



2. 福建医科大学神经生物学研究中心, 福建 福州 350108;
3. 福建医科大学附属第一医院, 福建 福州 350004;
4. 福建省神经病学研究所, 福建 福州 350005
2. Center for Neuroscience Research, Fujian Medical University, Fuzhou 350108, China;
3. the First Affiliated Hospital of Fujian Medical University, Fuzhou 350004, China;
4. Fujian Provincial Institute of Neurology, Fuzhou 350005, China
帕金森病(PD)的主要病变是脑部黑质含黑色素的神经元大量死亡,以致纹状体内神经递质多巴胺(DA)减少。酪氨酸羟化酶(TH)是多巴胺合成代谢的限速酶,胶质源性神经营养因子(GDNF)对DA能神经元具有特异性营养作用,在纹状体同时导入TH和GDNF基因并且能被调控表达是PD基因治疗的理想方法[1, 2]。用于调控哺乳动物细胞基因表达的Tet-On系统虽表现出很好应用前景,如诱导剂强力霉素(Dox)高效、无毒,在时间上准确调控外源基因表达等 [2],但亦有不足,主要体现在相对高的基础活动 [3, 4, 5, 6]。Tet-On系统由两部分组成,四环素调控的反式作用子(rtTA)与四环素应答元件(TRE)。研究表明,Tet-On系统缺陷与rtTA结构不完善[3, 4, 5, 6]以及TRE启动子人巨细胞病毒最小启动子(PminCMV)自身激活[3, 7]有关。在前期实验里,我们采用改良Tet-On修饰的慢病毒载体系统,即rtTA2s-M2[4, 8]替代rtTA;小鼠白蛋白启动子 (Palb)[3] 替代PminCMV,把TH和GDNF双治疗基因通过一个内部核糖体进入位点(IRES)构建于Palb的下游,获得重组慢病毒穿梭质粒pRRL-Palb-TH- IRES-GDNF,并包装成重组慢病毒Lv-TH-GDNF [1]。在本实验里,我们用获取的Lv-TH-GDNF与rtTA2s-M2病毒感染HeLa细胞,以及行PD大鼠纹状体内直接转移,拟通过体内、体外实验观察Dox对Lv-TH-GDNF目的基因的调控表达以及Lv-TH-GDNF对PD模型大鼠的治疗效应。
1 材料与方法 1.1 材料与试剂Lv-TH-GDNF慢病毒(滴度:2.1×1011 TU·L-1)、rtTA2s-M2病毒(滴度:2.01×1011 TU·L-1)、HeLa细胞(均本实验组提供);6-羟基多巴胺(6-OHDA)、维生素C、阿扑吗啡(APO)、多巴胺(DA)、3,4-二羟基苯乙酸(DOPAC)、兔抗大鼠TH单克隆抗体(Sigma公司);兔抗大鼠 GAPDH单克隆抗体、羊抗大鼠GDNF单克隆抗体(Abcam公司);SP免疫组化二抗试剂盒-通用型(上海麦约尔生物技术有限公司);其他均为进口或国产试剂。
1.2 实验动物SD大鼠,♂,体质量250~280 g,由福建医科大学实验动物中心提供。
1.3 Dox对Lv-TH-GDNF感染HeLa细胞后TH、GDNF蛋白表达调控 1.3.1 慢病毒感染HeLa细胞分组:HeLa 组、HeLa+Lv-TH-GDNF组、HeLa+Lv-TH-GDNF+rtTA2s-M2组、HeLa+Lv-TH-GDNF+rtTA2s-M2+Dox(5 mg·L-1)组。HeLa细胞以1× 105接种于6孔板,用高糖DMEM完全培养液培养。HeLa细胞组加入新鲜培养液。病毒组是将各病毒以20的MOI值感染HeLa细胞,12 h后更换新鲜培养液。病毒+Dox组是病毒感染48 h后加入Dox (5 mg·L-1),继续孵育48 h,收集细胞用于免疫印迹法测定TH、GDNF基因表达。
1.3.2 免疫印迹法测定TH、GDNF基因表达收集各组细胞,加入细胞裂解液抽提蛋白,按照实验操作说明分别进行蛋白定量、十二烷基硫酸钠聚丙烯酰胺(SDS-PAGE)凝胶电泳、转膜、封闭。随后免疫反应中一抗分别用兔抗大鼠TH单克隆抗体(工作浓度1 ∶ 1 000)或羊抗大鼠GDNF单克隆抗体(工作浓度1 ∶ 250),对照用兔抗大鼠GAPDH 单克隆抗体(工作浓度1 ∶ 250)。与二抗杂交、洗膜等操作后,将膜与化学发光底物室温孵育,曝光胶片,显影定影。
