 2011, Vol. 27
2011, Vol. 27



2. 新疆食管癌研究所;
3. 新疆医科大学附属肿瘤医院;
4. 新疆医科大学第一附属医院;
5. 新疆伊犁州友谊医院
人类白细胞抗原( hum an leuko cy te antigen,HLA )位于第 6号染色体,参与免疫识别和应答及抗原处理,是机体抵御异已的第一防线,是人类基因组中最复杂、最具多态性的遗传体系。研究显示,其中非经典HLA-Ñ 类抗原分子HLA-G 基因的HLA-G 14bp 缺失( rs16375) 多态性〔1, 2, 3, 4, 5〕、HLA-G* 0105N ( rs41557518) 的多态性〔6, 7〕分别与一些免疫性疾病有相关性。C astelli等〔8〕曾研究rs16375位点多态性与巴西人群膀胱癌易感性关系,但未发现14bp缺失多态性与膀胱癌易感性有关。为了探讨这2个位点与新疆哈萨克族食管癌的关系,从免疫学角度探讨食管癌发病机制,为临床诊断和治疗提供理论依据,本课题收集2005- 2007 年新疆部分新发食管癌病例,采用聚合酶链式反应- 限制性片段长度多态( PCRRFLP) 技术,检测分析上述2个位点多态性在食管癌发病中的作用。
1 对象与方法 1.1 对象收集2005- 2007 年新疆自治区人民医院、新疆医科大学第一附属医院、第二附属医院、附属肿瘤医院、伊犁州新华医院、伊犁州友谊医院等6所医院经内窥镜、X线及病理确诊的哈萨克族原发性食管癌患者132 例作为病例组; 选择同期住院与病例无血缘关系、同民族、同性别、年龄±5岁,非肿瘤、非消化道疾病、非自身免疫性疾病患者254例,作为对照组。
1.2 方法 1.2.1 基因组DNA提取在征得患者知情同意情况下,采集2组病例的外周静脉血2 mL,乙二胺四乙酸( e thy lene diamine tetraacetic ac id,EDTA )抗凝,- 20 ℃ 保存,采用改良M iller 盐析法〔10〕提取DNA,溶于三羟甲基氨基甲烷- 乙二胺乙酸二钠( TE)缓冲液( 10 mmo l/L,pH 810 Tris-HC,l 1 mmo l/L EDTA) 中,- 20 ℃ 保存备用。
1.2. 2 聚合酶链反应一限制性片段长度多态性( PCRRFLP) 分析( 1) 引物: 采用Pr im er 3. 0软件设计特异性引物,HLA-G 14bp缺失( rs16375 ) 引物序列: 上游引物5 G' TGATGGGCTGTTTAAAGTGTCACC 3 ,' 下游引物5 'GGAAGGAATGCAGTTCAGCATGA 3,'合成片段长度为224 bp。HLAG * 0105N( rs41557518) 引物序列: 上游引物5-'CCAGTGGATGATTGGCTGCG- 3,'下游引物5-'CCGTTCTCCAGGTATCTGTG-3 ,'合成片段长度为240 bp。引物由上海生物工程公司合成。 ( 2) 2个位点的扩增体系: 20 μL,其中DNA ( 50 ng /LL)模版 1μL,加入2 × PCR TaqM ix 10 μL,双向引物( 10 μm o l/L)各1 μL,加双蒸水至20 μL。( 3) 反应条件: 95 ℃ 预变性4 m in,95 ℃ 30 s、61 ℃ 退火30s、72 ℃ 30 s进行35 个循环,最后 72 ℃ 延伸10 m in。其中,HLA-G* 0105N ( rs41557518) 扩增后产物采用内切酶10U /E co0109 I( 识别序列为rg /gnccy) 进行37 ℃ 酶切过夜。( 4 ) 结果鉴定: H LA-G 14bp 缺失 ( rs16375) : 取PCR 产物10 μL,于4% 琼脂糖凝胶( 加有 21 5LL 核酸染料) 100 V 电泳40 m in后,凝胶成像仪检测,选缺失纯合子和插入纯合子样品的PCR原液送上海生物工程公司测序。H LA-G* 0105N: 取酶切产物10 μL,于21 5% 琼脂糖凝胶( 加有21 5LL核酸染料) 100V 电泳40 m in后,凝胶成像仪检测。
1.3 统计分析数据录入M icroso ft Ex cel 2003建立数据库,采用SPSS 15.0软件进行统计分析。遗传平衡定律( H ardy- W e inbe rg)检验及基因多态性位点基因型及等位基因频率比较采用χ2 检验,检验水准α= 0.05。
2 结果 2.1 一般情况收集哈萨克族食管癌患者132 例,其中男性 80例,女性52 例,平均年龄( 57. 92 ± 8.40) 岁; 对照组254 例,其中男性157例,女性97 例,平均年龄( 60. 01 ± 10.47) 岁。经检验,病例组与对照组在性别、年龄、婚姻状况、文化程度等方面的均衡性良好。
