 2012, Vol. 28
2012, Vol. 28

应激泛指机体受到强烈或有害刺激后产生的一系列生理病理反应。适度应激刺激有助于学习记忆功能,长期或过强应激则损害学习记忆功能。在脑内,海马和前额叶是最易受应激刺激损伤脑区,也是与学习记忆功能关系密切的重要区域[1]。研究表明,慢性应激可引起动物脑海马神经元形态改变和大量丢失,动物学习记忆能力受到明显影响[2, 3]。神经细胞膜流动性改变将影响各种膜功能,包括离子通透性、跨膜物质转运、胞内信号转导等;脑的许多疾病发生发展过程中,膜流动性也显示相应的改变。本研究采用多因素慢性应激动物模型,检测不同月龄小鼠Morris水迷宫空间学习记忆能力,测定小鼠前脑皮层和海马突触体膜流动性、突触体内游离Ca2+浓度变化,探讨慢性应激对小鼠学习记忆功能影响及其机制。
1 材料与方法 1.1 实验动物与分组健康昆明小白鼠3月龄(青年)、14月龄(老年),雌雄各半,中国医学科学院医学生物学研究所提供(许可证号:云卫动管第B05号)。将青年和老年小鼠随机分为对照组和应激组。应激组小鼠共接受21d不同慢性应激源。采用5种应激方式:足底电击(电压33V,每隔1min刺激1次,持续10s,共15次)、束缚(1h/d)、束缚+热刺激(1h/d,30℃)、冷水刺激(6℃冷水游泳3min)、高台(1h/d,高1.6m)。每天随机选择1种应激方式。对照组动物不接受应激源,以同样条件饲养。应激后,对小鼠进行各指标检测。
1.2 Morris水迷宫学习记忆能力检测定位航行试验历时2d,每只小鼠每天接受8次找平台训练,分2个训练周期(上午和下午),每个训练周期4次,每次训练2min。记录小鼠寻找到隐藏在水下平台时间即逃避潜伏期;空间搜索试验在第3d进行,撤除平台,从第三象限将小鼠放入水中,记录120s内小鼠在4个象限游泳(停留)时间,检测小鼠对原平台(目标象限)记忆保持能力。
1.3 小鼠前脑皮层和海马突触体制备将小鼠断头处死,迅速分离两侧前脑皮层和海马并称重,加入10倍体积0.32mol/L蔗糖溶液(内含10mmol/L Hepes,pH7.4,W:V=1:10)进行匀浆。所有操作均在0~4℃进行,匀浆液按差速蔗糖梯度离心法分离制备突触体,最后用人工脑脊液缓慢悬浮,得到突触体悬液。用Lowry法测定悬液蛋白浓度。突触体悬液保存在0~4℃,在分离后2~3h内完成测定。
1.4 突触体膜流动性和突触体内游离Ca2+浓度测定取前脑皮层和海马突触体悬液,用人工脑脊液调整蛋白浓度为0.3mg/mL,与等体积荧光探针1,6-二苯基-1,3,5-己三烯(1,6-diphenyl-1,3,5-hexatriene,DPH,Fluka公司,终浓度2×10-6mol/L)稀释液,37℃孵育30min。用LS50B荧光分光光度计(美国Perkin-Elmer公司)检测突触体膜荧光偏振度(P),用膜各向异性(r)值表示突触体膜流动性大小,r值越小,膜流动性越大。取前脑皮层和海马突触体悬液,用ACSF调整蛋白浓度为1.5~2.5mg/mL,加入Fura2-AM(美国Sigma公司,终浓度:5μmol/L),37℃孵育30min,于11000×g离心10min,取沉淀,用人工脑脊液悬浮,于0~4℃保存,采用LS 50B型荧光分光光度计,在37℃条件下,测定其荧光强度及荧光比值。计算突触体内钙离子浓度。
1.5 统计分析数据x ± s表示,采用SPSS 13.0软件进行统计处理,逃避潜伏期采用重复测量方差分析,各象限游泳时间采用单因素方差分析,组间比较采用LSD-t检验或非配对t检验。以P<0.05表示差异有统计学意义。
2 结果 2.1 不同月龄小鼠定位航行学习记忆能力变化(表 1)| 表 1 慢性应激后各组小鼠逃避潜伏期比较(s,x ± s,n=10) |

青年应激组小鼠第2、3、4训练周期逃避潜伏期比青年对照组明显延长(P<0.01);老年应激组小鼠在第3、4训练周期逃避潜伏期均明显长于老年对照组(P<0.01)。与青年对照组比较,老年对照组在第1、2训练周期逃避潜伏期均明显延长(t=2.382,P<0.05;t=2.786,P<0.01)。
2.2 不同月龄小鼠空间搜索能力变化(表 2)| 表 2 慢性应激后各组小鼠在各象限停留时间(s,x ± s,n=10) |
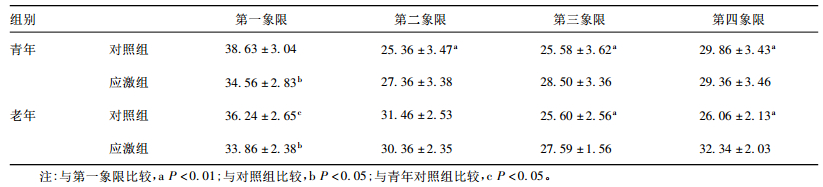
青年对照组和老年对照组小鼠在目标象限停留时间明显长于在其他3个象限时间(P<0.01);与对照组比较,青年和老年应激组小鼠在目标象限停留时间均明显缩短(t=2.532、2.326,P<0.05)。与青年对照组比较,老年对照组在目标象限停留时间明显缩短(t=2.238,P<0.05)。
2.3 突触体膜流动性和突触体内游离Ca2+浓度改变(表 3)| 表 3 慢性应激后各组小鼠突触体膜流动性和游离Ca2+浓度变化(x ± s,n=10) |
