 2025, Vol. 36
2025, Vol. 36扩展功能
文章信息
- 郭忱然, 韩笑, 刘雨秋, 任佳荣, 王晓旭, 常楠, 冀好强, 汪子豪, 刘起勇
- GUO Chen-ran, HAN Xiao, LIU Yu-qiu, REN Jia-rong, WANG Xiao-xu, CHANG Nan, JI Hao-qiang, WANG Zi-hao, LIU Qi-yong
- 2012-2021年内蒙古锡林郭勒盟达乌尔黄鼠密度及其体表寄生蚤时空分布特征
- Spatiotemporal distribution characteristics of the density and ectoparasitic fleas of Spermophilus dauricus in Xilin Gol League, Inner Mongolia Autonomous Region, China, 2012-2021
- 中国媒介生物学及控制杂志, 2025, 36(2): 215-225
- Chin J Vector Biol & Control, 2025, 36(2): 215-225
- 10.11853/j.issn.1003.8280.2025.02.013
-
文章历史
- 收稿日期: 2024-11-25




2 传染病溯源预警与智能决策全国重点实验室, 中国疾病预防控制中心传染病预防控制所媒介生物控制室, 世界卫生组织媒介生物监测与管理合作中心, 北京 102206;
3 山东大学齐鲁医学院公共卫生学院媒介生物控制学系, 山东 济南 250012
2 National Key Laboratory of Intelligent Tracking and Forecasting for Infectious Diseases, Department of Vector Biology and Control, National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention, WHO Collaborating Centre for Vector Surveillance and Management, Beijing 102206, China;
3 Department of Vector Control, School of Public Health, Cheeloo College of Medicine, Shandong University, Jinan, Shandong 250012, China
鼠疫是一种传播速度快、传染性强、危害严重的鼠源疾病[1],鼠类种群密度及其体表寄生蚤变化对鼠疫动态及流行有着重要的预测预警作用[2-4]。松辽平原察哈尔丘陵达乌尔黄鼠(Spermophilus dauricus)鼠疫自然疫源地、内蒙古高原长爪沙鼠(Meriones unguiculatus)鼠疫自然疫源地以及锡林郭勒高原布氏田鼠(Lasiopodomys brandtii)鼠疫自然疫源地是发生鼠疫风险较高的地区,这3大类鼠疫自然疫源地交叉重叠于内蒙古自治区(内蒙古)锡林郭勒盟所辖范围内12个旗(市、县)[5]。达乌尔黄鼠是松辽平原察哈尔丘陵达乌尔黄鼠鼠疫自然疫源地的主要宿主动物[6],是内蒙古高原长爪沙鼠鼠疫自然疫源地的次要宿主动物[7-9],对鼠疫传播具有重要影响,了解其密度变化及其体表寄生蚤时空分布情况对鼠疫监测与防控时间关口的前移、重点区域的确定具有重要意义[5, 10-11]。
本研究通过收集2012-2021年内蒙古锡林郭勒盟地区监测数据,描述达乌尔黄鼠体表寄生蚤构成,分析内蒙古锡林郭勒盟达乌尔黄鼠密度、体表主要寄生蚤及寄生蚤群落多样性的时间分布特征及其在锡林郭勒盟不同鼠疫自然疫源地的空间分布特征,旨在为鼠疫监测与防控策略的制定提供参考,以降低鼠疫发生风险。
1 材料与方法 1.1 研究地区概况锡林郭勒盟地处北纬42°32′~46°41′,东经111°59′~120°00′,行政区划面积20.26万km2,辖2个市、9个旗、1个县和1个管理区。地形以高平原为主体,兼有多种地貌单元,地势南高北低,自西南向东北倾斜。西部和北部地形平坦,东南部多低山丘陵,盆地错落其间,形成广阔的高原草场,见图 1。锡林浩特市、阿巴嘎旗、东乌珠穆沁旗和西乌珠穆沁旗属于布氏田鼠鼠疫自然疫源地,镶黄旗、正镶白旗、正蓝旗、多伦县属于达乌尔黄鼠鼠疫自然疫源地,二连浩特市、苏尼特左旗、苏尼特右旗、太仆寺旗、镶黄旗和正镶白旗属于长爪沙鼠鼠疫自然疫源地,其中镶黄旗、正镶白旗为达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区[5]。

|
| 图 1 研究地区内蒙古自治区锡林郭勒盟行政区划图 Figure 1 Administrative divisions of Xilin Gol League, Inner Mongolia Autonomous Region in this study |
| |
2012-2021年锡林郭勒盟12个旗(市、县)的达乌尔黄鼠密度及其体表寄生蚤相关数据均来自于中国疾病预防控制信息系统鼠疫防治管理信息系统,包含鼠密度、检蚤鼠数、染蚤鼠数、获蚤数、染蚤率、(总/某种蚤)蚤指数、获蚤种类。
1.3 研究方法 1.3.1 指标计算选用鼠密度、染蚤率、(总/某种蚤)蚤指数反映达乌尔黄密度及其体表寄生蚤感染情况。选用Shannon⁃Wiener多样性指数(H')及Simpson优势度指数(D)对达乌尔黄鼠体表所携带寄生蚤的群落结构多样性进行分析。各指标定义与计算如下:

|
式中S为达乌尔黄鼠体表寄生蚤物种数,Pi为达乌尔黄鼠第i种体表寄生蚤个体数(Ni)与体表寄生蚤总数(N)的比值。
1.3.2 统计分析用R 4.3.1软件计算达乌尔黄鼠体表寄生蚤群落结构多样性。使用Excel 2021软件对各个数据汇总整理,分别按年、年月、疫源地类型进行统计与描述。
使用Mann-Kendall时间趋势检验分析鼠密度、鼠体染蚤率、总蚤指数、方形黄鼠蚤(Citellophilus tesquorum)蚤指数、光亮额蚤(Frontopsylla luculenta)蚤指数、阿巴盖新蚤(Neopsylla abagaitui)蚤指数、多样性指数、优势度指数的年际趋势;使用季节性分解(seasonal decomposition of time series,STL)模型分解其趋势分量、季节周期分量及残差分量,分析年月时间序列的季节趋势。由于达乌尔黄鼠是冬眠动物,可以监测到其活动的月份多集中于4-10月,因此将STL模型的周期参数设置为7。
描述锡林郭勒盟境内不同鼠疫自然疫源地类型的达乌尔黄鼠密度及其寄生蚤感染情况,并利用Kruskal-Wallis H秩和检验分析不同疫源地间的整体分布差异,利用Dunn检验进行秩和检验后的两两比较,并通过Bonferroni法调整P值。对不同鼠疫自然疫源地的达乌尔黄鼠密度及其寄生蚤感染情况使用Mann-Kendall时间趋势检验分析其年际变化。
2 结果 2.1 整体情况描述2012-2021年锡林郭勒盟捕获达乌尔黄鼠共13 987只,平均密度为1.29只/hm2;带蚤鼠数为8 603只,鼠体染蚤率为61.51%;获体表寄生蚤共4科11属17种48 002只,总蚤指数为3.43。其中,方形黄鼠蚤38 084只,为优势蚤种,蚤指数为2.72,其次为光亮额蚤4 444只、阿巴盖新蚤3 986只,二者蚤指数分别为0.32和0.28。上述3种寄生蚤数量合计占全部寄生蚤数量的96.90%,其他蚤种数量相对较少,蚤指数较低,见表 1。达乌尔黄鼠体表寄生蚤群落的D=0.36,H'=0.76。
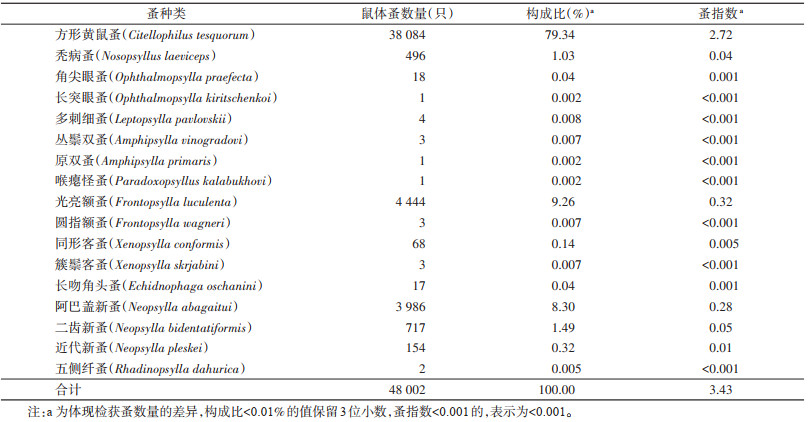
|
从年际趋势看,2013年平均鼠密度最高,为1.74只/hm2,2019年最低,仅为1.08只/hm2。鼠体染蚤率以2012年最高,为86.71%,2013年最低, 仅为53.32%。总蚤指数2012年最高,为5.24,2019年最低, 为2.47。方形黄鼠蚤蚤指数与总蚤指数的年际趋势一致,在2012年最高,2019年最低;光亮额蚤蚤指数在2014年最高,2019年最低;阿巴盖新蚤蚤指数在2012年最高,2021年最低。多样性指数与优势度指数均在2012年最高,2015年最低,二者趋势相似。见表 2。
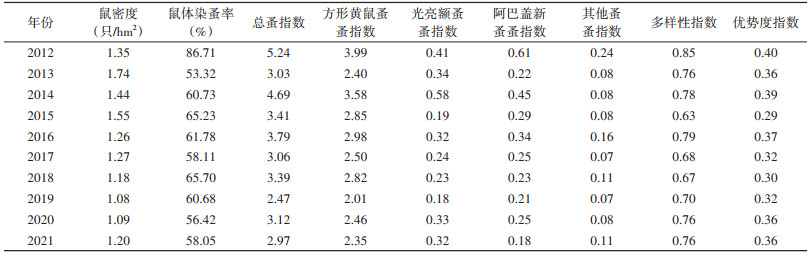
|
Mann-Kendall时间趋势检验分析显示锡林郭勒盟2012-2021年间鼠密度、总蚤指数、方形黄鼠蚤蚤指数、阿巴盖新蚤蚤指数均呈现下降趋势,结果均有统计学意义(均Z < 0,均P < 0.05)。而鼠体染蚤率、光亮额蚤蚤指数、多样性指数与优势度指数的变化趋势均无统计学意义。见表 3。
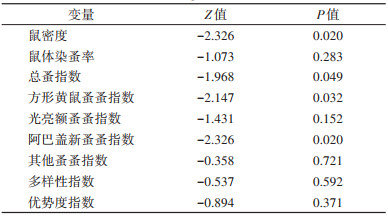
|
使用STL模型将达乌尔黄鼠密度及其体表寄生蚤感染情况的年月时间序列分解为随机误差时间序列、季节性时间序列与长期趋势时间序列,随机误差时间序列的波动形态与原始数据的形态相似。见图 2。

|
| 注:A达乌尔黄鼠密度时间分解图;B达乌尔黄鼠鼠体染蚤率时间分解图;C总蚤指数时间分解图;D方形黄鼠蚤蚤指数时间分解图;E光亮额蚤蚤指数时间分解图;F阿巴盖新蚤蚤指数时间分解图;G Shannon-Wiener多样性指数时间分解图;H Simpson优势度指数时间分解图;本图横坐标分隔节点以月为单位,且以每年的4-10月为1个周期,每年仅7个月。 图 2 2012-2021年内蒙古自治区锡林郭勒盟达乌尔黄鼠密度及其体表寄生蚤感染情况季节性分解模型时间分解图 Figure 2 Seasonal decomposition of time series of density and ectoparasitic flea infection of Spermophilus dauricus in Xilin Gol League, Inner Mongolia Autonomous Region, 2012-2021 |
| |
从长期趋势上看,鼠密度自2012-2020年呈现波动性下降趋势,而在2021年出现上升趋势,见图 2A。鼠体染蚤率在2020年后呈明显的下降趋势,见图 2B。总蚤指数(图 2C)与方形黄鼠蚤蚤指数(图 2D)的长期趋势较为一致,呈现波动下降趋势。光亮额蚤蚤指数的长期趋势波动较大,在2014-2016年其趋势变化出现了较为明显的波动,2021年呈现明显上升趋势, 见图 2E。阿巴盖新蚤蚤指数在2012年显著下降后趋势相对稳定,见图 2F。而多样性指数与优势度指数在2014年8月左右呈现明显的峰值,二者波动情况相似,见图 2G、2H。
从季节性趋势来看,锡林郭勒盟鼠疫自然疫源地鼠密度(图 2A)、鼠体染蚤率(图 2B)、总蚤指数(图 2C)、方形黄鼠蚤蚤指数(图 2D)、光亮额蚤蚤指数(图 2E)、阿巴盖新蚤蚤指数(图 2F)大多在7或8月呈现峰值,4-5月较低。多样性指数、优势度指数却相反,即4、5月呈现较高的峰值,而在7、8月呈现最低值,见图 2G、2H。