2. 2 HLA-G基因2个位点PCR 扩增图 2.2.1 HLA-G 基因14bp 缺失( rs16375) 位点基因型(图 1) | 注: M: LowMW DNA M arker-A; 1: - 14 bp /- 14 bp 基因型( 210 bp) ; 2: - 14 bp /+ 14 bp基因型(分别为210和224 bp ) ; 3、4: + 14 bp /+ 14 基因型( 224 bp)。 图 1 HLA-G基因14bp del( rs16375 )位点PCR扩增图 |
rs16375位点有3种基因型: 分别为- 14 bp /- 14 bp,长度为210 bp; - 14 bp /+ 14 bp,长度分别为210 和224 bp; + 14 bp / + 14 bp,长度为224 bp。测序结果与GeneBank B last 中HLA-G基因序列比对,判型一致,224 bp产物比210 bp产物多出1个14 bp的基因序列: AGGCATGAACAAAT。
2.2.2 H LA-G 基因* 0105N( rs41557518) 位点基因型(图 2) | 注: M: 50bp Ladder DNA M arker; 1、2、3、4、5、7: C /C 基因型( 136、 104 bp) ; 6、8: C /- 基因型( 240、136、104 bp )。 图 2 HLA-G* 0105N ( rs41557518)基因酶切产物琼脂糖凝胶电泳图 |
rs41557518 位点扩增产物片段长度为240 bp,包含 E co0109I 酶切片段。C /C 基因型的扩增产物被完全切开为 136、104 bp 2 条片段; C /- 基因型为杂合子,可见240、136、 104 bp 3条片段,未见- /- 纯合子基因型; 测序结果与酶切分型结果一致。
2.3 不同组别HLA-G基因2个位点基因型及等位基因(表 1)| 表 1 不同组别HLA-G基因不同基因型及等位基因分布 |
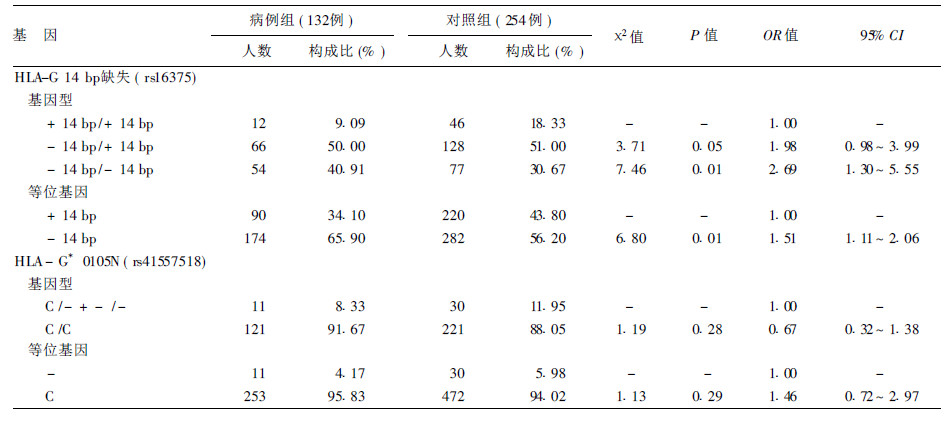
2组H LA-G 14bp 缺失( rs16375) 位点的基因型频率分布差异、等位基因频率差异均有统计学意义,携带- 14 bp /- 14 bp 基因型的个体发生食管癌的危险性是+ 14bp / + 14bp基因型的21 69倍; 2组HLA-G* 0105N( rs41557518)位点的基因型频率分布差异等位基因频率差异均无统计学意义。
2.4 H ardy-W e inberg遗传平衡检验检验结果显示,对照组 HLA-G基因rs16375、rs41557518位点基因型频率实际值与理论值比较,差异无统计学意义( P > 0.05) ,表明在哈萨克族人群中HLA-G基因型频率均已达到遗传平衡,具有人群代表性。
2.5 2 个位点多态性联合作用(表 2)| 表 2 哈萨克族食管癌与HLA-G基因14bp缺失与HLA-G* 0105N位点基因联合作用的关系 |
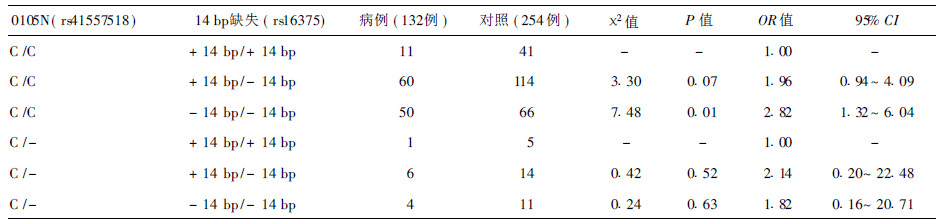
与同时携带HLA-G 14bp( + 14bp /+ 14bp) 和HLA-G* 0105N ( C /C) 的个体比较,同时携带HLA-G 14bp ( - 14bp / - 14bp) 和HLA-G* 0105N ( C /C)的个体患食管癌风险增加,提示HLA-G 基因14bp 缺失与HLA-G 基因* 0105N 在食管癌发生中存在联合作用。
3 讨论本研究结果显示,哈萨克族HLA-G 基因14bp 缺失 ( rs16375) 等位基因+ 14bp、- 14bp 分布频率为43180% 、 56.20%,与美国国家生物技术信息中心( National Center for B io techno logy Inform ation,NCB I) 多态数据库中此位点( db- SNP) ( http: / /www. ncb.i nlm. nih. gov /snp)欧洲人群分布频率 6810%、3210% 比较,差异无统计学差异( χ2 = 3.06,P = 0. 08); L in〔11〕等人研究欧洲人群HLA-G* 0105N 位点等位基因分布频率为97.50%、2.50%。本研究中,哈萨克族HLAG * 0105N等位基因C、- 分布频率为94. 20%、6. 0%,差异亦无统计学差异( χ2 = 1.08,P = 0.03),表明本研究所获得的样本与NCB I提供的数据一致性较好。