1.4 Lv-TH-GDNF对6-OHDA损毁多巴胺能神经元的治疗效应 1.4.1 完全损毁PD大鼠模型[9]将6-OHDA 8 μg(溶于0.2%抗坏血酸的生理盐水,4 g·L-1)注射到SD大鼠右侧内侧前脑束(MFB)。MFB坐标[9]AP:-4.3mm,ML:-1.5 mm,DV:-7.5 mm。注射速度1 μL·min-1,注射完毕留针2 min后缓慢退针,缝合皮肤,腹腔注射青霉素(0.5毫升/只)。手术第4周用APO(0.5mg·kg-1)皮下注射测转,>5圈/分钟为模型成功。
1.4.2 实验动物分组及实验实施选取造模成功的SD大鼠52只,随机分成病毒+Dox组13只、病毒组13只、PBS对照组13只和健康对照组13只。成功造模的第5周,病毒组每只动物右侧纹状体同时注射等量的Lv-TH-GDNF、rtTA2s-M2病毒(各2 μL),饮清水。病毒+Dox组接受同等量的病毒注射,给予含强力霉素(0.2 g·L-1)和蔗糖(40 g·L-1)的饮用水。PBS对照组接受等体积的 PBS注射,饮清水。健康对照组不做注射,饮清水。纹状体注射部位的坐标:AP:+1.0 mm,ML: -3.0 mm,DV: -5.5 mm。病毒注射后4周,每组大鼠行APO诱发旋转行为观察。病毒注射后6周,每组取5只大鼠行黑质TH免疫组化;每组取5只大鼠行右侧纹状体单胺类神经递质DA、DOPAC含量的检测;每组取3只大鼠的右侧纹状体,通过免疫印迹法检测GDNF和TH基因表达。
1.4.3 旋转行为观察APO(溶于0.1%抗坏血酸的生理盐水)以0.5 mg·kg-1皮下注射诱发大鼠旋转,用旋转行为测试系统记录。
1.4.4 黑质(SN)TH阳性细胞检测用免疫组化方法。动物灌注固定,大脑行腹侧中脑黑质冠状系列冰冻切片,30 μm厚,收集黑质切片于PBS缓冲液。方法按照SP免疫组化二抗试剂盒-通用型说明进行。每张切片的一抗为兔抗大鼠 TH单克隆抗体(工作浓度1 ∶ 1 000),阴性对照组以PBS缓冲液代替一抗),常规脱水、透明、封片、镜检。计算损伤侧与健侧的黑质致密部(SNpc)TH阳性细胞数的百分比。
1.4.5 移植侧纹状体DA、DOPAC含量检测用高效液相色谱-电化学方法(HPLC-ECD)测定。大鼠断头取脑,冰上分离右侧纹状体,称重后置-70℃冰箱备用。检测标本处理方法参照文献[10]。色谱条件:Hypersil ODS-3柱(250 mm×4.6 mm id),保护柱(10 mm×4.6 mm id),工作电极为玻璃碳电极,参比电极Ag/AgCl,电化学监测器设定电压为+650 mV。流动相为缓冲液(0.02 mol·L-1柠檬酸三钠和0.05 mol·L-1磷酸氢二钠) ∶ 甲醇=90 ∶ 10,流速为1.0 mL·min-1。实验常温进行。以标准品保留时间作为定性指标。
1.4.6 移植侧纹状体TH、GDNF基因表达检测用免疫印迹法测定。大鼠断头取脑,在移植区注射点附近取100 mg的纹状体加入全细胞裂解液匀浆,蛋白定量。取40 μg总蛋白上样,具体见方法1.3.2。用Quantity One软件进行灰度分析。
1.4.7 统计学处理实验数据以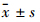 表示,采用SPSS 11.5统计软件做t检验。
表示,采用SPSS 11.5统计软件做t检验。
病毒感染HeLa细胞实验,免疫印迹法检测结果(Fig1):Dox阳性组见TH、GDNF蛋白条带,阴性各组未见目的蛋白条带。

|
| Fig 1 TH and GDNF protein in HeLa cells examined by Western blot 1:HeLa group; 2:HeLa+Lv-TH-GDNF group; 3:HeLa+Lv-TH-GDNF+rtTA2s-M2 group; 4:HeLa+Lv-TH-GDNF+rtTA2s-M2+Dox(5 mg·L-1)group. GAPDH:intrinsic control. |

|
| Fig 2 Time course of experiment |
完全损毁PD大鼠模型,观察到动物少食少动、尾部强直,有时有自发性旋转。在病毒或PBS移植注射后4周,APO能诱导PD模型大鼠向左旋转(Fig3)。记录大鼠60 min平均旋转次数,实验组与各自移植前相比,或者移植后与PBS对照组相比,仅病毒+Dox组均明显降低(P<0.01)。

|