2.3 空间分布整理并分析锡林郭勒盟各鼠疫自然疫源地达乌尔黄鼠密度及其体蚤感染情况,可见布氏田鼠鼠疫自然疫源地平均鼠密度最低,仅0.24只/hm2,但该疫源地的鼠体染蚤率、总蚤指数、方形黄鼠蚤蚤指数及其他蚤指数在3个疫源地中最高。达乌尔黄鼠鼠疫自然疫源地的平均鼠密度最高,同时阿巴盖蚤指数、多样性指数与优势度指数在3个疫源地中最高。长爪沙鼠鼠疫自然疫源地中光亮额蚤蚤指数在3个疫源地中最高。见表 4。
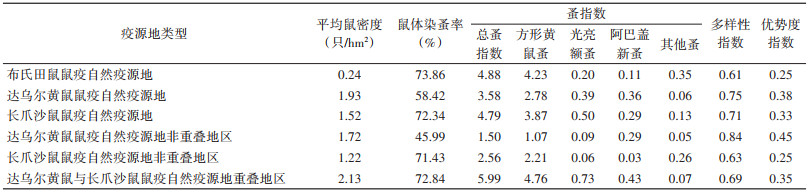
|
分析达乌尔黄鼠与长爪沙鼠鼠疫疫源地重叠地区与非重叠地区的达乌尔黄鼠及其染蚤情况,平均鼠密度、鼠体染蚤率、总蚤指数、方形黄鼠蚤蚤指数、光亮额蚤蚤指数、阿巴盖新蚤蚤指数在重叠地区最高。而重叠地区的其他蚤蚤指数低于长爪沙鼠鼠疫自然疫源地非重叠地区,多样性指数与优势度指数也均比达乌尔黄鼠鼠疫自然疫源地非重叠地区低。见表 4。
Kruskal-Wallis H秩和检验分析发现除其他蚤蚤指数外,达乌尔黄鼠密度及其体表寄生蚤感染情况在布氏田鼠鼠疫自然疫源地、达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠与非重叠地区的分布差异均具有统计学意义(均H > 35.00,均P < 0.001)。
长爪沙鼠鼠疫自然疫源地非重叠地区的达乌尔黄鼠密度(图3A)、总蚤指数(图3C)、方形黄鼠蚤蚤指数(图3D)与另外3种疫源地的分布差异均有统计学意义;光亮额蚤蚤指数(图3E)、阿巴盖新蚤蚤指数(图3F)、多样性指数(图3H)、优势度指数(图3I)与达乌尔黄鼠鼠疫自然疫源地非重叠地区、达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区的分布差异均有统计学意义;鼠体染蚤率(图3B)与达乌尔黄鼠鼠疫自然疫源地非重叠地区的分布差异有统计学意义。

|
| 注:A达乌尔黄鼠密度;B达乌尔黄鼠鼠体染蚤率;C总蚤指数;D方形黄鼠蚤蚤指数;E光亮额蚤蚤指数;F阿巴盖新蚤蚤指数;G其他蚤蚤指数;H Shannon-wiener多样性指数;I Simpson优势度指数;a代表调整后P < 0.05,b代表调整后P < 0.01,c代表调整后P < 0.001;疫源地指鼠疫自然疫源地。 图 3 内蒙古自治区锡林郭勒盟不同鼠疫自然疫源地达乌尔鼠密度与蚤感染多重比较结果 Figure 3 Multiple comparisons of density and flea infection of Spermophilus dauricus in different natural plague foci in Xilin Gol League, Inner Mongolia Autonomous Region |
| |
达乌尔黄鼠鼠疫自然疫源地非重叠地区与布氏田鼠鼠疫自然疫源地、达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区的鼠体染蚤率(图3B)、总蚤指数(图3C)、方形黄鼠蚤蚤指数(图3D)、阿巴盖新蚤蚤指数(图3F)、多样性指数(图3H)分布差异均有统计学意义;与布氏田鼠鼠疫自然疫源地的鼠密度(图3A)、优势度指数(图3I)分布差异均有统计学意义;与达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区的光亮额蚤蚤指数(图3E)分布差异有统计学意义。
达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区与布氏田鼠鼠疫自然疫源地的达乌尔黄鼠密度(图 3A)、总蚤指数(图 3C)、光亮额蚤蚤指数(图 3E)、阿巴盖新蚤蚤指数(图 3F)、多样性指数(图 3H)及优势度指数(图 3I)均有统计学差异。
2.4 时空分布Mann-Kendall时间趋势检验分析不同鼠疫自然疫源地的达乌尔黄鼠密度及其寄生蚤感染情况的年际趋势发现,布氏田鼠鼠疫自然疫源地达乌尔黄鼠平均密度10年间呈下降趋势,趋势有统计学意义(Z=-2.147,P=0.032);达乌尔黄鼠鼠疫自然疫源地非重叠地区达乌尔黄鼠平均密度、总蚤指数、方形黄鼠蚤蚤指数、阿巴盖新蚤蚤指数、其他蚤蚤指数、多样性指数与优势度指数均呈下降趋势(均Z < 0,P < 0.05);长爪沙鼠鼠疫自然疫源地非重叠地区的达乌尔黄鼠密度与其体表寄生蚤变化均无统计学意义(均P > 0.05);达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区的达乌尔黄鼠密度与鼠体染蚤率的下降趋势有统计学意义(均Z=-1.968,P=0.049)。