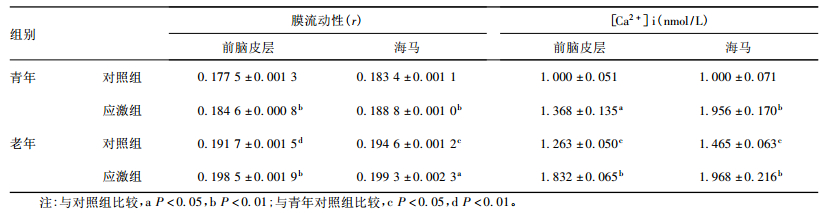
与青年对照组比较,老年对照组小鼠前脑皮层和海马突触体膜r值均明显增加(t=3.263,P<0.05;t=5.536,P<0.01),膜流动性明显降低。青年和老年应激组小鼠前脑皮层和海马突触体膜流动性明显均低于对照组小鼠(青年t=6.436、4.068,P<0.01;老年t=5.378、3.232,P<0.05)。与青年对照组比较,老年对照组小鼠前脑皮层和海马突触体内游离Ca2+浓度均明显增加(t=2.632、3.086,P<0.05);与对照组比较,青年和老年应激组小鼠前脑皮层和海马突触体内游离Ca2+浓度均明显升高(t=2.883~5.162,P<0.05)。
3 讨论多因素慢性应激,可引起动物脑内神经递质、神经激素和下丘脑-垂体-肾上腺轴改变,导致动物行为和学习记忆功能损害[4]。本研究采用多因素慢性应激模型,结果表明,多因素慢性应激引起小鼠Morris水迷宫空间学习记忆能力严重损伤;小鼠前脑皮层和海马突触体膜流动性明显降低,突触体内游离Ca2+浓度明显升高。研究报道,慢性应激后应激激素水平增高,可使细胞内Ca2+明显增加,并由此激活超氧化自由基产生过程,引起脑组织脂质过氧化反应,导致海马依赖空间学习记忆能力明显降低[5]。研究表明,在衰老中脑海马的脂质过氧化活动明显增强,动物学习记忆能力明显降低[6]。已经明确,慢性应激引起动物空间学习记忆功能受损,与前脑皮层和海马突触体膜流动性明显降低,突触体内游离Ca2+浓度明显增加有关[7]。
慢性应激可以通过改变海马突触可塑性,对海马介导的记忆有持久性损伤[8];在脑内,应激因素除对海马有影响外,对前额叶、杏仁核等也有作用,明显损害大鼠、猴的依赖前额叶的工作记忆能力[3, 9]。本研究结果表明,慢性应激可明显损害小鼠空间学习记忆能力,同时小鼠前脑皮层和海马突触体内游离Ca2+浓度显著升高,突触体膜流动性明显降低。提示,突触体膜流动性和突触体内游离Ca2+浓度在衰老和慢性应激引起的空间学习记忆能力损伤中可能发挥重要作用。
| [1] | McEwen BS.Protective and damaging effects of stress mediators:central role of the brain[J].Dialo Clini Neurosci,2006,8(4):367-381. |
| [2] | Dranovsky A,Hen R.Hippocampal neurogenesis:regulation by stress and antidepressants[J].Biol Psychiatry,2006,59(12):1136-1143. |
| [3] | Arnsten AF.Stress signalling pathways that impair prefrontal cortex structure and function[J].Nature,2009,10(6):410-422. |
| [4] | Lupien SJ,McEwen BS,Gunnar MR,et al.Effects of stress throughout the lifespan on the brain,behaviour and cognition[J]. Nat Rev Neurosci,2009,10(6):434-445. |
| [5] | Bruel-Jungerman E,Rampon C,Laroche S.Adult hippocampal neurogenesis,synaptic plasticity and memory:facts and hypotheses[J].Rev Neurosci,2007,18(2):93-114. |
| [6] | 李亚,王建军.β-AP对衰老模型大鼠脑海马组织中脂质过氧化的影响[J].中国公共卫生,2002,18(2):306. |
| [7] | 李亚,孔宏,宋倩,等.慢性应激对大鼠空间学习记忆及海马和前脑皮层突触体膜流动性的影响[J].心理学报,2010,42(2):235-240. |
| [8] | Hill MN,Hunter RG,McEwen BS.Chronic stress differentially regulates cannabinoid CB1 receptor binding in distinct hippocampal subfields[J].Eur J Pharmacol,2009,614(1-3):66-69. |
| [9] | Roozendaal B,McEwen BS,Chattarji S.Stress,memory and the amygdala[J].Nat Rev Neurosci,2009,10(6):423-433. |