| Fig 3 Cumulative apomorphine-induced contralateral rotations(n=13) **P<0.01 vs PBS-treated groups; △△P<0.01 vs pre-treated group |
移植后6周,黑质TH免疫组化图片显示:TH阳性细胞被染成棕褐色,PBS对照组损伤侧的TH阳性细胞变性、消失,数目明显少于健侧(Fig4)。计算损伤侧与健侧SNpc的TH阳性细胞数的百分比(Fig5),与健康对照组比较,各实验组均明显下降(P<0.01);在PD模型鼠中,与PBS对照组比较,仅病毒+Dox组明显升高(P<0.01)。

|
| Fig 4 Photomicrographs of TH immunohistochemistry in SN A~B:Photomicrographs of TH-positive cells in intact-side(A)or lesion-side(B) SNpc of a PBS-treated rat(×200); C:The SN of a non-injected-treated rat(×10); D:The SN of a PBS-treated rat(×10). |

|
| Fig 5 Number of TH-positive cells in SNpc(n=5) **P<0.01 vs PBS-treated groups; △△P<0.01 vs non-injected-treated groups |
移植后6周,HPLC-ECD检测结果(Fig6):与健康对照组比较,各实验组损伤侧纹状体DA、DOPAC含量明显降低(P<0.01)。在PD模型鼠中,与PBS对照组比较,仅病毒+Dox组数值明显升高(P<0.01)。单位以每g脑组织湿重所含单胺类递质质量mg表示。

|
| Fig 6 Content of DA and DOPAC in lesion-side striatum measured by HPLC-ECD(n=5) A:DA;B:DOPAC. **P<0.01 vs PBS-treated groups;△△P<0.01 vs non-injected-treated groups |
根据纹状体移植区TH、GDNF免疫印迹结果及灰度表明(Fig7),在PD模型鼠中,与PBS对照组比较,TH、GDNF蛋白量在病毒+Dox组均明显增高(P<0.01),而病毒组差异无显著性(P>0.05)。

|
| Fig 7 Expression of TH and GDNF genes of tissue near injection site in striatum analyzed by Western blot 1:Lv-TH-GDNF+rtTA2s-M2-treated groups; 2:PBS-treated groups; 3:Lv-TH-GDNF+rtTA2s-M2+Dox-treated groups. GAPDH:intrinsic control. |
Tet-On系统是在1995年Gossen等[11]成功构建,专被用于调控哺乳动物细胞的基因表达。其rtTA由反式四环素阻遏蛋白(rtetR)与单纯疱疹病毒的VP16 转录活化域融合而成,其TRE由四环素操纵序列(TetO)与下游的人PminCMV融合而成。但该系统存在不足,首先是诱导基因的不稳定表达[4, 5]。其次,系统激活效率较低[4, 5, 12]。最后是系统最显著的缺陷:相对高的基础活动[3, 4, 5, 6],即诱导剂不存在时,rtTA也会与TRE结构中TetO结合,诱导外源基因转录。研究表明,系统缺陷与rtTA结构不完善[3, 4, 5, 6]以及PminCMV自身激活[3, 7]有关。rtTA结构不完善主要体现为VP16的毒性作用干扰压制细胞的正常转录活动[3, 4];结构中的原核密码子、隐性剪接供体和受体位点可能造成其在真核细胞翻译成无活性蛋白,二者都将导致系统诱导基因不稳定性表达[4, 5] 。2000年Urlinger等[4]在rtTA基础上构建一个新型Tet-On反式激活因子rtTA2s-M2。Urlinger等通过改造rTetR的结构以及VP16转录活化域,降低了VP16毒性,提高系统诱导基因表达的稳定性、对Dox的敏感性以及明显的低背景活动。此外,为了降低Tet-On系统的背景活动,Zabala等[3]用一个片段为196bp的肝特异性表达的小鼠Palb做为TRE的启动子,背景效应也明显下降。