3 讨论达乌尔黄鼠是内蒙古锡林郭勒盟3种鼠疫自然疫源地的重要宿主动物,对鼠疫传播有重要作用。疫源地内鼠密度的变化对动物间鼠疫的流行具有重要影响[3, 12]。2012-2021年间锡林郭勒盟达乌尔黄鼠平均鼠密度为1.29只/hm2,同时盟内达乌尔黄鼠鼠疫自然疫源地监测点平均鼠密度为1.93只/hm2,虽仍略高于我国达乌尔黄鼠鼠疫自然疫源地最低鼠疫流行病学的密度控制标准(1只/hm2)[3],但相较于2001-2013年内蒙古察哈尔丘陵地区达乌尔黄鼠鼠疫自然疫源地平均鼠密度3.29只/hm2有所降低[3],说明近年锡林郭勒盟达乌尔黄鼠的灭鼠工作取得一定成效。同时STL模型分解的趋势分量发现鼠密度10年间整体变动呈下降趋势,但2021年却呈上升趋势,提示仍需加强对达乌尔黄鼠种群的监测与防控,预防其种群再次扩张。分解的季节周期分量发现鼠密度在7月呈现峰值,分析可能是由于其出蛰后4-6月繁衍[13-14],7月左右新生幼鼠几乎全部出来活动,加之该季节气候适宜,黄鼠活动频繁,因此在此时监测到的黄鼠种群密度最高,这一点与董维惠等[13]在内蒙古正镶白旗及费荣中等[14]在内蒙古赤峰调查结果一致。刘冠纯等[8]利用集中度法说明达乌尔黄鼠密度变化具有一定的季节性,但该研究发现河北省鼠疫自然疫源地康保牧场达乌尔黄鼠的密度高发峰值在4月,其次是7-9月,这与本研究结果存在差异,考虑可能是由于不同研究地区间温湿度差异影响黄鼠活动节律而致[15-16]。达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区平均鼠密度最高,同时年际下降趋势没有达乌尔黄鼠鼠疫自然疫源地非重叠地区明显。长爪沙鼠鼠疫自然疫源地非重叠地区平均鼠密度也较高,10年内也没有明显的下降趋势。白雪薇等[12]发现河北省长爪沙鼠鼠疫自然疫源地内达乌尔黄鼠与长爪沙鼠的种群存在演替现象,认为达乌尔黄鼠在河北省动物间鼠疫流行中起着协同作用,崇岩岷等[9]发现达乌尔黄鼠的鼠洞与长爪沙鼠洞穴之间没有严格的界限,常混合分布,均提示应加强对长爪沙鼠鼠疫自然疫源地特别是其与达乌尔黄鼠鼠疫自然疫源地重叠地区的达乌尔黄鼠种群监测及控制工作。

|
| 图 4 内蒙古自治区锡林郭勒盟不同鼠疫自然疫源地鼠密度与蚤感染年际趋势 Figure 4 Yearly trends of rodent density and flea infection in different natural plague foci in Xilin Gol League, Inner Mongolia Autonomous Region |
| |
蚤类可以传播多种疾病,也是传播鼠疫的重要媒介生物[11, 17]。蚤指数对鼠疫在动物间传播具有重要影响[18]。陈永明等[7]研究发现,达乌尔黄鼠体表寄生蚤的染蚤率在发现动物间疫情的年份高于平均染蚤率,可见染蚤率对鼠疫疫情也有一定的警示作用[19]。锡林郭勒盟10年鼠体染蚤率为61.51%,10年总蚤指数为3.43;其中达乌尔黄鼠鼠疫自然疫源地的10年鼠体染蚤率58.42%,10年总蚤指数为3.58,与2001-2013年达乌尔黄鼠鼠疫自然疫源地内的察哈尔丘陵地区[3]、2012-2021年赤峰市[20]、2011-2020年通辽市[21]、2013-2022年吉林省相比均较高[22]。尤其值得注意的是,锡林郭勒盟内布氏田鼠、长爪沙鼠鼠疫自然疫源地中的鼠体染蚤率均 > 70.00%,总蚤指数均 > 4.50,应该在加强锡林郭勒盟全盟灭蚤工作的同时着重对这2个疫源地的寄生蚤进行管控。本研究发现达乌尔黄鼠的鼠体染蚤率变化具有季节性规律,在8月和10月有2个峰值,这与陈永明等[7]的研究结果一致。同时,本研究发现,锡林郭勒盟这10年在鼠体总蚤指数下降的情况下,鼠体染蚤率并无明显下降。达乌尔黄鼠鼠疫的流行主要由黄鼠互相接触和交换寄生蚤来实现[23],提示即便寄生蚤数量减少,仍要关注可能由于鼠种群中感染寄生蚤的黄鼠数量的增多,导致鼠疫风险增加的情况。
媒介蚤类是中国人间鼠疫的主要感染途径,方形黄鼠蚤、光亮额蚤、阿巴盖新蚤等多种蚤均是可以自然染疫的媒介蚤类[24],而不同蚤种对鼠疫的保存与传播具有不同的效能[19],因此有必要加强对达乌尔黄鼠体表不同寄生蚤种的相关研究,为鼠疫防治提供针对性指导。本研究发现,方形黄鼠蚤是2012-2021年达乌尔黄鼠体表寄生的绝对优势蚤种,与内蒙古、吉林省、河北省等多地研究结果一致[7, 22, 25]。光亮额蚤和阿巴盖新蚤的数量也较多,是达乌尔黄鼠体表主要寄生蚤,与吴馨羽等[25]的研究结果相同。但与陈磊等[22]发现在吉林省除方形黄鼠蚤外,达乌尔黄鼠体表感染最多的寄生蚤为二齿新蚤的研究结果不同,考虑可能是不同地理位置的环境生态不同所致[25]。范蒙光等[26]发现达乌尔黄鼠鼠疫自然疫源地中阿巴盖新蚤与光亮额蚤在达乌尔黄鼠体表寄生蚤中的占比上升,这与本研究结果一致。本研究分析发现,在达乌尔黄鼠体表寄生蚤中,方形黄鼠蚤与阿巴盖新蚤的蚤指数降低有统计学意义,但光亮额蚤与其他蚤的蚤指数下降无统计学意义,提示光亮额蚤及其他蚤在达乌尔黄鼠体表寄生蚤群落的占比可能呈现一定程度的升高。总蚤指数与方形黄鼠蚤蚤指数的STL分解结果中长期趋势与季节趋势一致,较为明显地体现了总蚤指数受其绝对优势蚤的影响较大。白林庆等[27]认为在同一区域内,宿主动物的混居可以影响到疫源地内主要贮存宿主的寄生蚤种类,本研究发现主要寄生蚤蚤指数在达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地重叠地区大于布氏田鼠鼠疫自然疫源地和长爪沙鼠、达乌尔黄鼠2类鼠疫疫源地的非重叠地区,考虑可能是由于重叠地区内达乌尔黄鼠与长爪沙鼠体蚤频繁交换造成的。
