 2010, Vol. 26
2010, Vol. 26


日本血吸虫感染是一个慢性过程,虫体在体内持续存在,不断释放抗原刺激物,宿主体液免疫应答呈现潜伏期短,上升快,平台期高,下降期持久的特点,其中抗体应答的主要抗体为IgG,且抗体亲和力高〔1〕。既往研究多从肉眼进行动态趋势观察,其结论可能存在误差。本实验用BALB/c和C57BL/6两种近交品系小鼠,给予相应处理,定期采集血样,测定血清中虫卵特异性抗体IgG抗体水平,用SAS统计软件中的PROCMIXED过程步从数理统计学角度进行数据分析和统计推断。
1 材料与方法 1.1 材料及主要试剂日本血吸虫感染阳性钉螺(江苏省日本血吸虫病防治研究所),BALB/C和C57BL/6小鼠(扬州大学动物中心),雌性,6~8周龄,平均体重约20g;辣根过氧化物酶(HRP)标记羊抗鼠IgG多克隆抗体(英国AbDSerotec公司);虫卵可溶性抗原(SEA)(江西省血吸虫病防治研究所检测中心)。
1.2 小鼠感染及血清样品采集按常规法自阳性钉螺逸出尾蚴,小鼠在清醒状态下用钻肤法感染尾蚴,(10±2)条/鼠。每种品系小鼠均分为单纯感染、吡喹酮治疗与重复感染组,每组5只。吡喹酮治疗组在感染后第50d用灌胃法一次性给予吡喹酮400mg/kg,重复感染组在首次感染后70d,再次感染(10±2)条/鼠。自感染第1周开始,每隔10d用毛细有机纤维软管自小鼠内眦静脉采血,4000r/min离心15min,分离血清,冻存于-70℃备用,共采血12次。
1.3 酶联免疫吸附法 (ELISA) 测定抗体首先确定最佳抗原包被浓度,4℃包被过夜。再用脱脂奶粉封闭,37℃1h。加一抗(稀释后的小鼠血清,每孔100μL),于37℃反应2h。再加HRP标记的二抗,37℃反应70min。最后加四甲基联苯胺(TMB)行显色反应,37℃0.5h后取出,用H2SO4终止反应,酶标仪450nm波长读取吸光度(A)值。
1.4 数据读入采用SAS软件中的PROCMIXED过程步对实验数据进行分析〔2〕。按照SAS的数据步要求,读入5个变量,分别为小鼠(mice)、品系(breed)、处理(treat)、时间点(t1~t12)以及y(IgG的A值)。其中,小鼠编号为1~30,依次代表不同个体;品系取值为1~2,分别代表BALB/C和C57BL/62种品系小鼠;处理取值为1~3,1为感染组,2为吡喹酮治疗组,3为重复感染组;时间点(t1~t12)取值1~12,依次代表不同采血时间。创建备用数据集。
1.5 PROCMIXED的执行在过程步的CLASS语句中,将小鼠、品系、处理、时间点均看成分组变量;MODEL语句中,固定效应项设为处理和时间点;RANDOM语句中,随机效应项设为处理;Repeated语句中,按照单因素重复测量模型,设为小鼠;由于本研究中的数据属于纵向数据,其数值具有自相关特性,不可以随意互换位置,所以将协方差结构type选项设为AR(autocorrelation)结构。并根据赤池信息准则(AIC)和贝叶斯信息准则(BIC)的结果,评价数据执行结果的拟合度。
2 结 果 2.1 PROCMIXED 固定效应估计结果 (表 1)| 表 1 日本血吸虫IgG抗体在时间上的数值分析结果(从后往前逐次扣除1个时间点) |
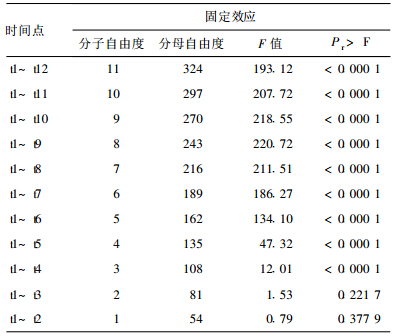
将12个时间点的所有数据读入后进行统计分析,其中IgG抗体在时间(age)之间呈现明显变化(P<0.01)。为更细致定位产生差异时间段,将所有数据从最后1个时间点逐次舍弃,调整程序相应语句后,提交系统运行,从统计分析结果可以看出,前3个时间点(t1,t2,t3,即自感染日起至感染后20d之间)IgG抗体水平无明显变化(P>0.05)。将所有数据从第1个时间点逐次舍弃,调整程序相应的语句后,提交系统运行,结果表明,后6个时间点(t7,t8,t9,t10,t11,t12,即感染后70~120d)IgG抗体水平亦无明显变化(P>0.05)。
