 2010, Vol. 26
2010, Vol. 26


2. 广州军区广州总医院;
3. 中山大学第三附属医院;
4. 南方医科大学公共卫生与热带医学学院
脑卒中是中国第二大死亡原因和首位致残原因,是以缺血缺氧性脑神经元死亡为病理特征的一类疾病,但目前临床治疗上缺乏有效的治疗手段。三磷酸腺苷(ATP)敏感性钾通道(KATP通道)参与多种细胞功能调控,同时也涉及神经元再生及死亡等过程〔1, 2, 3〕。有研究表明,在缺氧的情况下,KATP通道的激活对缺氧中的神经元具有明显保护作用〔4〕,但其机制不明。缺氧诱导因子-1(hypoxia-inducib lefactor,HIF-1)是机体缺氧刺激诱导产生的特异性转录因子,其激活相关基因的表达,以适应缺氧的环境〔5〕。本研究通过离体培养海马神经元缺氧模型,探讨KATP通道的开放对缺氧神经元的保护作用是否通过调节HIF-1 表达变化实现,为脑卒中等疾病的临床治疗提供科学依据。
1 材料与方法 1.1 试剂与仪器DMEM高糖培养液干粉(美国GIBCO公司);甲糖宁、二氮嗪、Hoechst 33342染色液、10 mg/L多聚赖氨酸(美国Sigma公司);血清(杭州四季青公司);Trizol试剂 (美国Invitrogen公司);一步法RNA PCR试剂盒(日本Takara公司);HIF-1 多克隆抗体(美国SantaCruz公司);β-actin多克隆抗体(武汉博士德公司)。MODEL818 CO2细胞培养箱(美国NAPCO公司);酶标仪(台湾ERMA公司);BH2-RFL-T3荧光显微镜、IX700倒置显微镜(日本Olympus公司);低温高速离心机、DU-530核酸蛋白分析仪(美国Beckman公司);DocG el2000凝胶扫描成像系统、电泳仪(美国Bio-Rad公司)。
1.2 实验动物出生24 h以内的SD新生大鼠100只(南方医科大学实验动物中心,许可证号:SCXK粤200620015),雌雄各半。
1.3 细胞培养与分组取出生24 h以内的SD大鼠,无菌条件下分离出海马,0 125%胰酶消化、分散并制成1×106/mL密度的细胞悬液,接种于包被有多聚赖氨酸的35 mm塑料培养皿或96孔板中(2 mL/皿或100μL/孔),置于37%、含5% CO2的培养箱培养。接种后第3 d,加入细胞分裂抑制剂阿糖胞苷4μg/mL以抑制非神经细胞的过度增殖,作用48 h后更换新鲜培养液。之后每3 d换液1次,每次更换50%新鲜培养液。培养1周后,将细胞分为4组:第1组为正常氧分压组,在正常氧状态中(5% CO2、95%空气)孵育;另外3组分别为单纯缺氧组、缺氧+二氮嗪组、缺氧+甲糖宁组,均在(5% CO2、95% N2)模拟的慢性重度缺氧状态孵育〔6, 7〕,缺氧时间持续8,12,24 h。缺氧+二氮嗪组、缺氧+甲糖宁组在缺氧同时分别给予KATP通道的激动剂二氮嗪(100μmol/L)和阻断剂甲糖宁(100μmol/L)。
1.4 神经元凋亡检测采用免疫印迹方法检测试验持续12 h后各组HIF-1蛋白表达水平。将收集的神经元裂解,进行聚丙烯酰胺凝胶电泳,电泳后将蛋白转移至聚偏二氟乙烯膜 (PVDF膜)。转移结束后,取出PVDF膜,依次置于5%脱脂奶粉、磷酸盐缓冲液、一抗、二抗中孵育,使用化学发光法暗室显影,使用Alpha凝胶成像系统采集信号,使用Fluo rChem软件进行光密度分析。
1.6 统计分析采用SPSS 10.0软件进行单因素方差分析和最小显著差法(LSD法)检验。
2 结 果 2.1 缺氧诱导海马神经元凋亡将培养7 d的海马神经元暴露于缺氧环境中,缺氧后不同时间显微镜下观察,发现随缺氧时间延长,缺氧组出现胞体肿胀或皱缩、突起回缩或消失、胞核凝聚的神经元比例增加。MTT结果表明,正常氧分压组细胞存活率为(100±0.98)%,缺氧8,12,24 h后,细胞的存活率分别降至(85.76±3.31)%,(80.13±1.76)%,(72.24 ±3.87)%,差异均有统计学意义(P < 0.05)。
2.2 各组不同时间点神经元凋亡率(表 1)| 表 1 不同时间点各组神经元凋亡率(%) |
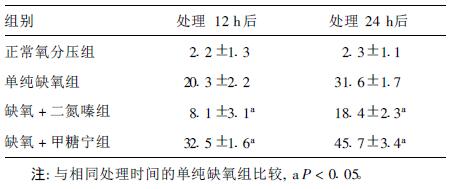
处理缺氧+二氮嗪组和缺氧+甲糖宁组12,24 h后的神经凋亡率与单纯缺氧组比较,差异均有统计学意义(P < 0.05)。
2.3 处理12h后各组HIF-1 蛋白表达情况单纯缺氧组 HIF-1 蛋白表达较正常氧分压组明显升高(P < 0.05)。与单纯缺氧组比较,缺氧+甲糖宁组HIF-1 的蛋白表达水平明显降低(P < 0.05),而缺氧+二氮嗪组HIF-1 的蛋白表达水平明显升高(P < 0.05)。
 |
注: 1: 正常氧分压组; 2: 单纯缺氧组; 3: 缺氧+ 二氮嗪组; 4: 缺氧+ 甲糖宁组。 图 1 处理12 h后各组HIF-1α 蛋白表达情况 |
有研究表明,在缺氧情况下,KATP通道的激活对缺氧中的神经元具有明显保护作用〔4〕,其作用机制尚不清楚。本研究结果表明:在缺氧条件下,KATP通道的开放剂二氮嗪能够增加HIF-1 的蛋白表达并减少神经元凋亡,而KATP通道阻断剂甲糖宁则降低HIF-1 的蛋白表达并促进神经元凋亡,与文献〔9, 10〕结果一致,而且提出了KATP通道保护细胞新的可能机制,即促进HIF-1 的蛋白表达。HIF-1可结合缺氧反应元件(HRE)中的特定序列,促使其下游具有神经保护作用的靶基因如血管内皮生长因子(VEGF)等转录增加〔11, 12〕,从而使组织和细胞适应低氧环境〔13〕。