鼠类体表寄生蚤群落多样性的变化对鼠疫的维持与传播具有重要意义[11],加强对寄生蚤群落多样性的研究有助于了解寄生蚤群落特征,为防范及预测鼠疫等蚤传疾病溢出提供建议[25]。吴馨羽等[25]的研究发现达乌尔黄鼠体表寄生蚤群落多样性在春季较高,这与本研究结果一致。本研究发现达乌尔黄鼠体表寄生蚤群落多样性在时空上均表现出蚤指数较低时,多样性指数与优势度指数较高的现象。在总蚤指数与主要寄生蚤种蚤指数较低的季节,达乌尔黄鼠体表寄生蚤群落多样性指数较高,同时在总蚤指数与各主要寄生蚤种蚤指数较低的达乌尔黄鼠鼠疫自然疫源地,其体表寄生蚤群落多样性指数较高,推测多样性指数的变化一定程度上会受到蚤指数的影响。Shannon-Wiener多样性指数对群落中的稀有种较为敏感,Simpson优势度指数对群落中的优势种和普通种较为敏感[28],本研究发现这2个指标在时空上的变化几乎一致,可见达乌尔黄鼠体表寄生蚤的群落结构变化是较为稳定的,或许是由于不同蚤种对不同季节不同环境中的适应性不同所致,对于多样性季节性变化的具体原因值得进一步探究。群落研究在鼠疫媒介昆虫生态学研究中较为薄弱[29]。本研究发现不同鼠疫自然疫源地的群落多样性指数差异具有统计学意义,可能是由于宿主动物分布地区和生境内宿主动物群落结构不同导致寄生蚤群落结构存在差异[25]。陈永明等[30]分析发现,在长爪沙鼠体表寄生蚤优势度较高的时候更有利于动物间鼠疫的发生。提示我们后续应该加强对达乌尔黄鼠体表寄生蚤群落多样性的相关研究,以便为鼠疫及相关蚤传疾病提出更有针对性的应对措施。
综上所述,加强对锡林郭勒盟地区的达乌尔黄鼠及其体表寄生蚤感染的防治工作不容忽视,根据各监测点的具体情况因地制宜加强防鼠灭蚤工作,要着重注意达乌尔黄鼠与长爪沙鼠鼠疫自然疫源地特别是各疫源地重叠地区的鼠类种群变化,重点关注布氏田鼠及长爪沙鼠鼠疫自然疫源地的蚤类寄生情况。在黄鼠及其体表寄生蚤数量到达峰值前,黄鼠出蛰后的4-6月繁殖期[31],针对所在区域内的所有鼠类种群进行灭鼠灭蚤工作,更有利于控制黄鼠及其体表寄生蚤种群数量,严防鼠疫发生和流行。
利益冲突 无
| [1] |
徐爱玲, 栗冬梅, 刘起勇. "鼠源疾病"的概念与内涵的系统评价[J]. 中国媒介生物学及控制杂志, 2022, 33(2): 161-170. Xu AL, Li DM, Liu QY. The concept and connotation of "rodent-borne diseases": A systematic study[J]. Chin J Vector Biol Control, 2022, 33(2): 161-170. DOI:10.11853/j.issn.1003.8280.2022.02.001 |
| [2] |
Reis Da Silva Fernandes DL, Filgueira Bezerra M, Sobreira Bezerra Da Silva M, et al. Rodent hosts and flea vectors in Brazilian plague foci: A review[J]. Integr Zool, 2021, 16(6): 810-819. DOI:10.1111/1749-4877.12480 |
| [3] |
韩冰, 赵钢, 张忠兵, 等. 内蒙古地区2001-2013年达乌尔黄鼠鼠疫疫源地疫情分析[J]. 中国媒介生物学及控制杂志, 2016, 27(2): 190-193. Han B, Zhao G, Zhang ZB, et al. The analysis of Spermophilus dauricus plague natural foci in Inner Mongolia during 2001-2013[J]. Chin J Vector Biol Control, 2016, 27(2): 190-193. DOI:10.11853/j.issn.1003.8280.2016.02.026 |
| [4] |
李仲来, 周方孝, 李书宝, 等. 达乌尔黄鼠鼠疫预报的数学模型(Ⅷ)[J]. 中国地方病防治杂志, 2003, 18(3): 129-130. Li ZL, Zhou FX, Li SB, et al. Mathematical models for forecast of epizootic plague of Spermophilus dauricus (Ⅷ)[J]. Chin J Ctrl Endem Dis, 2003, 18(3): 129-130. DOI:10.3969/j.issn.1001-1889.2003.03.001 |
| [5] |
张璐, 韦晓慧, 常楠, 等. 内蒙古锡林郭勒盟2012-2021年鼠类寄生蚤构成和蚤指数时空分布特征分析[J]. 中国媒介生物学及控制杂志, 2023, 34(1): 82-90. Zhang L, Wei XH, Chang N, et al. Composition of parasitic fleas on rodents and spatiotemporal distribution of flea index in Xilin Gol league, Inner Mongolia Autonomous Region, China, 2012-2021[J]. Chin J Vector Biol Control, 2023, 34(1): 82-90. DOI:10.11853/j.issn.1003.8280.2023.01.015 |
| [6] |
张春华, 吕景生, 浦清江. 达乌尔黄鼠疫源地的现状及防治对策[J]. 中国地方病防治杂志, 2004, 19(6): 345-348. Zhang CH, Lyu JS, Pu QJ. The present situation and control countermeasure of Spermophilus dauricus plague natural foci[J]. Chin J Ctrl Endem Dis, 2004, 19(6): 345-348. DOI:10.3969/j.issn.1001-1889.2004.06.010 |