2.2 PROCM IXED的模型拟合度结果 (表 2)| 表 2 日本血吸虫IgG抗体拟合度分析结果 |
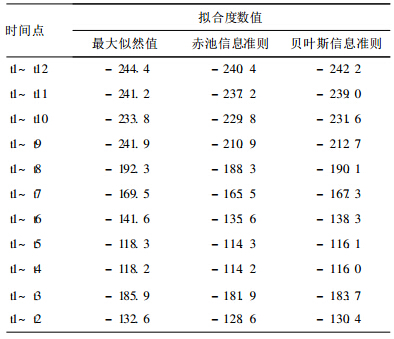
PROCMIXED的运行结果表明,在时间点t4~t12(从前往后递减)以及t1~t6(从后往前递减)之间,随着逐次舍弃,AIC,BIC绝对值越来越小。根据AIC,BIC绝对值越小拟合度越好的原理,表明可溶性虫卵特异性IgG抗体在第t4~t63个时间点(感染后30~60d)随时间呈现明显变化,而在其余时间点变化不明显。
3 讨 论本实验运用SAS软件中的PROCMIXED过程步对日本血吸虫感染小鼠血清中的IgG抗体水平进行了分析。仅2个指标的均数进行简单的相关性分析,则往往只考虑了固定效应,而忽略了随机效应。在本次数据的统计分析中,由于考虑到可溶性虫卵抗原特异性IgG抗体随时间的变化趋势,故对各时间点采用自前向后或自后向前逐步舍弃方法,对数据进行灵活分析,可根据实验数据的结构,选择合适算法,并进行可靠而快速的验证〔2〕。
分析结果表明,将可溶性虫卵抗原特异性抗体IgG水平的动态变化趋势大致分为3个阶段,即感染早期(感染后0~20d),感染早中期(感染后20~30d至60~70d),感染中晚期(感染后70~120d)。其中感染早期IgG抗体水平随时间变化不明显,变化率接近。感染早中期IgG抗体水平则随时间快速上升,而在感染中晚期IgG抗体水平随时间没有明显变化,维持较高水平的平台期。这种特征与生物学上观察到的现象一致。有研究报导〔3, 4, 5〕,日本血吸虫感染宿主后的3周内,由于日本血吸虫尾蚴尚未发育成熟,未排出虫卵,故体内尚未产生可溶性虫卵特异性抗体。随后,日本血吸虫成虫发育成熟,陆续产卵,在可溶性虫卵抗原的刺激下持续产生相应的特异性IgG抗体,故其呈现快速上升趋势。而在感染中后期,由于可溶性虫卵抗原的持续刺激,而IgG抗体水平衰减缓慢,故维持较高水平的平台期。
本实验通过对2种不同品系小鼠进行尾蚴感染,在不同时间点采集血样,并检测相应IgG抗体水平,属于典型的混合线性模型数据,具备重复测量特征〔6, 7, 8, 9〕。本次研究也表明,SAS提供的混合线性模型及相应的PROCMIXED过程可以应用于医学数据的相关性分析中。
| 〔1〕 | 吴观陵.我国血吸虫病免疫诊断发展的回顾与展望[J].中国寄生虫与寄生虫病杂志,2005,23(5):323-328. |
| 〔2〕 | 薛富波,张文彤,田晓燕.SAS 8.2统计应用教程[M].北京:兵器工业出版社,北京希望电子出版社,2004:123-46. |
| 〔3〕 | 杨坤,王显红,周晓农,等.空间流行病学技术在血吸虫病防治研究中应用[J].中国公共卫生,2007,23(8):1017-1019. |
| 〔4〕 | 何家昶,强广翔,伍青,等.山丘型地区人群血吸虫季节性感染规律调查[J].中国公共卫生,2006,22(10):1233-1234. |
| 〔5〕 | 俞小淙,吴观陵,章子豪,等.日本血吸虫感染大鼠血清特异性IgG及IgG2a IgG2c同型水平的动态观察[J].中国血吸虫病防治杂志,2000,12(3):148-150. |
| 〔6〕 | 张岩波,何大卫,刘桂芬,等.重复测量数据的混合模型及其Mixed过程实现——混合线性模型及其SAS软件实现(二)[J].中国卫生统计,2001,18(5):272-275. |
| 〔7〕 | 陈峰,姚晨,孙高,等.新药临床试验中重复测量资料的混合效应模型[J].中国卫生统计,2000,17(6):373-376. |
| 〔8〕 | 萨建刘桂芬.非独立数据的相关性分析[J].数理医药学杂志,2007,20(6):821-824. |
| 〔9〕 | 赵晋芳,刘桂芬.重复测量线性混合模型中方差-协方差结构的选择[J].现代预防医学,2004,31(1):11-12. |