近来研究表明,H LA-G 在多种肿瘤细胞中表达,而且被意外地发现于多种恶性细胞就像HLA-G 在滋养细胞逃避母体异体识别过程中发挥的作用,它在肿瘤细胞中的异常表达同样被认为可能是肿瘤细胞得以逃避宿主免疫监视的手段〔12〕。HLA-G在转录后其相应的蛋白质产物有: 4种膜结合型( mH LA-G,即G1、G2、G3、G4 ) 和3 种可溶型HLA-G ( sH LA-G,即G5、G6、G7) 蛋白质分子。多数学者认为1132,sHLA-G既可以同mH LA-G 一起在肿瘤的原发位置发挥作用,也能通过血液循环在远处发挥作用,因此,血清sHLA-G 的浓集会降低机体的免疫监督功能,间接促进了肿瘤的发生发展。
HLA-G 14bp缺失多态性是指存在于HLA-G 基因外显子8 中3c非翻译区的一个长度为14bp片断的缺失或插入。Pouseeu 〔14〕的研究认为,14bp插入/缺失多态性能影响HLA-G mRNA 的稳定性,提示HLA-G14bp多态性参与了HLA-G转录后的调控; 14bp 缺失多态性还与sHLA-G 分子的表达水平有关〔15, 16〕。本研究结果显示,哈萨克族人携带- 14bp /- 14bp 基因型比携带含+ 14bp的基因型增加食管癌的患病风险( χ2 = 71 .54,P = 0.02,OR = 1. 51,95%C I= 1. 11~ 2.06) ,这可能与血清sH LA-G水平升高有一定关系,从而增加了食管癌的患病风险。
本研究结果还显示,在哈萨克族群体里存在HLA-G* 0105N基因的基因型C /C、C / - ,而未发现- /- 纯合子基因型,HLA-G* 0105N多态性与哈萨克族食管癌的发生无相关性 ( χ2 = 1.13,P = 0.29 )。理论上认为,HLA-G* 0105N 不编码 mHLA-G1和sHLA-G5,但是编码mHLA-G2、G3,sH LA-G6、 G7,而这些可以阻止NK细胞的溶解,而且可溶性HLA-G1 可诱发活性T 细胞的凋亡〔17〕,尽管HLA-G1和H LA-G5蛋白的生物学功能不存在,但是其他亚型的蛋白可以补偿他们的功能〔18〕,因此,可以认为0105N 可能是保护性因素,而C /C 基因型有可能是危险因素,可能增加食管癌的发病风险。本研究观察2个位点联合作用的结果表明,同时携带HLA-G14bp ( - 14bp / - 14bp) 和HLA-G* 0105N ( C /C )的个体比同时携带 H LA-G14bp( + 14bp / + 14bp)和H LA-G* 0105N( C /C )的个体患食管癌风险增加( OR = 2.82,95% CI= 1.32 ~ 6.04),提示今后应增大样本量,对HLA-G* 0105N 位点多态性与食管癌易感性的相关性做进一步研究。
| 〔1〕 | Kroner A, Grmim A, Johannssen K, et al.The genetic in fluence of the nonclassical MHC molecule HLA-G on multiple sclerosis[J].Human Immunology, 2007, 68(5): 422-425. |
| 〔2〕 | Iversen AC, Nguyen OT, Tommerdal LF, et al.The HLA-G 14bp gene polymorphism and decidual HLA-G 14 bp gene expression in pre-eclamptic and normal pregnancies[J].Journal of Reproductive Immunology, 2008, 78(2): 158-165. |
| 〔3〕 | Sipak-Szmigiel O, Cybulski C, Wokolorczyk D, et al.HLA-G polymorphism and in vitro fertilization failure in a Polish population[J].Tissue Antigens, 2009, 73(4): 348-352. |
| 〔4〕 | Veit TD, Cordero EA, Mucenic T, et al.Association of the HLA-G 14bp polymorphism with systemic lupus erythematosus[J].Lupus,2009, 18(5): 424-430. |
| 〔5〕 | Torres M I, Luque J, Lorite P, et al.14-base pair polymorphism of human leukocyte antigen-Gas genetic determinant in heart trans-plantation and cyclosporine therapy monitoring[J].Human Immunology, 2009, 70(10): 830-835. |