| [7] |
陈永明, 康东梅, 闫东, 等. 1981-2020年河北省鼠疫自然疫源地康保牧场达乌尔黄鼠寄生蚤调查分析[J]. 中华卫生杀虫药械, 2022, 28(3): 226-229. Chen YM, Kang DM, Yan D, et al. Investigation and analysis on parasitic fleas of Spermophilus dauricus in Kangbao pasture of plague natural focus in Hebei Province from 1981 to 2020[J]. Chin J Hyg Insectic Equip, 2022, 28(3): 226-229. DOI:10.19821/j.1671-2781.2022.03.010 |
| [8] |
刘冠纯, 康东梅, 闫东, 等. 应用集中度法探讨鼠疫宿主动物达乌尔黄鼠密度变化规律[J]. 中国媒介生物学及控制杂志, 2021, 32(2): 158-160. Liu GC, Kang DM, Yan D, et al. A study of the density of plague host Spermophilus dauricus by concentration method[J]. Chin J Vector Biol Control, 2021, 32(2): 158-160. DOI:10.11853/j.issn.1003.8280.2021.02.007 |
| [9] |
崇岩岷, 徐崇洋岷, 史献明, 等. 河北省鼠疫自然疫源地2007-2016年达乌尔黄鼠数量和分布变化的研究[J]. 中国媒介生物学及控制杂志, 2018, 29(6): 639-640. Chong YM, Xu CYM, Shi XM, et al. Study on the change of numbers and distributions of Spermophilus dauricus in plague natural foci of Hebei Province in 2007-2016[J]. Chin J Vector Biol Control, 2018, 29(6): 639-640. DOI:10.11853/j.issn.1003.8280.2018.06.023 |
| [10] |
李轲, 王振旭, 张璐, 等. 内蒙古自治区锡林郭勒盟2012-2021年气象因素和鼠体蚤指数对长爪沙鼠种群密度的影响[J]. 中国媒介生物学及控制杂志, 2024, 35(2): 212-217. Li K, Wang ZX, Zhang L, et al. The impact of meteorological factors and flea index on the density of Meriones unguiculatus in Xilingol League, Inner Mongolia Autonomous Region, China, 2012-2021[J]. Chin J Vector Biol Control, 2024, 35(2): 212-217. DOI:10.11853/j.issn.1003.8280.2024.02.015 |
| [11] |
Eads DA, Biggins DE, Gage KL. Ecology and management of plague in diverse communities of rodents and fleas[J]. Vector Borne Zoonotic Dis, 2020, 20(12): 888-896. DOI:10.1089/vbz.2020.2625 |
| [12] |
白雪薇, 牛艳芬, 陈永明, 等. 2001-2020年河北省鼠疫自然疫源地长爪沙鼠与达乌尔黄鼠密度变化趋势研究[J]. 中华卫生杀虫药械, 2021, 27(3): 226-230. Bai XW, Niu YF, Chen YM, et al. Study on the density change trend of Meriones unguiculatus and Spermophilus dauricus in the plague natural foci in Hebei Province from 2001 to 2020[J]. Chin J Hyg Insectic Equip, 2021, 27(3): 226-230. DOI:10.19821/j.1671-2781.2021.03.010 |
| [13] |
董维惠, 侯希贤, 周延林, 等. 达乌尔黄鼠种群繁殖与数量动态研究[J]. 卫生杀虫药械, 1998, 4(3): 4-7. Dong WH, Hou XX, Zhou YL, et al. A study of the reproduction and population dynamics of Spermophilus Dauricus[J]. Pestic Equip Pest Control, 1998, 4(3): 4-7. DOI:10.19821/j.1671-2781.1998.03.002 |
| [14] |
