⅡ-7正是通过升高细胞内的活性氧水平上调乳腺癌细胞中DR4、DR5的表达水平,进而促进TRAIL对上述细胞的杀伤。尽管多种化疗药物均证实可不同程度地提高肿瘤细胞对TRAIL的敏感性,但上述传统化疗药物对多药耐药的肿瘤细胞无效。且多数情况下,大剂量的化疗药物可以通过不同途径有效杀死肿瘤细胞,但治疗过程中的毒性反应难以避免,开发新的低毒高效的化疗方案一直是肿瘤学界学者们追求的目标。利用TRAIL对肿瘤细胞的选择性,同时借助于低浓度的PHⅡ-7对敏感和多药耐药的肿瘤细胞相当地促进TRAIL杀伤的抗肿瘤作用,低浓度的靛玉红衍生物PHⅡ-7联合TRAIL分子将有希望成为今后肿瘤治疗可选择的有效治疗方案。
(致谢:本实验所用细胞系均为中国医学科学院血液学研究所熊冬生教授惠赠,实验工作主要在中国医学科学院血液学研究所实验血液学重点实验室及南昌大学第一附属医院医学科研中心完成,在此一并感谢。)
| [1] | Lemke J, von Karstedt S, Zinngrebe J, Walczak H. Getting TRAIL back on track for cancer therapy[J].Cell Death Differ,2014,21(9):1350-64. |
| [2] | Sarhan D, D'Arcy P, Lundqvist A. Regulation of TRAIL-receptor expression by the ubiquitin-proteasome system[J].Int J Mol Sci,2014,15(10):18557-73. |
| [3] | Su Y, Chen X, Tan Y H,et al. Synthesis of a dual functional anti-MDR tumor agent PHⅡ-7 with elucidations of anti-tumor effects and mechanisms[J]. PLoS One,2012,7(3):e32782. |
| [4] | 彭洪薇,师锐赞,袁向飞,等.乳腺癌耐药细胞中c-fos抗凋亡作用的研究[J].中国癌症杂志,2014,24(8):581-8. Peng H W, Shi R Z, Yuan X F, et al. The anti-apoptotic effect of c-fos in drug-resistant breast cancer cells[J]. China Oncol, 2014, 24(8):581-8. |
| [5] | 彭洪薇,袁向飞,李真真,等.PHⅡ-7通过升高ROS诱导K562和K562/A02凋亡[J].中国药理学通报,2012,28(7):911-6. Peng H W, Yuan X F, Li Z Z, et al. PHⅡ-7 induces apoptosis on K562 and K562/A02 by increasing ROS production[J]. Chin Pharmacol Bull, 2012, 28(7):911-6. |
| [6] | 仇凤启,赵 丹,孙大鹏,等.5-Aza-2'-dC通过上调DR4与DR5表达增加TRAIL诱导乳腺癌细胞凋亡的敏感性[J].中国医科大学学报,2012,41(12):1102-5. Qiu F Q, Zhao D, Sun D P, et al. 5-Aza-2'-dC enhances TRAIL-induced apoptosis of breast cancer cells by inducing DR4 and DR5 expression[J]. J China Med Univ, 2012, 41(12):1102-5. |
| [7] | Holland P M. Targeting Apo2L/TRAIL receptors by soluble Apo2L/TRAIL[J]. Cancer Lett,2013,332(2):156-62. |
| [8] | Zhang X, Zhang L, Yang H, et al. c-Fos as a proapoptotic agent in TRAIL-induced apoptosis in prostate cancer cells[J]. Cancer Res, 2007,67(19):9425-34. |
| [9] | Horak P, Pils D, Kaider A, et al. Perturbation of the tumor necrosis factor-related apoptosis-inducing ligand cascade in ovarian cancer:overexpression of FLIP(L) and deregulation of the functional receptors DR4 and DR5[J].Clin Cancer Res,2005,11(24):8585-91. |
| [10] | Abdulghani J, El-Deiry W S. TRAIL receptor signaling and therapeutics[J]. Expert Opin Ther Targets, 2010,14(10):1091-108. |
| [11] | von Haefen C, Gillissen B, Hemmatti P G, et al. Multidomain Bcl-2 homolog Bax but not Bak mediates synergistic induction of apoptosis by TRAIL and 5-FU through the mitochondrial apoptosis pathway[J].Oncogene,2004,23(50):8320-32. |
| [12] | Hwang I T, Chung Y M, Kim J J, et al. Drug resistance to 5-FU linked to reactive oxygen species modulator 1[J].Biochem Biophys Res Commun,2007,359(2):301-10. |
| [13] | Peng H W, Yuan X F, Shi R Z, et al. PHⅡ-7 inhibits cell growth and induces apoptosis in leukemia cell line K562 as well as its MDR-counterpart K562/A02 through producing reactive oxygen species[J]. Europ J Pharmacol,2013,718(1-3):459-68. |
| [14] | Mellier G, Pervaiz S. The three Rs along the TRAIL:resistance, re-sensitization and reactive oxygen species(ROS)[J].Free Radic Res,2012,46(8):996-1003. |