在前期实验中,我们获得改良Tet-On系统修饰的Lv-TH-GDNF和rtTA2s-M2病毒。在本次实验里,Lv-TH-GDNF与rtTA2s-M2病毒以相同 MOI值感染HeLa细胞,免疫印迹显示,仅Dox阳性组可见GDNF、TH蛋白条带,阴性组未见(如Dox不存在或者只有慢病毒 Lv-TH-GDNF单独感染细胞),说明改良Tet-On系统能调控目的基因表达,在离体细胞实验未见基础活动。
在整体实验,我们建立完全损毁PD大鼠模型,该模型90%的黑质多巴胺能神经元和纹状体纤维受损[9],我们将Lv-TH-GDNF与rtTA2s-M2病毒共同注射到PD大鼠患侧纹状体,用Dox诱导TH、GDNF基因表达。移植后4周,在PD模型鼠中,通过与PBS对照组比较,我们观察到仅有病毒+Dox组大鼠,APO诱导旋转行为能得到明显改善(P<0.01)。与此一致,移植后6周,病毒+Dox组大鼠损伤侧(移植侧)黑质致密部TH阳性细胞数、纹状体DA、DOPAC含量均明显增加(P<0.01),说明在Dox诱导下Lv-TH-GDNF能减缓6-OHDA诱导DA能神经元的变性。随后,我们通过免疫印迹法检测纹状体移植区TH、GDNF基因表达,结果显示,与PBS对照组相比,病毒+Dox组TH、GDNF的蛋白量均明显增高(P<0.01),说明移植是成功的,在Dox诱导下,rtTA2s-M2与Lv-TH-GDNF结构中的TRE结合,激活TRE启动子Palb,使外源基因GDNF和TH表达。由于Tet-On系统最显著的缺陷是相对高的基础活动,我们根据以往研究结果[13, 14]:单侧黑质纹状体TH阳性细胞或DA含量丢失达90%以上,才出现药物诱发的不对称行为。结合本实验采用的PD大鼠模型,有90%的黑质多巴胺能神经元和纹状体纤维受损,而在病毒移植后,与PBS对照组相比,无诱导剂的病毒组APO诱发的旋转行为无改善、移植区纹状体TH、GDNF蛋白量均无明显增加的实验结果,我们虽不能肯定改良的Tet-On系统调控的基因表达在体内不存在基础活动,但可以推测这种基础活动是极其微弱的。
综上所述,新型Tet-On修饰Lv-TH-GDNF纹状体内直接转移,在Dox诱导下对PD大鼠有一定治疗作用,且目的基因表达在体外不存在基础活动。
| [1] | 张阳, 张志坚, 杨光华, 等. 新型Tet-On系统大鼠GDNF和TH双基因的慢病毒载体的构建与表达[J].中国药理学通报, 2010, 26(8):1079-84. Zhang Y, Zhang Z J, Yang G H, et al. Construction of lentiviral vectors containing rat GDNF and TH bi-gene and expression by improved Tet-On system[J]. Chin Pharmacol Bull, 2010, 26(8):1079-84. |
| [2] | Yang W H, Yang C, Xue Y Q, et al. Regulated expression of lentivirus-mediated GDNF in human bone marrow-derived mesenchymal stem cells and its neuroprotection on dopaminergic cells in vitro[J].PLoS One, 2013, 8(5):e64389. |
| [3] | Zabala M, Wang L, Hernandez Alcoceba R, et al. Optimization of the Tet-On system to regulate interleukin 12 expression in the liver for the treatment of hepatic tumors[J]. Cancer Res, 2004, 64(8):2799-804. |
| [4] | Urlinger S, Baron U, Thellmann M, et al. Exploring the sequence space for tetracycline-dependent transcriptional activators:novel mutations yield expanded range and sensitivity[J]. Proc Natl Acad Sci USA, 2000, 97(14):7963-8. |