费荣中, 李景原, 商志宽, 等. 达乌尔黄鼠的生态研究[J]. 动物学报, 1975, 21(1): 18-29. Fei RZ, Li JY, Shang ZK, et al. Ecological research on Spermophilus dauricus[J]. Curr Zool, 1975, 21(1): 18-29. |
| [15] |
王静, 纪维红, 苏军虎, 等. 达乌尔黄鼠(Spermophilus dauricus)生态学研究进展[J]. 中国农学通报, 2015, 31(8): 33-39. Wang J, Ji WH, Su JH, et al. The research status on Spermophilus dauricus[J]. Chin Agric Sci Bull, 2015, 31(8): 33-39. |
| [16] |
张贵, 孙启廷, 刘振才, 等. 吉林省鼠疫自然疫源地达乌尔黄鼠活动规律的研究[J]. 中国地方病防治杂志, 2009, 24(4): 273-274. Zhang G, Sun QT, Liu ZC, et al. A study on law of activity of S. dauricus in Jilin Province plague nature focus[J]. Chin J Ctrl Endem Dis, 2009, 24(4): 273-274. |
| [17] |
Bitam I, Dittmar K, Parola P, et al. Fleas and flea-borne diseases[J]. Int J Infect Dis, 2010, 14(8): e667-e676. DOI:10.1016/j.ijid.2009.11.011 |
| [18] |
刘振才, 周晓磊, 张博宇, 等. 动物鼠疫预测模型及预警指标的建立[J]. 中国地方病防治杂志, 2015, 30(1): 1-3. Liu ZC, Zhou XL, Zhang BY, et al. Establishment of predictive model and warning index of animal plague[J]. Chin J Ctrl Endem Dis, 2015, 30(1): 1-3. |
| [19] |
蔡文凤, 罗剑龙, 苏丽琼, 等. 云南省剑川县鼠疫疫源地小型兽类体蚤构成及群落多样性特征[J]. 中国媒介生物学及控制杂志, 2019, 30(5): 557-560. Cai WF, Luo JL, Su LQ, et al. Composition and community diversity of small mammals fleas in the plague foci in Jianchuan County, Yunnan Province, China[J]. Chin J Vector Biol Control, 2019, 30(5): 557-560. DOI:10.11853/j.issn.1003.8280.2019.05.018 |
| [20] |
张雅娴, 刘艳华, 郭东. 赤峰市达乌尔黄鼠动物鼠疫监测流行病学分析[J]. 中国地方病防治, 2022, 37(1): 66-67. Zhang YX, Liu YH, Guo D. Epidemiological analysis of the surveillance of the animal plague of Spermophilus dauricus in Chifeng City, China[J]. Chin J Ctrl Endem Dis, 2022, 37(1): 66-67. |
| [21] |
马丽, 马跃, 张磊, 等. 通辽市2011-2020年动物间鼠疫疫情监测结果分析[J]. 医学动物防制, 2022, 38(8): 801-803. Ma L, Ma Y, Zhang L, et al. Analysis on surveillance results of animal plague in Tongliao from 2011 to 2020[J]. J Med Pest Control, 2022, 38(8): 801-803. DOI:10.7629/yxdwfz202208022 |
| [22] |
陈磊, 张贵军, 阚朋甲. 2013-2022年吉林省鼠疫自然疫源地蚤类种群结构特征分析[J]. 中国地方病防治, 2023, 38(5): 399-400. Chen L, Zhang GJ, Kan PJ. Analysis of the species composition characteristics of fleas in the natural focus of plague in Jilin Province from 2013 to 2022[J]. Chin J Ctrl Endem Dis, 2023, 38(5): 399-400. |
| [23] |
赵思阳, 石杲. 内蒙古西辽河流域黄鼠寄生蚤与鼠疫动物病关系的研究[J]. 医学动物防制, 2002, 18(4): 180-181. Zhao SY, Shi G. Research on the relationship between fleas of the Spermophilus dauricus and animal plague in the West Liaohe River basin of Inner Mongolia[J]. J Med Pest Control, 2002, 18(4): 180-181. |