| 〔6〕 | Kroner A, Grmim A, Johannssen K, et al.The genetic influence of the nonclassical MHC molecule HLA-G on multiple sclerosis[J].Human Immunology, 2007, 68(5): 422-425. |
| 〔7〕 | Moreau P, Contu L, Alba F, et al.HLA-G gene polymorphism in human placentas: possible association of G* 0106 allele with preeclampsia and miscarriage[J].Biol Reprod, 2008, 79(3): 459-467. |
| 〔8〕 | Castelli EC, Mendes-Junior CT, Vianade Camargo JL, et al.HLA-G polymorphism and transitional cell carcinoma of the bladder in a Brazilian population[J].Tissue Antigens, 2008, 72(2): 149-157. |
| 〔9〕 | 廖佩花, 马彦清, 曾同霞, 等.哈萨克族食管癌与HLA-DR9等位基因关系[J].中国公共卫生, 2009, 25(7): 798-800. |
| 〔10〕 | Miller SA, Dykes DD, Polesky H F.A simple salting out procedure for extracting DNA from human nucleated cells[J].Nucleic Acids Res, 1988, 16(3): 1215. |
| 〔11〕 | Lin A, Li M, Xu D P, et al.Ethnic variation of the HLA-G* 0105N allele in two Chinese populations[J].Tissue Antigens, 2009, 73(3): 270-274. |
| 〔12〕 | 王飞, 李辉, 温泽清.HLA-G 与肿瘤细胞的免疫逃逸[J].中华肿瘤防治杂志, 2007, 14(22): 1751-1754. |
| 〔13〕 | 赵勇, 郑建勇, 王为忠.食管鳞癌患者血清可溶性HLA-G分子表达水平及其临床意义[J].科学技术与工程, 2009, 9(8): 2056-2061. |
| 〔14〕 | Rousseau P, Discorde M L, Mouillot G, et al.The 14bp deletion-insertion polymorphism in the 3' untrans lated region of the HLA-G gene influences HLA-G mRNA stability[J].Hum Immunol, 2003, |
| 〔15〕 | Tripathi P, Abbas A, Naik S, et al.Role of 14 bp deletion in the HLA-Ggene in the maintenance of pregnancy[J].Tissue Antigens,2004, 64(6): 706-710. |
| 〔16〕 | O'Brien M, McCarthy T, Jenkins D, et al.Altered HLA-Gtranscription in pre-eclampsia is associated with allele specific inheritance: possible role of the HLA-G gene in susceptibility to the disease[J].Cell Mol Life Sci,2001, 58(12-13): 1943-1949. |
| 〔17〕 | 王福生, 单际平, 蒋丹斌, 等.食管癌发病特点的变化[J].胃肠病学和肝病学杂志, 2002, 3(11): 256-257. |
| 〔18〕 | Le Discorde M, Le Danff C, Moreau P, et al.HLA-G* 0105N null allelev encodes functional HLA-Gisoforms[J].Biology of Reproduction, 2005, 73(2): 280-288. |