| [5] | Qu Z, Thottassery J V, Van Ginkel S, et al. Homogeneity and long-term stability of tetracycline-regulated gene expression with low basal activity by using the rtTA2S-M2 transactivator and insulator-flanked reporter vectors[J]. Gene, 2004, 327(1):61-73. |
| [6] | Liu Q, Hill P J, Karamitri A, et al. Construction of a doxycycline inducible adipogenic lentiviral expression system[J]. Plasmid, 2013, 69(1):96-103. |
| [7] | Kramer M G, Barajas M, Razquin N, et al. In vitro and in vivo comparative study of chimeric liver-specific promoters[J]. Mol Ther, 2003, 7(3):375-85. |
| [8] | Michalon A, Koshibu K, Baumgärtel K, et al. Inducible and neuron-specific gene expression in the adult mouse brain with the rtTA2S-M2 system[J]. Genesis, 2005, 43(4):205-12. |
| [9] | 韩济生.神经科学[M]. 第3版. 北京:北京大学医学出版社, 2009:1086. Han J S. Neuroscience[M]. 3rd ed. Beijing:Peking University Medical Press, 2009:1086. |
| [10] | 阮国洪, 郑力行, 吴强恩, 等. 血清及脑组织中单胺类神经递质高效液相色谱电化学检测方法研究[J].卫生研究, 2006, 35(2):146-9. Ruan G H, Zheng L X, Wu Q E, et al. Study on monoamine neurotransmitters in rat serum and brain tissue by high performance liquid chromatography with electrochemical detection[J]. J Hygiene Res, 2006, 35(2):146-9. |
| [11] | Gossen M, Freundlieb S, Bender G, et al. Transcriptional activation by tetracyclines in mammalian cells[J]. Sci, 1995, 268(5218):1766-9. |
| [12] | Sato T, Muroyama Y, Saito T. Inducible gene expression in postmitotic neurons by an in vivo electroporation-based tetracycline system[J]. J Neurosci Methods. 2013, 214(2):170-6. |
| [13] | Schwarting R K, Huston J P. The unilateral 6-hydroxydopamine lesion model in behavioral brain research. Analysis of functional deficits, recovery and treatments[J]. Prog Neurobiol, 1996, 50(2-3):275-331. |
| [14] | Simola N, Morelli M, Carta A R. The 6-hydroxydopamine model of Parkinson's disease[J]. Neurotox Res, 2007, 11(3-4):151-67. |