| [24] |
刘成赋, 石杲. 中国达乌尔黄鼠鼠疫媒介控制的研究[J]. 中国媒介生物学及控制杂志, 2007, 18(4): 320-321. Liu CF, Shi G. Research on plague vector control of Chinese Spermohilus dauricus[J]. Chin J Vector Biol Control, 2007, 18(4): 320-321. DOI:10.3969/j.issn.1003-4692.2007.04.019 |
| [25] |
吴馨羽, 明明, 袁帅, 等. 典型草原两种优势鼠种体表寄生蚤群落的影响因素[J]. 中国草地学报, 2024, 46(9): 96-104. Wu XY, Ming M, Yuan S, et al. Effects of factors on flea community dynamics on the body surface of two dominant rodent species in typical steppe ecosystem[J]. Chin J Grassl, 2024, 46(9): 96-104. DOI:10.16742/j.zgcdxb.20230311 |
| [26] |
范蒙光, 李建云, 常子丽, 等. 内蒙古达乌尔黄鼠疫源地主要宿主寄生蚤调查结果分析[J]. 医学动物防制, 2015, 31(9): 945-948. Fan MG, Li JY, Chang ZL, et al. Spermophilus dauricus epidemic focus main hosts parasitical fleas investigation results analysis of Inner Mongolia[J]. J Med Pest Control, 2015, 31(9): 945-948. |
| [27] |
白林庆, 司晓艳, 涛波. 内蒙古主要鼠疫传播媒介蚤类的分布特征及其流行病学意义[J]. 实用预防医学, 2015, 22(5): 639-641. Bai LQ, Si XY, Tao B. Distribution characteristics and epidemiological significance of fleas, the main transmitting vectors of the plague in Inner Mongolia[J]. Pract Prev Med, 2015, 22(5): 639-641. DOI:10.3969/j.issn.1006-3110.2015.05.044 |
| [28] |
王利清, 杨玉平, 董维惠, 等. 典型草原鼠类群落结构研究[J]. 中国草地学报, 2012, 34(3): 75-81. Wang LQ, Yang YP, Dong WH, et al. Study on rodent community structure in typical grassland[J]. Chin J Grassl, 2012, 34(3): 75-81. DOI:10.3969/j.issn.1673-5021.2012.03.013 |
| [29] |
李晔, 柳付明, 潘孝猛, 等. 浙西南地区鼠疫宿主动物体表寄生虫群落多样性和病原体携带现况调查[J]. 中国地方病防治杂志, 2021, 36(3): 207-211. Li Y, Liu FM, Pan XM, et al. Investigation on community diversity and pathogen carrying status of plague host animals in the southwest region of Zhejiang Province[J]. Chin J Ctrl Endem Dis, 2021, 36(3): 207-211. |
| [30] |
陈永明, 闫东, 刘冠纯, 等. 河北省鼠疫自然疫源地长爪沙鼠体蚤多样性研究[J]. 中华卫生杀虫药械, 2018, 24(3): 255-257, 260. Chen YM, Yan D, Liu GC, et al. A diversity of body fleas on Meriones unguiculatus in plague natural epidemic foci of Hebei Province[J]. Chin J Hyg Insectic Equip, 2018, 24(3): 255-257, 260. DOI:10.19821/j.1671-2781.2018.03.011 |
| [31] |
董维惠, 侯希贤, 杨玉平. 我国草原常见害鼠药物防治适宜时机的选择[J]. 中国草地学报, 2008, 30(4): 107-112. Dong WH, Hou XX, Yang YP. The choice of appropriate opportunity of controlling common rodents with rodenticides in grassland of China[J]. Chin J Grassl, 2008, 30(4): 107-112. |