KATP通道对HIF-1的调节作用依赖于其对钾离子的通透作用,但具体的下游信号转导通路还有待进一步研究。一种可能的机制是,当KATP通道激活时,外流的钾离子影响细胞内钙离子的浓度,从而激活钙依赖性的信号转导机制,进而影响HIF-1的表达。同时,KATP通道的激动剂及阻断剂也可能改变通道的构象,并进而影响与KATP通道相互结合的蛋白质而改变下游的信号转导通路,从而影响HIF-1的表达。
| 〔1〕 | Alkan T.Neuroproctective effects of ischemic tolerance(preconditioning)and postconditioning[J].Turk Neurosurg,2009,19(4):406-412. |
| 〔2〕 | Assad AR,Delou JM,Fonseca LM,et al. The role of KATP channels on propofol preconditioning in a cellular model of renal ischemia reperfusion[J].Anesth Analg,2009,109(5):1486-1492. |
| 〔3〕 | Gier B,Krippeit Drews P,Sheiko T,et al. Suppression of KATP channel activity protectsmurine pancreatic beta cells against oxidative stress[J].J Clin Invest,2009,119(11):3246-3256. |
| 〔4〕 | BallanyiK.Protective role of neuronal KATP channels in brain hypoxia[J].J.Exp.Biol, 2004,207:3201-3212. |
| 〔5〕 | Webb JD,Coleman ML,Pugh CW.Hypoxia,hypoxia inducible factors(HIF),HIF hydroxylases and oxygensensing[J].Cell Mol Life Sci, 2009,66(22):3539-3554. |
| 〔6〕 | Craw ford RM,Jovanovi S,Budas GR,et al. Chronic mild hypoxia protects heart derived H9c2 cells againstacute hypoxia/reoxygenation by regulating expression of the SUR 2A subunit of the ATP sens itive K+ channe.l[J].J Biochem,2003,278(33):31444-31455. |
| 〔7〕 | DelToro R,Lev itsky KL,L pez Barneo J,et al. Induction of T type calcium channel gene expression by chronic hypoxia.[J].J Biochem,2003,278(25):22316-22324. |
| 〔8〕 | Smiard JM,Woo SK,Bhatta S,et al. Drugs acting on SUR1 to treat CNS ischemia and trauma[J].Curr Opin Pharmacol, 2008,8(1):42-49. |
| 〔9〕 | 赵吉清,林海,赛燕,等.缺氧对PC12细胞IL-6及GP130基因表达影响[J].中国公共卫生,2009,25(3):289-291. |
| 〔10〕 | NicholsCG.KATP channels as molecular sensors of cellularmetabolism[J].Nature,2006,440(7083):470-476. |
| 〔11〕 | Mu D,Jiang X,Sheldon RA,et al. Regulation of hypoxia inducible factorla lpha and induction of vascular endothelial growth factor in a ratneonatal strokemodel[J].Neurobiol Dis,2003,14(3):524-534. |
| 〔12〕 | Chang YS,Mu D,WendlandM,et al. Erythropo ietin miproves functional and histological outcome in neonatal stroke[J].Pediatr Res,2005,58(1):106-111. |
| 〔13〕 | SemenzaGL.Regulation of oxygen homeostasisby hypoxia inducible factor 1[J].Physiology(Bethesda),2009,24:97-106. |